Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Pedro Duque | -- | 4561 | 2022-11-03 11:49:36 | | | |
| 2 | Amina Yu | + 1 word(s) | 4562 | 2022-11-04 02:32:15 | | | | |
| 3 | Amina Yu | + 2 word(s) | 4564 | 2022-11-04 02:38:53 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Duque, P.; Vieira, C.P.; Vieira, J. Novel Animal Vitamin C Biosynthesis Pathways. Encyclopedia. Available online: https://encyclopedia.pub/entry/32750 (accessed on 08 August 2026).
Duque P, Vieira CP, Vieira J. Novel Animal Vitamin C Biosynthesis Pathways. Encyclopedia. Available at: https://encyclopedia.pub/entry/32750. Accessed August 08, 2026.
Duque, Pedro, Cristina P. Vieira, Jorge Vieira. "Novel Animal Vitamin C Biosynthesis Pathways" Encyclopedia, https://encyclopedia.pub/entry/32750 (accessed August 08, 2026).
Duque, P., Vieira, C.P., & Vieira, J. (2022, November 03). Novel Animal Vitamin C Biosynthesis Pathways. In Encyclopedia. https://encyclopedia.pub/entry/32750
Duque, Pedro, et al. "Novel Animal Vitamin C Biosynthesis Pathways." Encyclopedia. Web. 03 November, 2022.
Copy Citation
Vitamin C (VC) is a water-soluble compound that, at physiological pH, is mainly found in a reduced ionizable form (L-ascorbate). The potential applications of VC have been the subject of research since it was first synthesized in 1933 to the present day, in many areas relevant to mankind, such as human nutrition, animal feed supplementation, agricultural and industrial waste management, and sustainable nanomaterial synthesis. This nutrient gained further relevance as a potential therapeutic agent for COVID-19 prevention and treatment.
ascorbic acid
evolution
synthesis
aldonolactone oxidoreductases
insects
nematodes
1. Vitamin C Biosynthetic Pathways: The Current Status
Although the origins of the divergent VC synthesis mechanisms in eukaryotes have not been completely elucidated, three main metabolic pathways have been well-described in photosynthetic protists, algae, plants and animals [1][2][3][4][5][6][7][8]. In addition, fungi species are known to synthesize VC analogs, such as D-erythroascorbate [9]. Due to the comprehensive reviews on the subject [1][10][11][12][13], and lack of significant recent advances, these will be only briefly discussed to highlight the main differences between pathways.
One of the distinctive features of the main VC synthesis pathways is the sugar precursor. Therefore, the main VC synthesis pathways are shown, not according to taxonomic relationships, but in order to highlight this point, as well as the sharing of the last enzyme of the pathway between photosynthetic protists and plants and green algae, and the sharing of the first enzyme of the animal and photosynthetic protists pathways (Figure 1). While plants rely mainly on D-Mannose as the main substrate for VC synthesis, animals and photosynthetic protists utilize D-Glucose [10]. Starting from different sugars, and by the use of divergent enzymatic mechanisms, plants and animals convert these sugars to L-Galactono-1,4-lactone and L-Gulono-1,4-lactone in the penultimate step of the pathway, respectively [10]. In photosynthetic protists, however, VC synthesis relies on a mix of animal and plant enzyme orthologs to confer the hybrid capacity for the conversion of D-Glucose to L-Galactono-1,4-lactone [10]. L-Galactono-1,4-lactone is ultimately metabolized to VC by the action of a L-Galactono-1,4-lactone dehydrogenase (GALDH) in plants and photosynthetic protists, while in animals, L-Gulono-1,4-lactone originates VC through an oxidation step catalyzed by L-Gulonolactone oxidase (GULO) [10]. It should be noted that some of the intermediate sugar substrate conversion steps in the animal and photosynthetic protists pathways are still hypothetical, since the putative enzymes involved in these steps have not yet been isolated and functionally characterized. For instance, the UDP-Glucuronidase that catalyzes the transition of UDP-D-Glucuronate to D-Glucuronate is thought to exist in vertebrate species but has not yet been identified [14]. Fungi species rely on the five carbon D-Arabinose sugar to synthesize D-erythroascorbate after two conversion steps catalyzed by D-Arabinose dehydrogenase and D-Arabinono-1,4-lactone oxidase (ALO), respectively [10].
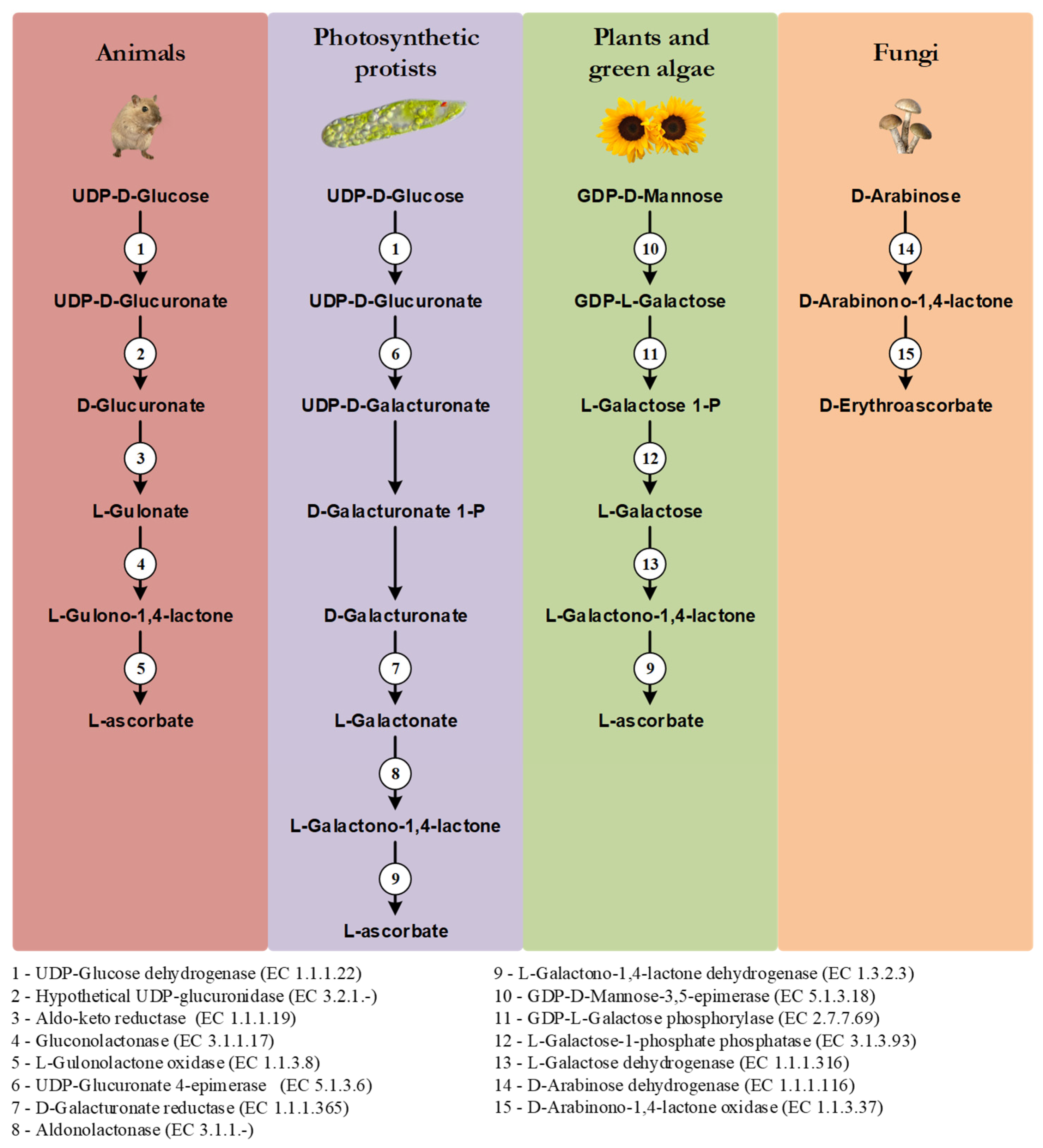
Figure 1. Graphical illustration of the main VC and D-erythroascorbate synthesis pathways, adapted from the MetaCyc database [15]. The numbered circles represent enzymes that are known to catalyze the substrate conversions within the pathways (see the enzyme list presented below the illustration). Reaction steps without an attributed enzyme are performed by unknown enzymes.
In plants, alternative mechanisms in which myo-Inositol, L-Gulose and D-Galacturonate are used as precursors of VC synthesis, have been proposed [16][17][18]. They are based on species-specific observations, and they have not been yet verified in a wider taxonomic context, and thus are not here discussed.
2. Alternative Animal VC Synthesis Pathways
The pseudogenization or even complete loss of the GULO gene is often reported as the primary cause of VC synthesis inability, in species such as humans, teleost fish, insects and nematodes [19][20][21][22][23]. Nevertheless, recent findings challenged the idea that the absence of a functional GULO gene can be used as a marker of VC synthesis inability in all animals [24][25][26]. In addition, although scarcely mentioned, the notion that D-Glucose is exclusively used for animal VC synthesis has also been challenged [27].
2.1. VC Synthesis in Caenorhabditis Elegans
C. elegans, where both GULO and Regucalcin (Regucalcin precedes GULO in the animal VC synthesis pathway) genes are missing, synthesizes VC [25]. Indeed, after exposing individuals to 13C-labeled Escherichia coli as food source, 13C was found in the VC pool, which led to the conclusion that VC must be synthesized de novo [25]. It was also observed that total VC content did not correlate with the concentration of sugar precursors commonly used by animals or plants, as well as their relevant derivatives [25]. Nevertheless, it is not clear that an increase in such sugar precursors would necessarily lead to an increase in VC synthesis [25]. In addition, a comprehensive search for homologs of the enzymatic constituents of the plant and animal VC synthesis pathways in C. elegans was also performed, revealing that no pathway was present as a complete set [25]. The loss of GULO and Regucalcin, is also observed in other nematode species [28]. Given the evidence, it was suggested that VC synthesis in C. elegans likely occurs through a yet undescribed metabolic pathway [25]. No enzymatic constituents have been proposed for this pathway [25].
2.2. VC Synthesis in Drosophila Melanogaster
In insects, the need for VC synthesis, or adequate overall VC levels, varies depending on the species group that is considered [29]. In species from the Diptera, Lepidoptera and Coleoptera taxonomic groups, VC plays an important metabolic role [29]. Indeed, VC synthesis has been observed in the dipteran D. melanogaster [30] and in the lepidopteran Bombyx mori [26]. Massie et al. [30] were the first to note that D. melanogaster Oregon-R and Swedish-C male flies maintained for over 10 generations in food and water sources deprived of VC, displayed measurable total body VC concentrations. Moreover, by observing the continuous decrease in VC content in flies with age, it was suggested that VC synthesis happened during the developmental stages, rather than in adult individuals [30]. Nevertheless, the amount of total VC increases in adult flies after exposure to a brief cold shock [30], indicating that in D. melanogaster VC synthesis might be also involved in stress response mechanisms. It was evaluated that the impacts of supplementation with L-Gulonolactone, D-Galactonolactone, and Glucose in adult flies, but none of these substrates contributed to increased VC content after the cold shock stimulus [30]. As it was discussed, the enzymatic mechanisms might simply not be stimulated by dietary precursors [30]. This would suggest that D. melanogaster is able to synthesize VC precursors above the limiting rate of consumption, and that this self-sufficiency overshadows their addition. Herein, however, did not address the role of the flies’ gut microbiome in the synthesis process. This assessment is essential to determine the source of the VC found in the whole body homogenates, as several microbes have been found to synthesize and supply vitamins to hosts incapable of endogenous synthesis [31], and VC synthesis by insect symbionts has been previously observed [32].
Based on these findings, Henriques et al. [24] have demonstrated that axenic D. melanogaster Oregon-R flies possess similar levels of VC relative to normal individuals, and that the ex vivo culture of the microbiome did not lead to any VC accumulation in the resulting bacterial pellet and supernatant. Thus, Oregon-R flies’ microbiome does not significantly contribute to VC synthesis [24]. The impact of flies’ gender and sexual activity, not considered in [30], was also evaluated [24]. VC content in virgin flies had no significant difference relative to mated flies, but females had approximately three times more VC than males, and this observation could not be exclusively explained by the different bodyweight between genders [24]. This result implies that female flies can demonstrate increased amounts of VC due to: (i) the overexpression of the VC synthesis pathway; (ii) the presence of gender-specific structures, such as the ovaries; and (iii) increased VC storage and retention. Plasma VC concentrations are also higher in human and mouse females [33][34], but by a slight margin when in comparison to what is observed in D. melanogaster. The VC difference observed in mice may be explained in part by distinct renal VC excretion rates between males and females, since females are able to retain more VC from urine than males [35]. Therefore, gender-specific VC excretion rates along the D. melanogaster Malpighian tubule system, as well as the 50% difference in bodyweight between the sexes [24], might account for the observed results. These hypotheses, however, remain to be experimentally addressed.
It has been suggested that the physiological responses of insects can vary considerably depending on the cold exposure conditions [36]. Thus, as a complement to the brief cold shock exposure experiment reported in [30], Henriques et al. [24] verified the effects of extended cold exposure on the VC content of D. melanogaster Oregon-R. Unlike what was observed in [30], flies acclimated one day at 15 °C had a 23% decrease in VC levels relative to flies kept at 25 °C [24]. Nevertheless, cold-acclimated flies were able to replenish base VC levels after one day of recovery at 25 °C [24]. Therefore, different cold exposure conditions impose contrasting changes in D. melanogaster’s VC metabolism. Interestingly, the gene that encodes the lactonase that precedes GULO in the animal VC synthesis pathway, Regucalcin, has a duplicate in Sophophora species such as D. melanogaster, called Drosophila cold acclimation (Dca) [37][38]. The functions of this gene are still being determined, but so far they appear to be correlated with the response to cold exposure and wing size variation [37][38][39][40]. In regard to cold exposure, Dca has contrasting expression changes depending on the applied stress, since it is upregulated under acclimation conditions [36][38], but downregulated under cold shock [41][42]. This information implies that Dca expression changes are likely not correlated with the VC synthesis changes that occur under cold exposure responses in Sophophora species. Nevertheless, without further experimental data, the involvement of Dca in VC synthesis under cold exposure conditions cannot be yet ruled out. A direct involvement of Dca or Regucalcin in VC synthesis would imply that at least the lactonase component of the animal pathway is conserved in D. melanogaster. Using an in silico methodology, both Regucalcin and Dca were inferred to be involved in calcium homeostasis and in the oxidative stress response, but so far, no evidence was found for its involvement in VC synthesis [28]. The involvement in the calcium homeostasis and in the oxidative stress response is expected given the multiple functions of Regucalcin in vertebrates (reviewed in [43]). The inference of a divergent protein interaction surface at Dca in comparison with Regucalcin, along with the detection of positively selected amino acids near and within the active site region, suggests that Dca might have also acquired novel functions involving its lactonase activity and overall contribution to protein complexes [28]. The constitutive suppression, under the actin 5C driver, of either Regucalcin or Dca in D. melanogaster, did not clarify the role of these genes in VC synthesis [28]. Indeed, both genes are required for D. melanogaster viability, since in both cases over 70% of the individuals with suppressed expression were only able to reach the pupal stage [28], and thus the biological processes in which they are involved could not be determined. The reduced viability could be attributed, for instance, to the dysregulation of calcium homeostasis/signaling, as stated in [28]. Nevertheless, a cascade of metabolic events caused by VC deprivation could also explain the negative effects, especially when considering the previous evidence that VC synthesis occurs mainly during the developmental stages [30], which coincides with the lethal phase observed in the fly RNAi experiments [28]. In B. mori, a Regucalcin-like gene has been inferred to be involved in VC synthesis [26] (see below).
2.3. VC Synthesis in B. mori
In the lepidopteran B. mori, where GULO is absent, an alternative pathway for VC synthesis has been proposed, based upon the identification of a protein with GULO-like activity [26]. Evidence for VC synthesis comes from the observation that during silkworm development, the concentration of this nutrient is stable or even increased during stages without an active consumption of VC from mulberry leaves [26]. As in D. melanogaster [24], it was addressed that the impact of the microbiome, and found that it does not significantly contribute to VC synthesis [26]. Furthermore, the contribution of VC regeneration mechanisms in VC content fluctuations was also dismissed, since DHA levels remained unaltered during egg development [26]. Thus, it was hypothesised that B. mori is capable of endogenous VC synthesis [26]. Using a bioinformatics approach, Hou et al. [26] were able to identify two candidate GULO-like sequences (BGIBMGA012624 and BGIBMGA005735). Because BGIBMGA005735 codes for a protein that has a domain with similarity with another aldonolactone oxidoreductase (AO), it was selected for further analyses [26]. Enzymatic activity assays showed that this GULO-like protein is capable of VC synthesis using the same substrate as GULO, and that its activity is in part modulated by physiological changes that occur in the fat body during pupae development, implicating this tissue in the process [26]. This information is intriguing, as the B. mori fat body is considered to be analogous to the vertebrate liver and adipose tissue [44], and VC synthesis is known to occur in the liver of higher vertebrates [19][45]. Nevertheless, it is believed that in vertebrates, VC synthesis transitioned from the kidney to the liver during evolution and not the opposite [19], as now suggested by the observation that in protostomians the tissue responsible for VC synthesis is the structure that is analogous to the liver. It is, however, conceivable that VC synthesis might have transitioned from the kidney analogous tissue to the fat body in insects, in a manner similar, but independent of what is observed in vertebrates. In B. mori, the siRNA suppression of the GULO-like gene expression in the pupae fat body decreased VC synthesis but was ultimately non-lethal, while in the egg stage it considerably reduced overall survivability and led to delayed hatching [26]. Thus, although not ubiquitously essential for the complete growth of silkworms, this gene appears to have relevant functions implicated in viable transitioning from the egg to the larval stage [26]. The direct correlation between the reduced fitness of silkworm eggs and VC levels is, however, missing in that study. It was noted that, however, that VC synthesis associated with the activity of the GULO-like protein studied, is not sufficient to fulfil the nutritional needs of developing individuals without additional supplementation [26]. The B. mori GULO-like protein is the ortholog of the vertebrate Delta(24)-sterol reductase (DHCR24) [46][47]. Like the vertebrate GULO, sterol reductases belong to the vanillyl alcohol oxidase/para-cresol methylhydroxylase (VAO/PCMH) flavoprotein family, but are usually known to catalyze distinct reactions [48]. While the DHCR24 protein is known to catalyze the NADPH-dependent reduction of a carbon–carbon double bond during the synthesis of cholesterol [48], using as substrate desmosterol that derives from endogenous [49] or dietary [50] sources, GULO is known to catalyze the oxidation of L-Gulonolactone, using O2 as an electron receptor, in the synthesis of VC [48][51]. In agreement with the functional divergence, phylogenetic studies also show that sterol reductases and AOs have diverged early in evolution [48]. Thus, it was not expected that a sterol reductase could metabolically replace GULO.
Considering that B. mori depends exclusively on mulberry leaves for nutrition purposes [52][53], and the VC content in these tends to differ depending on factors such as the climate or plant variety [54], the novel DHCR24 GULO-like functions might be the result of selective pressures to enhance VC availability during the development stages of this specific organism. Assuming that the reduced viability of silkworm eggs was caused by lower VC levels after DHCR24 knockout, this evolutionary perspective could explain why VC synthesis mediated by DHCR24 seems to play an important but not essential role in silkworm growth and fitness. This interpretation correlates well with previous data that imply that some enzymes involved in insect sterol metabolism have likely evolved additional functions throughout evolution [55]. Nevertheless, the available data do not rule out the diminished sterol reductase activity as the partial or complete cause of the increased egg lethality.
Hou et al. [26] proposed that B. mori synthesizes VC through an adapted animal pathway in which the identified GULO-like enzyme functionally replaced GULO in the catalysis of the final oxidation step. This hypothesis appears to be the most parsimonious, since although GULO was lost in B. mori, the genes that encode enzymes that catalyze upstream enzymatic reactions in the animal pathway, such as Regucalcin, are present in this organism [28]. It should be noted, however, that this GULO-like enzyme is not present in D. melanogaster (by performing a “blastp” search (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 28 July 2022) using the B. mori DHCR24 sequence (XP_004926865.1) as query, provided no homologous sequence hits in D. melanogaster) and thus, the B. mori VC pathway evolved independently from that of D. melanogaster. The nomenclature used in Hou et al. [26] can be confusing, since it was referred to this animal modified pathway as the L-Gulose pathway, and state that it is analogous to the one found in plants. The proposed plant L-Gulose pathway has partial similarity with the animal D-Glucose pathway, since they both rely on L-Gulonate as intermediate substrate [56]. They differ, however, in the molecular mechanisms that lead to this step, as in the characterized animal pathway this molecule derives from D-Glucuronate and in plants from L-Gulose [56][57]. The metabolism of L-Gulose seems to be restricted to plants and archaea species [58], since the formation of L-Gulose likely involves a reaction catalyzed by a GDP-Mannose 3′,5′-Epimerase (GME) [18][58][59][60], and such epimerase activity was only found associated with the conversion of GDP-D-Mannose to GDP-L-Galactose in animals [61]. Considering these observations, and that naturally occurring L-Gulose is considerably rare [62], VC synthesis in B. mori likely does not rely on this precursor, and thus the nomenclature referred to in [26] should be interpreted with caution. Nevertheless, since plant GME can convert GDP-D-mannose to a mixture of GDP-L-Galactose, GDP-L-Gulose and GDP-D-Altrose [60], the contribution of enzymes with GDP-Mannose 3′,5′-Epimerase activity to protostomian L-Gulose metabolism, such as the GDP-4-keto-6-deoxy-D-mannose 3,5-epimerase/4-reductase of D. melanogaster [63] and its homologs in other species, cannot at present be excluded as playing a role in VC synthesis in protostomian species that do not use the described animal synthesis pathway.
2.4. The Role of Myo-Inositol in Animal VC Synthesis
Like in plants, the role of myo-Inositol as a precursor for animal VC synthesis through a partially alternative pathway has been addressed. The animal myo-Inositol Oxygenase (MIOX) converts this sugar to D-Glucuronate [64][65], which is a key molecule in the glucuronate-xylulose pathway [66][67] and an intermediate precursor in the vertebrate VC synthesis process ([10]; Figure 1). Experiments in Cavia porcellus (guinea pig) first suggested that myo-Inositol played an important role in VC synthesis or sparing [68], but later studies showed that this was not the case [69]. Indeed, there is no functional GULO gene in C. porcellus, and this is why this species is highly regarded as a scorbutic model organism when missing a dietary source of this nutrient [70][71][72]. The auxotroph nature of the guinea pig should have been considered a limiting methodologic factor in addressing the role of myo-Inositol in animal VC synthesis, as the absence of a functional VC synthesis pathway could invalidate the study of potential precursors in this organism. This criticism stands even considering that this polyalcohol is naturally present in the guinea pig [73][74] in sufficient concentration for viable VC synthesis [27]. The conclusions referred in [69] were thus, expected. In this sense, teleost species are also scurvy prone in the absence of VC supplementation [75][76], and thus should be avoided as experimental subjects in further explorations on the subject.
Myo-Inositol was found not to be involved in VC synthesis in Rattus norvegicus (rat) [77]. Unlike the guinea pig, rats are capable of synthesizing VC using the characterized animal pathway [78]. In placental mammal species capable of doing so, VC synthesis occurs in the liver since it relies on the hepatocyte glycogen reserves [79], and GULO is exclusively found in this tissue [19][45][80]. The D-Glucuronate needed for the process can be obtained as the by-product of the glucuronidation of endobiotic and xenobiotic compounds in the mammals liver [14][81][82], as well as the conversion of UDP-D-Glucuronate to D-Glucuronate by a putative UDP-glucuronidase [14] (see Figure 1). However, myo-Inositol is only metabolized in the vertebrate kidney [83], and, as expected given this observation, MIOX appears to be a kidney-specific enzyme [65][84]. The D-Glucuronate originated by the action of MIOX is likely mostly used in the glucuronate-xylulose pathway that is particularly active in the vertebrate kidney [85]. Therefore, although it is conceivable that myo-Inositol could play a role both in VC synthesis and in the glucuronate-xylulose pathways [86][87], it is likely that only a small amount of D-Glucuronate diffuses from the kidney to the liver, thus playing a negligible contribution in the context of VC metabolism in mammals that synthesise VC in the liver.
The observations made in rats may not apply to all mammal species capable of endogenous VC synthesis. Indeed, in egg-laying mammals (Monotremata), GULO is exclusively found in the kidney [88]. In the Peramelina order (Marsupials) GULO is found in both the liver and kidney, although in species from the Diprotodonta order (Marsupials), GULO is mainly found in the liver [88]. Therefore, in basal mammal groups, besides D-Glucose, myo-Inositol may also be a relevant substrate for VC synthesis in species where GULO is found in the kidney.
Hänninen et al. [27] compared the capacity for VC synthesis from myo-Inositol in the Leghorn hen (Gallus domesticus) kidney and rat liver. In [27], the findings of [77] were replicated, as rat liver extracts displayed no MIOX activity and were unable to synthesize VC from myo-Inositol [27]. However, the analyses performed on the galliform species capable of VC synthesis in the kidney [89] showed opposite results [27]. Herein, it was able to determine that 14C-labelled myo-Inositol was incorporated into the hen kidney VC pool, in a manner partly dependent on the MIOX ability to convert this sugar to D-Glucuronate [27]. In addition, the results also showed that the concentration of this polyalcohol in hens is sufficient for biologically relevant VC synthesis [27]. Not all birds have, however, GULO in the kidney, an essential requisite for VC synthesis in this organ. For instance, most Passeriformes species synthesize VC in the liver [89].
In amphibians [90][91], reptiles [90][91] and basal fish groups [90][92], GULO is only found in the kidney as well. Therefore, although generally disregarded, the contribution of myo-Inositol to VC synthesis in animals, particularly in ancestral vertebrate species, cannot be excluded.
2.5. Summary and Future Perspectives
In this section, hhypotheses regarding VC synthesis pathways was put forwarded in nematodes and insects (Figure 2), and present possible methodological approaches for further exploration of this subject. The available information strongly suggests that VC synthesis in C. elegans occurs by a very distinct pathway from the one used by most animals, since it may not rely on the known substrates of the described pathways as precursors [25], and the two key enzymes in the process, namely Regucalcin and GULO [25][28], are also missing (see detailed discussion above). The precursor supplementation assays have some experimental limitations [25], that can be overcome using radiolabeled compounds, since this methodology allows the direct visualization of substrate incorporation and excretion rates [93]. In addition to the classical substrates, the assays could also be expanded to include myo-Inositol, since the vertebrate MIOX gene has an ortholog in C. elegans (C54E4.5; Ensembl accession number WBGene00016920), and the D-Glucuronate that is generated could contribute to VC synthesis as it happens in birds (Figure 2). The use of a complete or partial knockout of C54E4.5 could also be used to address the involvement of this gene in VC synthesis by looking at VC levels that can be easily measured using the methodology described in [24].
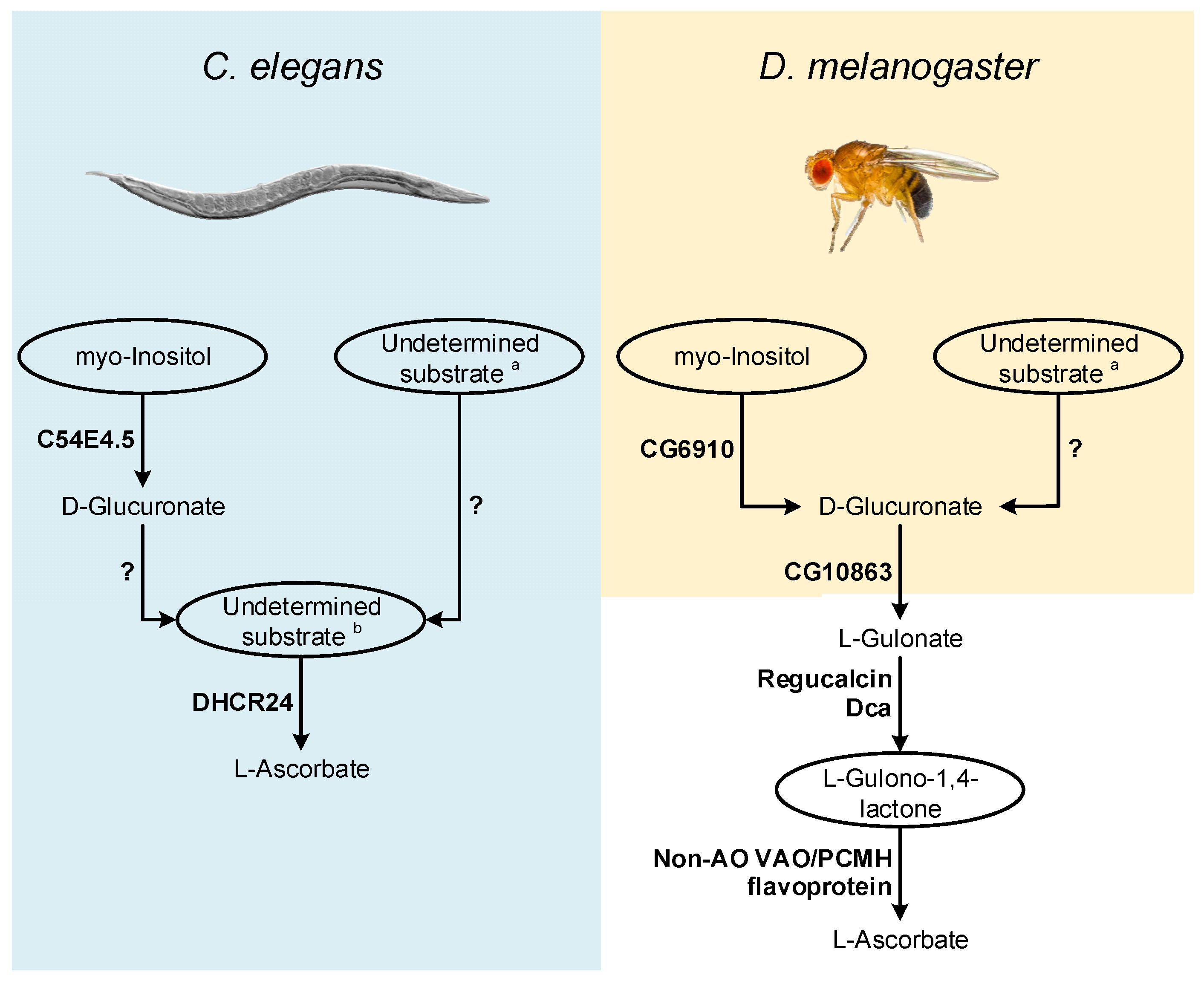
Figure 2. Inferences on enzymatic mechanisms and possible substrates utilized by C. elegans and D. melanogaster in the VC animal synthesis pathways. The ellipses highlight the main substrates possibly used in the proposed pathways, but that remain to be addressed using appropriate experimental setups. Enzymes that may be correlated with VC synthesis in these species are highlighted in bold next to the metabolic steps they are likely to catalyze. The interrogation marks indicate uncertainty in the steps regarding mechanisms of conversion of uncharacterized substrates, and the relative contribution of these processes to the availability of VC precursors. Common sugars (such as D-mannose, L-Galactose, D-Glucose and D-Arabinose) are assigned with a while L-Galactono-1,4-lactone, L-Gulono-1,4-lactone and D-Arabinono-1,4-lactone are assigned with b.
The report of an AO function for the B. mori DHCR24 sterol reductase [26] also raises questions regarding possible adaptations of non-AO VAO/PCMH flavoproteins for VC synthesis (Figure 2). C. elegans contains a functional DHCR24 ortholog [94][95] that should be also studied in detail.
As in C. elegans, in flies, the supplementation of food with precursors naturally used in the animal VC synthesis pathway did not contribute to increased VC content [8] (see detailed discussion above). However, as suggested for C. elegans, the use of radiolabeled substrates is likely more adequate to address this issue. Such approach could be particularly relevant in deciphering the role of myo-Inositol in D. melanogaster VC metabolism, since a MIOX ortholog has been identified in this organism (CG6910; [96]; Figure 2). In D. melanogaster, under cold acclimation conditions, the expression of the CG6910 gene is reduced [36]. Dca is overexpressed under cold acclimation conditions [36], but VC levels decrease during this stimulus [24], which may be due to the suppression of CG6910 expression during the cold stress response.
Both Dca and Regucalcin likely catalyze the conversion of L-Gulonate to L-Gulono-1,4-lactone in D. melanogaster, and as such, if Dca is involved in VC synthesis, through a pathway resembling the one found in vertebrates, then the availability of its substrate might be a limiting factor for VC synthesis during cold acclimation conditions. L-Gulonate is obtained by the transformation of D-Glucoronate by the action of the aldo-keto reductases AKR1A1 and AKR1B1 in mammals [97][98] (Figure 1). The catalytical contribution of the two enzymes is very distinct, as AKR1A1 and AKR1B1 are responsible for approximately 85% and 15% of the conversion ratio, respectively [98]. The reduced contribution of AKR1B1 can be explained by its reduced specific activity rather than substrate specificity [99], reflected in the reduced efficiency for D-Glucuronate reduction when compared to AKR1A1 [98]. The human AKR1A1 gene has no orthologs in D. melanogaster (Ensembl accession number ENSG00000117448), but the AKR1B1 gene displays a one-to-many ortholog relationship (Ensembl accession number ENSG00000117448). One of these homologs, namely CG10863, encodes an enzyme known to share catalytic similarities to AKR1A1 and AKR1B1 [100]. This gene is under-expressed during cold acclimation, and D-Glucoronate is notably more abundant in cold acclimated flies [36]. Therefore, it was hypothesized that under the mentioned stress conditions, VC synthesis might be impaired by the reduced expression levels of both CG6910 (see above) and CG10863, the latter leading to the accumulation of D-Glucoronate in flies. Since, in contrast to Dca, that is overexpressed, Regucalcin is also under-expressed under cold acclimation conditions [36], when flies are placed in recovery conditions, along with the restoration of normal CG6910, CG10863 and Regucalcin expression levels, the role of Dca might be important for the rapid increase in VC synthesis, by providing a boost to the conversion of L-Gulonate to L-Gulono-1,4-lactone. Nevertheless, since Regucalcin and Dca are likely to perform important roles in calcium homeostasis as well [28], they may also have other crucial functions during cold exposure recovery. The regulation of calcium levels might be important for the maintenance of viable muscle function [36] in a manner unrelated to D. melanogaster VC synthesis. Thus, even if Dca does not intervene in VC metabolism, the reduced expression of CG10863 and CG6910 and Regucalcin under cold acclimation, and subsequent normalized expression on recovery conditions might be sufficient to explain the results obtained in [24]. While further studies on these genes may give insight into the VC synthesis pathway, there is still no candidate for the functional replacement of the GULO gene that has been lost in insects, such as D. melanogaster. Indeed, this species is a cholesterol auxotroph [101][102], that has lost the ability to convert desmosterol to cholesterol [103], likely due to the loss of the DHCR24 orthologous gene (the one involved in VC synthesis in B. mori [26]).
References
- Wheeler, G.; Ishikawa, T.; Pornsaksit, V.; Smirnoff, N. Evolution of alternative biosynthetic pathways for vitamin C following plastid acquisition in photosynthetic eukaryotes. eLife 2015, 4, e06369.
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The biosynthetic pathway of vitamin C in higher plants. Nature 1998, 393, 365–369.
- Shigeoka, S.; Nakano, Y.; Kitaoka, S. The biosynthetic pathway of L-ascorbic acid in Euglena gracilis Z. J. Nutr. Sci. Vitaminol. 1979, 25, 299–307.
- Nishikimi, M.; Yagi, K. Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis. Am. J. Clin. Nutr. 1991, 54, 1203S–1208S.
- Vargas, J.A.; Leonardo, D.A.; D’Muniz Pereira, H.; Lopes, A.R.; Rodriguez, H.N.; Cobos, M.; Marapara, J.L.; Castro, J.C.; Garratt, R.C. Structural Characterization of L-Galactose Dehydrogenase: An Essential Enzyme for Vitamin C Biosynthesis. Plant Cell Physiol. 2022, 63, 1140–1155.
- Gan, L.; Fan, H.; Nie, W.; Guo, Y. Ascorbic acid synthesis and transportation capacity in old laying hens and the effects of dietary supplementation with ascorbic acid. J. Anim. Sci. Biotechnol. 2018, 9, 71.
- Goggin, F.L.; Avila, C.A.; Lorence, A. Vitamin C content in plants is modified by insects and influences susceptibility to herbivory. BioEssays 2010, 32, 777–790.
- Ishikawa, T.; Shigeoka, S. Recent advances in ascorbate biosynthesis and the physiological significance of ascorbate peroxidase in photosynthesizing organisms. Biosci. Biotechnol. Biochem. 2008, 72, 1143–1154.
- Loewus, F.A. Biosynthesis and metabolism of ascorbic acid in plants and of analogs of ascorbic acid in fungi. Phytochemistry 1999, 52, 193–210.
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129.
- Zheng, X.; Gong, M.; Zhang, Q.; Tan, H.; Li, L.; Tang, Y.; Li, Z.; Peng, M.; Deng, W. Metabolism and Regulation of Ascorbic Acid in Fruits. Plants 2022, 11, 1602.
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519.
- Linster, C.L.; Van Schaftingen, E. Vitamin C: Biosynthesis, recycling and degradation in mammals. FEBS J. 2007, 274, 1–22.
- Linster, C.L.; Van Schaftingen, E. Glucuronate, the precursor of vitamin C, is directly formed from UDP-glucuronate in liver. FEBS J. 2006, 273, 1516–1527.
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453.
- Agius, F.; González-Lamothe, R.; Caballero, J.L.; Muñoz-Blanco, J.; Botella, M.A.; Valpuesta, V. Engineering increased Vitamin C levels in plants by overexpression of a D-galacturonic acid reductase. Nat. Biotechnol. 2003, 21, 177–181.
- Lorence, A.; Chevone, B.I.; Mendes, P.; Nessler, C.L. myo-Inositol Oxygenase Offers a Possible Entry Point into Plant Ascorbate Biosynthesis. Plant Physiol. 2004, 134, 1200–1205.
- Wolucka, B.A.; Van Montagu, M. GDP-mannose 3′,5′-epimerase forms GDP-L-gulose, a putative intermediate for the de novo biosynthesis of vitamin C in plants. J. Biol. Chem. 2003, 278, 47483–47490.
- Drouin, G.; Godin, J.R.; Pagé, B. The genetics of vitamin C loss in vertebrates. Curr. Genom. 2011, 12, 371–378.
- Yang, H. Conserved or Lost: Molecular Evolution of the Key Gene GULO in Vertebrate Vitamin C Biosynthesis. Biochem. Genet. 2013, 51, 413–425.
- Lachapelle, M.Y.; Drouin, G. Inactivation dates of the human and guinea pig vitamin C genes. Genetica 2011, 139, 199–207.
- López-Fernández, H.; Duque, P.; Henriques, S.; Vázquez, N.; Fdez-Riverola, F.; Vieira, C.P.; Reboiro-Jato, M.; Vieira, J. A bioinformatics protocol for quickly creating large-scale phylogenetic trees. In Practical Applications of Computational Biology and Bioinformatics, 12th International Conference; Fdez-Riverola, F., Mohamad, M.S., Rocha, M., De Paz, J.F., González, P., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 88–96.
- Chatterjee, I.B. Evolution and the Biosynthesis of Ascorbic Acid. Science 1973, 182, 1271–1272.
- Henriques, S.F.; Duque, P.; López-Fernández, H.; Vázquez, N.; Fdez-Riverola, F.; Reboiro-Jato, M.; Vieira, C.P.; Vieira, J. Multiple independent L-gulonolactone oxidase (GULO) gene losses and vitamin C synthesis reacquisition events in non-Deuterostomian animal species. BMC Evol. Biol. 2019, 19, 126.
- Patananan, A.N.; Budenholzer, L.M.; Pedraza, M.E.; Torres, E.R.; Adler, L.N.; Clarke, S.G. The invertebrate Caenorhabditis elegans biosynthesizes ascorbate. Arch. Biochem. Biophys. 2015, 569, 32–44.
- Hou, M.; Dai, T.M.; Liang, X.Y.; Zhang, S.X.; Cui, W.Z.; Qiu, J.F.; Sima, Y.H.; Cui, W.Z.; Xu, S.Q. Bombyx mori can synthesize ascorbic acid through the l-gulose pathway to varying degrees depending on developmental stage. Arch. Insect Biochem. Physiol. 2021, 106, e21783.
- Hänninen, O.; Raunio, R.; Marniemi, J. Biosynthesis of L-ascorbic acid from myo-inositol. Carbohydr. Res. 1971, 16, 343–351.
- Duque, P.; Vieira, C.P.; Bastos, B.; Vieira, J. The evolution of vitamin C biosynthesis and transport in animals. BMC Ecol. Evol. 2022, 22, 84.
- Kramer, K.J.; Seib, P.A. Ascorbic acid and the growth and development of insects. In Ascorbic Acid: Chemistry, Metabolism, and Uses; Advances in Chemistry; American Chemical Society: Washington, DC, USA, 1982; Volume 200, pp. 275–291.
- Massie, H.R.; Shumway, M.E.; Whitney, S.J.P.; Sternick, S.M.; Aiello, V.R. Ascorbic acid in Drosophila and changes during aging. Exp. Gerontol. 1991, 26, 487–494.
- Steinert, R.E.; Lee, Y.-K.; Sybesma, W. Vitamins for the Gut Microbiome. Trends Mol. Med. 2020, 26, 137–140.
- Pierre, L.L. Synthesis of Ascorbic Acid by the Normal Fat-Body of the Cockroach, Leucophaea maderae (F.), and by its Symbionts. Nature 1962, 193, 904–905.
- Travica, N.; Ried, K.; Hudson, I.; Sali, A.; Scholey, A.; Pipingas, A. The Contribution of Plasma and Brain Vitamin C on Age and Gender-Related Cognitive Differences: A Mini-Review of the Literature. Front. Integr. Neurosci. 2020, 14, 47.
- Aumailley, L.; Bourassa, S.; Gotti, C.; Droit, A.; Lebel, M. Vitamin C Differentially Impacts the Serum Proteome Profile in Female and Male Mice. J. Proteome Res. 2021, 20, 5036–5053.
- Kuo, S.-M.; MacLean, M.E.; McCormick, K.; Wilson, J.X. Gender and sodium-ascorbate transporter isoforms determine ascorbate concentrations in mice. J. Nutr. 2004, 134, 2216–2221.
- MacMillan, H.A.; Knee, J.M.; Dennis, A.B.; Udaka, H.; Marshall, K.E.; Merritt, T.J.S.; Sinclair, B.J. Cold acclimation wholly reorganizes the Drosophila melanogaster transcriptome and metabolome. Sci. Rep. 2016, 6, 28999.
- Arboleda-Bustos, C.E.; Segarra, C. The Dca gene involved in cold adaptation in Drosophila melanogaster arose by duplication of the ancestral regucalcin gene. Mol. Biol. Evol. 2011, 28, 2185–2195.
- Goto, S.G. Expression of Drosophila homologue of senescence marker protein-30 during cold acclimation. J. Insect Physiol. 2000, 46, 1111–1120.
- Lee, S.F.; Chen, Y.; Varan, A.K.; Wee, C.W.; Rako, L.; Axford, J.K.; Good, R.T.; Blacket, M.J.; Reuter, C.; Partridge, L. Molecular basis of adaptive shift in body size in Drosophila melanogaster: Functional and sequence analyses of the Dca gene. Mol. Biol. Evol. 2011, 28, 2393–2402.
- McKechnie, S.W.; Blacket, M.; Song, S.; Rako, L.; Carroll, X.; Johnson, T.; Jensen, L.; Lee, S.; Wee, C.; Hoffmann, A.A. A clinally varying promoter polymorphism associated with adaptive variation in wing size in Drosophila. Mol. Ecol. 2010, 19, 775–784.
- Qin, W.; Neal, S.J.; Robertson, R.M.; Westwood, J.T.; Walker, V.K. Cold hardening and transcriptional change in Drosophila melanogaster. Insect Mol. Biol. 2005, 14, 607–613.
- Sinclair, B.J.; Gibbs, A.G.; Roberts, S.P. Gene transcription during exposure to, and recovery from, cold and desiccation stress in Drosophila melanogaster. Insect Mol. Biol. 2007, 16, 435–443.
- Yamaguchi, M. The transcriptional regulation of regucalcin gene expression. Mol. Cell. Biochem. 2011, 346, 147–171.
- Tian, L.; Guo, E.; Wang, S.; Liu, S.; Jiang, R.-J.; Cao, Y.; Ling, E.; Li, S. Developmental Regulation of Glycolysis by 20-hydroxyecdysone and Juvenile Hormone in Fat Body Tissues of the Silkworm, Bombyx mori. J. Mol. Cell Biol. 2010, 2, 255–263.
- Gupta, S.D.; Choudhury, P.; Chatterjee, I. Synthesis of l-ascorbic acid from d-glucurono-1, 4-lactone conjugates by different species of animals. Int. J. Biochem. 1973, 4, 309–314.
- Ciufo, L.F.; Murray, P.A.; Thompson, A.; Rigden, D.J.; Rees, H.H. Characterisation of a desmosterol reductase involved in phytosterol dealkylation in the silkworm, Bombyx mori. PLoS ONE 2011, 6, e21316.
- Fujimori, H.; Zhou, Y.J.; Fukumura, K.; Matsumoto, S.; Tukamoto, Y.; Nagata, S. Specific distribution of expression and enzymatic activity of cholesterol biosynthetic enzyme DHCR24 orthologs in the phytophagous insect. Biosci. Biotechnol. Biochem. 2020, 84, 126–133.
- Ewing, T.A.; Fraaije, M.W.; Mattevi, A.; van Berkel, W.J. The VAO/PCMH flavoprotein family. Arch. Biochem. Biophys. 2017, 632, 104–117.
- Zerenturk, E.J.; Sharpe, L.J.; Ikonen, E.; Brown, A.J. Desmosterol and DHCR24: Unexpected new directions for a terminal step in cholesterol synthesis. Prog. Lipid Res. 2013, 52, 666–680.
- Takeshima, M.; Ogihara, M.H.; Kataoka, H. Characterization and functional analysis of BmSR-B1 for phytosterol uptake. Steroids 2022, 184, 109039.
- Leferink, N.G.H.; van Berkel, W.J.H. Aldonolactone Oxidoreductases. In Flavins and Flavoproteins: Methods and Protocols; Weber, S., Schleicher, E., Eds.; Springer: New York, NY, USA, 2014; pp. 95–111.
- Kanafi, R.R.; Ebadi, R.; Mirhosseini, S.; Seidavi, A.; Zolfaghari, M.; Etebari, K. A review on nutritive effect of mulberry leaves enrichment with vitamins on economic traits and biological parameters of silkworm Bombyx mori L. Invertebr. Surviv. J. 2007, 4, 86–91.
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412.
- Etebari, K.; Matindoost, L. Application of Multi-vitamins as Supplementary Nutrients on Biological and Economical Characteristics of Silkworm Bombyx mori L. J. Asia-Pac. Entomol. 2005, 8, 107–112.
- Vinci, G.; Xia, X.; Veitia, R.A. Preservation of Genes Involved in Sterol Metabolism in Cholesterol Auxotrophs: Facts and Hypotheses. PLoS ONE 2008, 3, e2883.
- Valpuesta, V.; Botella, M.A. Biosynthesis of L-ascorbic acid in plants: New pathways for an old antioxidant. Trends Plant Sci. 2004, 9, 573–577.
- Wolucka, B.A.; Van Montagu, M. The VTC2 cycle and the de novo biosynthesis pathways for vitamin C in plants: An opinion. Phytochemistry 2007, 68, 2602–2613.
- Yamauchi, N.; Nakayama, Y. Biosynthetic Mechanism for l-Gulose in Main Polar Lipids of Thermoplasma acidophilum and Possible Resemblance to Plant Ascorbic Acid Biosynthesis. Biosci. Biotechnol. Biochem. 2013, 77, 2087–2093.
- Major, L.L.; Wolucka, B.A.; Naismith, J.H. Structure and Function of GDP-Mannose-3‘,5‘-Epimerase: An Enzyme which Performs Three Chemical Reactions at the Same Active Site. J. Am. Chem. Soc. 2005, 127, 18309–18320.
- Beerens, K.; Gevaert, O.; Desmet, T. GDP-Mannose 3,5-Epimerase: A View on Structure, Mechanism, and Industrial Potential. Front. Mol. Biosci. 2021, 8, 784142.
- Goudsmit, E.M.; Neufeld, E.F. Formation of GDP-L-galactose from GDP-D-mannose. Biochem. Biophys. Res. Commun. 1967, 26, 730–735.
- Woodyer, R.D.; Christ, T.N.; Deweese, K.A. Single-step bioconversion for the preparation of L-gulose and L-galactose. Carbohydr. Res. 2010, 345, 363–368.
- Roos, C.; Kolmer, M.; Mattila, P.; Renkonen, R. Composition of Drosophila melanogaster proteome involved in fucosylated glycan metabolism. J. Biol. Chem. 2002, 277, 3168–3175.
- Thorsell, A.-G.; Persson, C.; Voevodskaya, N.; Busam, R.D.; Hammarström, M.; Gräslund, S.; Gräslund, A.; Hallberg, B.M. Structural and biophysical characterization of human myo-inositol oxygenase. J. Biol. Chem. 2008, 283, 15209–15216.
- Brown, P.M.; Caradoc-Davies, T.T.; Dickson, J.M.; Cooper, G.J.; Loomes, K.M.; Baker, E.N. Crystal structure of a substrate complex of myo-inositol oxygenase, a di-iron oxygenase with a key role in inositol metabolism. Proc. Natl. Acad. Sci. USA 2006, 103, 15032–15037.
- Chang, H.-H.; Chao, H.-N.; Walker, C.S.; Choong, S.-Y.; Phillips, A.; Loomes, K.M. Renal depletion of myo-inositol is associated with its increased degradation in animal models of metabolic disease. Am. J. Physiol.-Ren. Physiol. 2015, 309, F755–F763.
- Lu, Y.; Agarwal, A. Myo-inositol oxygenase in cadmium-induced kidney injury. Am. J. Physiol.-Ren. Physiol. 2022, 322, F470–F472.
- Oggioni, G. Experiments on the antiscorbutic action of inositol. Boll. Della Soc. Ital. Biol. Sper. 1953, 29, 1421–1422.
- Anderson, L.; Coots, R.H.; Halliday, J.W. A Critical Evaluation of Myo-Inositol as an Ascorbic Acid-Sparing Agent. J. Nutr. 1958, 64, 167–175.
- Kipp, D.E.; McElvain, M.; Kimmel, D.B.; Akhter, M.P.; Robinson, R.G.; Lukert, B.P. Scurvy results in decreased collagen synthesis and bone density in the guinea pig animal model. Bone 1996, 18, 281–288.
- Peterkofsky, B. Ascorbate requirement for hydroxylation and secretion of procollagen: Relationship to inhibition of collagen synthesis in scurvy. Am. J. Clin. Nutr. 1991, 54, 1135S–1140S.
- Chatterjee, I.B.; Gupta, S.D.; Majumder, A.K.; Nandi, B.K.; Subramanian, N. Effect of ascorbic acid on histamine metabolism in scorbutic guinea-pigs. J. Physiol. 1975, 251, 271–279.
- Freinkel, N.; Dawson, R.M.C.; Ingbar, S.H.; White, R.W. The Free Myo-Inositol of Thyroid Tissue. Proc. Soc. Exp. Biol. Med. 1959, 100, 549–551.
- Dawson, R.; Freinkel, N. The distribution of free mesoinositol in mammalian tissues, including some observations on the lactating rat. Biochem. J. 1961, 78, 606.
- Halver, J.E.; Hardy, R.W. L-Ascorbyl-2-Sulfate Alleviates Atlantic Salmon Scurvy. Proc. Soc. Exp. Biol. Med. 1994, 206, 421–424.
- Halver, J.E.; Smith, R.R.; Tolbert, B.M.; Baker, E.M. Utilization of ascorbic acid in fish. Ann. N. Y. Acad. Sci. 1975, 258, 81–102.
- Howard, C.F.; Anderson, L. Metabolism of myo-inositol in animals: II. Complete catabolism of myo-inositol-14C by rat kidney slices. Arch. Biochem. Biophys. 1967, 118, 332–339.
- Burns, J.; Evans, C. The synthesis of L-ascorbic acid in the rat from D-glucuronolactone and L-gulonolactone. J. Biol. Chem 1956, 223, 897–905.
- Bánhegyi, G.; Braun, L.; Csala, M.; Puskás, F.; Mandl, J. Ascorbate Metabolism and Its Regulation in Animals. Free Radic. Biol. Med. 1997, 23, 793–803.
- Jenness, R.; Birney, E.C.; Ayaz, K.L.; Buzzell, D.M. Ontogenetic development of l-gulonolactone oxidase activity in several vertebrates. Comp. Biochem. Physiol. Part B Comp. Biochem. 1984, 78, 167–173.
- Walter Bock, K.; Köhle, C. UDP-Glucuronosyltransferase 1A6: Structural, Functional, and Regulatory Aspects. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2005; Volume 400, pp. 57–75.
- Strassburg, C.P.; Strassburg, A.; Kneip, S.; Barut, A.; Tukey, R.H.; Rodeck, B.; Manns, M.P. Developmental aspects of human hepatic drug glucuronidation in young children and adults. Gut 2002, 50, 259.
- Kiani, A.; Paolacci, S.; Calogero, A.; Cannarella, R.; Di Renzo, G.; Gerli, S.; Della Morte, C.; Busetto, G.; De Berardinis, E.; Del Giudice, F. From Myo-inositol to D-chiro-inositol molecular pathways. Eur. Rev. Med. Pharmacol. Sci 2021, 25, 2390.
- Arner, R.J.; Prabhu, K.S.; Reddy, C.C. Molecular cloning, expression, and characterization of myo-inositol oxygenase from mouse, rat, and human kidney. Biochem. Biophys. Res. Commun. 2004, 324, 1386–1392.
- Sochor, M.; Baquer, N.Z.; McLean, P. Regulation of pathways of glucose metabolism in kidney. The effect of experimental diabetes on the activity of the pentose phosphate pathway and the glucuronate-xylulose pathway. Arch. Biochem. Biophys. 1979, 198, 632–646.
- Barski, O.A.; Papusha, V.Z.; Ivanova, M.M.; Rudman, D.M.; Finegold, M.J. Developmental expression and function of aldehyde reductase in proximal tubules of the kidney. Am. J. Physiol.-Ren. Physiol. 2005, 289, F200–F207.
- Agrawal, N.; Hossain, M.S.; Skelton, A.A.; Muralidhar, K.; Kaushik, S. Unraveling the mechanism of l-gulonate-3-dehydrogenase inhibition by ascorbic acid: Insights from molecular modeling. Comput. Biol. Chem. 2018, 77, 146–153.
- Birney, E.C.; Jenness, R.; Hume, I.D. Ascorbic acid biosynthesis in the mammalian kidney. Experientia 1979, 35, 1425–1426.
- Chaudhuri, C.R.; Chatterjee, I.B. L-Ascorbic Acid Synthesis in Birds: Phylogenetic Trend. Science 1969, 164, 435–436.
- Nishikimi, M.; Yagi, K. Biochemistry and Molecular Biology of Ascorbic Acid Biosynthesis. In Subcellular Biochemistry: Ascorbic Acid: Biochemistry and Biomedical Cell Biology; Harris, J.R., Ed.; Springer: Boston, MA, USA, 1996; pp. 17–39.
- Roy, R.N.; Guha, B.C. Species Difference in regard to the Biosynthesis of Ascorbic Acid. Nature 1958, 182, 319–320.
- Dabrowski, K. Primitive actimoterigian fishes can synthesize ascorbic acid. Experientia 1994, 50, 745–748.
- White, R.E.; Evans, D.C.; Hop, C.E.C.A.; Moore, D.J.; Prakash, C.; Surapaneni, S.; Tse, F.L.S. Radiolabeled mass-balance excretion and metabolism studies in laboratory animals: A commentary on why they are still necessary. Xenobiotica 2013, 43, 219–225.
- Chitwood, D.J. Biochemistry and Function of Nematode Steroids. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 273–284.
- Waterham, H.R.; Koster, J.; Romeijn, G.J.; Hennekam, R.C.M.; Vreken, P.; Andersson, H.C.; FitzPatrick, D.R.; Kelley, R.I.; Wanders, R.J.A. Mutations in the 3β-Hydroxysterol Δ24-Reductase Gene Cause Desmosterolosis, an Autosomal Recessive Disorder of Cholesterol Biosynthesis. Am. J. Hum. Genet. 2001, 69, 685–694.
- Jones, M.K.; Eldon, E.D.; Klig, L.S. myo-Inositol oxygenase identified in Drosophila melanogaster. FASEB J. 2012, 26, 759.13.
- Lai, C.W.; Chen, H.L.; Tu, M.Y.; Lin, W.Y.; Röhrig, T.; Yang, S.H.; Lan, Y.W.; Chong, K.Y.; Chen, C.M. A novel osteoporosis model with ascorbic acid deficiency in Akr1A1 gene knockout mice. Oncotarget 2017, 8, 7357–7369.
- Gabbay, K.H.; Bohren, K.M.; Morello, R.; Bertin, T.; Liu, J.; Vogel, P. Ascorbate synthesis pathway: Dual role of ascorbate in bone homeostasis. J. Biol. Chem. 2010, 285, 19510–19520.
- O’Connor, T.; Ireland, L.S.; Harrison, D.J.; Hayes, J.D. Major differences exist in the function and tissue-specific expression of human aflatoxin B1 aldehyde reductase and the principal human aldo-keto reductase AKR1 family members. Biochem. J. 1999, 343, 487–504.
- Di Luccio, E.; Elling, R.A.; Wilson, D.K. Identification of a novel NADH-specific aldo-keto reductase using sequence and structural homologies. Biochem. J. 2006, 400, 105–114.
- Jing, X.; Behmer, S.T. Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu. Rev. Entomol. 2020, 65, 251–271.
- Lavrynenko, O.; Rodenfels, J.; Carvalho, M.; Dye, N.A.; Lafont, R.; Eaton, S.; Shevchenko, A. The ecdysteroidome of Drosophila: Influence of diet and development. Development 2015, 142, 3758–3768.
- Svoboda, J.A.; Feldlaufer, M.F. Neutral sterol metabolism in insects. Lipids 1991, 26, 614–618.
More
Information
Subjects:
Evolutionary Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.6K
Revisions:
3 times
(View History)
Update Date:
04 Nov 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No