 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Geou-Yarh Liou | -- | 3564 | 2022-10-25 03:22:44 |
Video Upload Options
Protein tyrosine kinase Bruton’s tyrosine kinase (BTK) is essential for B cell maturation and proliferation. Dysregulation of BTK signaling in B cells leads to B cell lymphoma. In addition to B cells, BTK is also expressed in other types of immune cells including myeloid derived suppressor cells (MDSC), dendritic cells, mast cells and macrophages, all of which comprise the tumor microenvironment in solid cancers.
1. Introduction
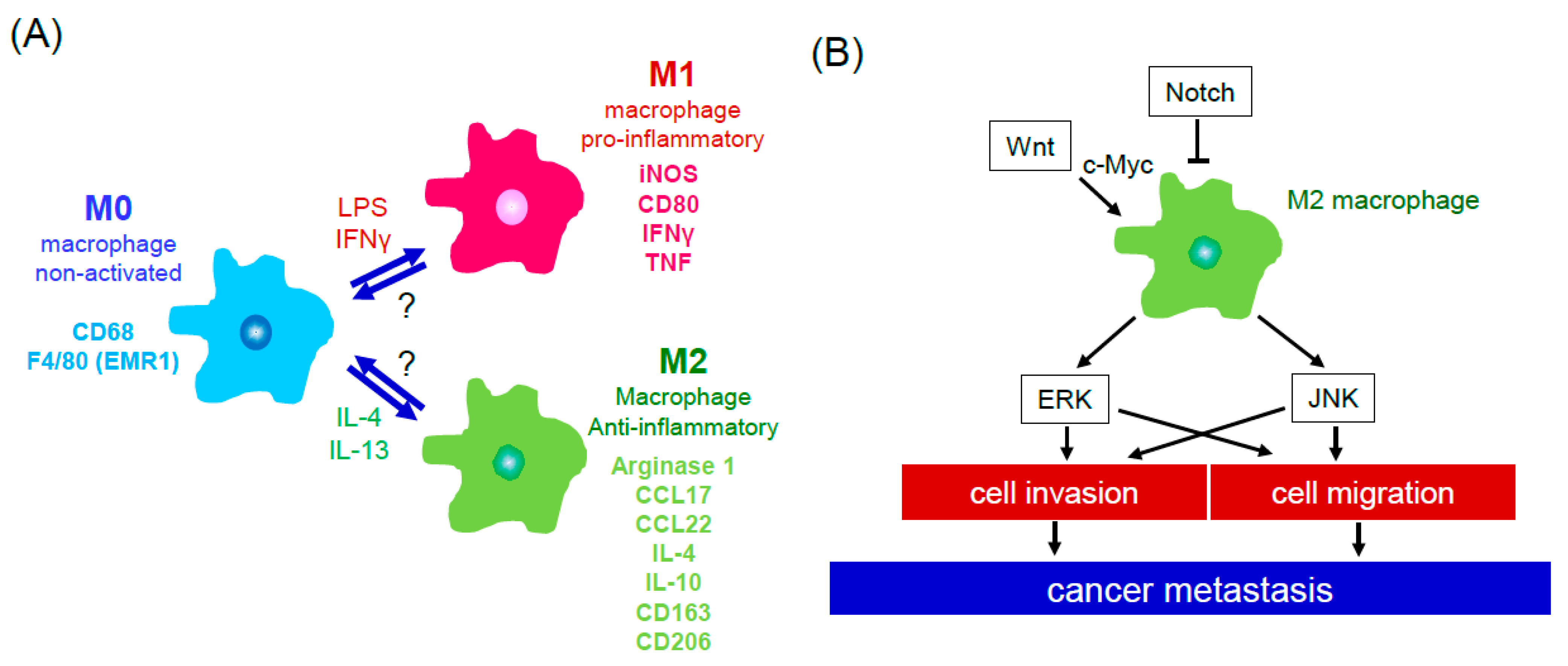
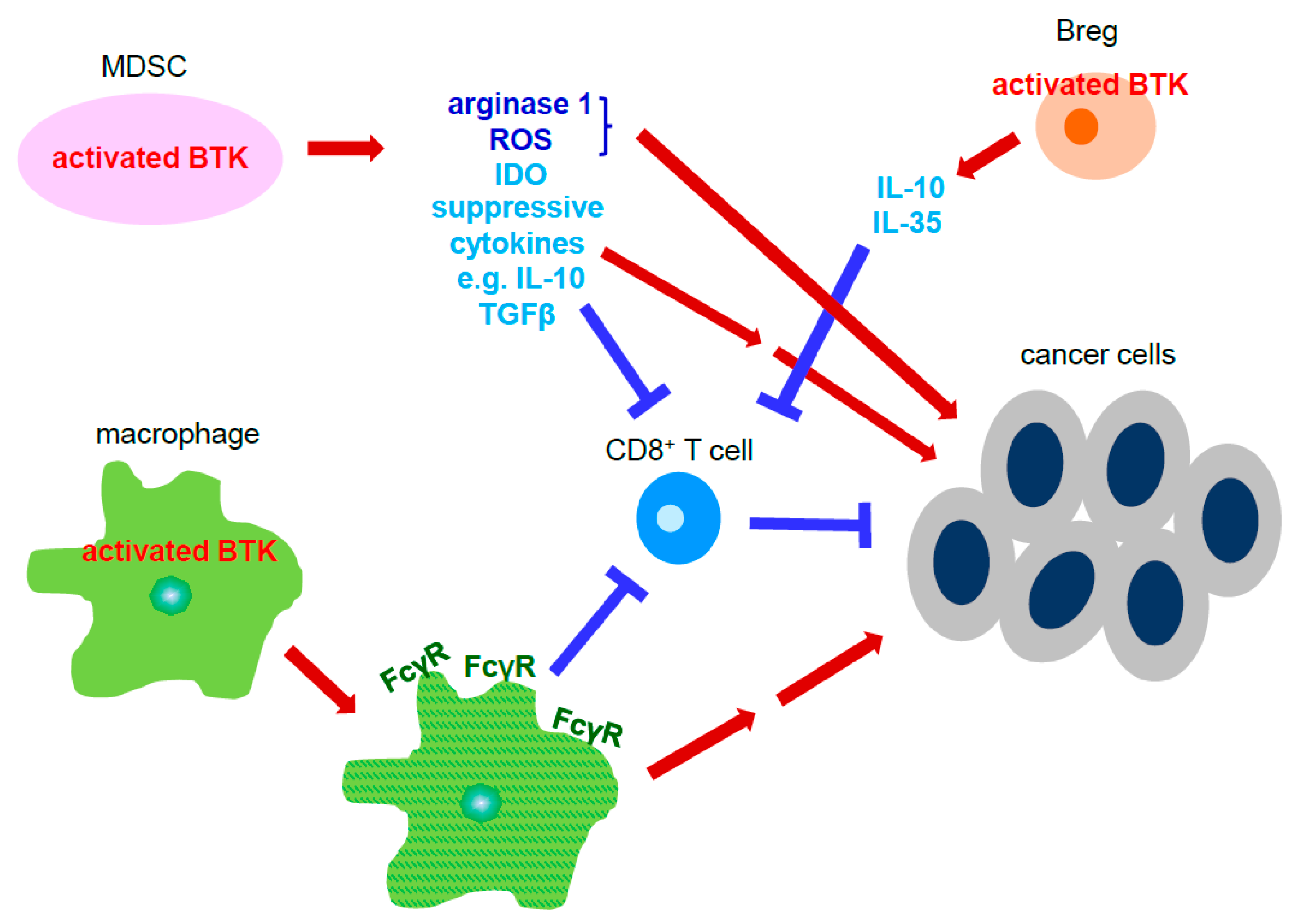
2. Cells of the Tumor Microenvironment
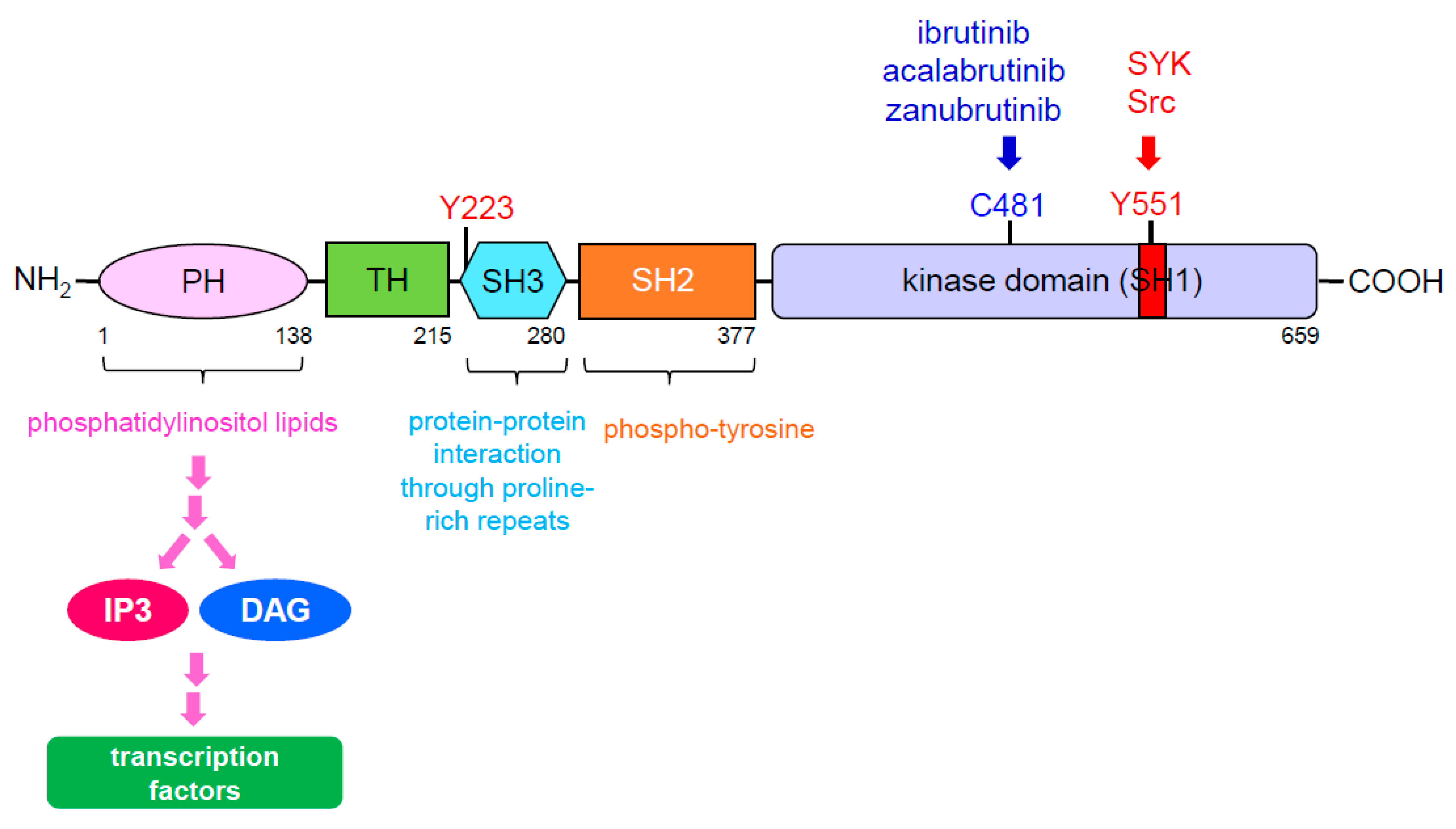
3. BTK Signaling
|
BTK Inhibitor |
Study Type |
Cancer Type |
|---|---|---|
|
ibrutinib |
in vitro |
B cell lymphoma |
|
chronic lymphocytic leukemia |
||
|
pancreatic cancer |
||
|
in vivo |
chronic lymphocytic leukemia |
|
|
pancreatic cancer |
||
|
colorectal cancer |
||
|
breast cancer |
||
|
acalabrutinib |
in vitro |
chronic lymphocytic leukemia |
|
in vivo |
B cell lymphoma |
|
|
chronic lymphocytic leukemia |
||
|
pancreatic cancer * |
||
|
urothelial cancer * |
||
|
tirabrutinib |
in vivo |
pancreatic cancer |
|
zanubrutinib |
in vivo |
mantle cell lymphoma * |
4. BTK Inhibitors and Their Clinic Application for Cancer Therapy
References
- Catalano, V.; Turdo, A.; di Franco, S.; Dieli, F.; Todaro, M.; Stassi, G. Tumor and Its Microenvironment: A Synergistic Interplay. In Seminars in Cancer Biology; Elsevier: London, UK, 2013; Volume 23, pp. 522–532.
- Ozdemir, B.C.; Pentcheva-Hoang, T.; Carstens, J.L.; Zheng, X.; Wu, C.C.; Simpson, T.R.; Laklai, H.; Sugimoto, H.; Kahlert, C.; Novitskiy, S.V.; et al. Depletion of Carcinoma-Associated Fibroblasts and Fibrosis Induces Immunosuppression and Accelerates Pancreas Cancer with Reduced Survival. Cancer Cell 2014, 25, 719–734.
- Whiteside, T.L. The Tumor Microenvironment and Its Role in Promoting Tumor Growth. Oncogene 2008, 27, 5904–5912.
- Mittendorf, E.A.; Philips, A.V.; Meric-Bernstam, F.; Qiao, N.; Wu, Y.; Harrington, S.; Su, X.; Wang, Y.; Gonzalez-Angulo, A.M.; Akcakanat, A.; et al. PD-L1 Expression in Triple-Negative Breast Cancer. Cancer Immunol. Res. 2014, 2, 361–370.
- Vetrie, D.; Vorechovsky, I.; Sideras, P.; Holland, J.; Davies, A.; Flinter, F.; Hammarstrom, L.; Kinnon, C.; Levinsky, R.; Bobrow, M.; et al. The Gene Involved in X-linked Agammaglobulinaemia is a Member of the SRC Family of Protein-Tyrosine Kinases. Nature 1993, 361, 226–233.
- Tsukada, S.; Saffran, D.C.; Rawlings, D.J.; Parolini, O.; Allen, R.C.; Klisak, I.; Sparkes, R.S.; Kubagawa, H.; Mohandas, T.; Quan, S.; et al. Deficient Expression of a B Cell Cytoplasmic Tyrosine Kinase in Human X-linked Agammaglobulinemia. Cell 1993, 72, 279–290.
- Bauer, J.; Emon, M.A.B.; Staudacher, J.J.; Thomas, A.L.; Zessner-Spitzenberg, J.; Mancinelli, G.; Krett, N.; Saif, M.T.; Jung, B. Increased Stiffness of the Tumor Microenvironment in Colon Cancer Stimulates Cancer Associated Fibroblast-Mediated Prometastatic Activin a Signaling. Sci. Rep. 2020, 10, 50.
- Liu, C.; Pei, H.; Tan, F. Matrix Stiffness and Colorectal Cancer. OncoTargets Ther. 2020, 13, 2747–2755.
- Day, S.D.; Enos, R.T.; McClellan, J.L.; Steiner, J.L.; Velazquez, K.T.; Murphy, E.A. Linking Inflammation to Tumorigenesis in a Mouse Model of High-Fat-Diet-Enhanced Colon Cancer. Cytokine 2013, 64, 454–462.
- Kern, L.; Mittenbuhler, M.J.; Vesting, A.J.; Ostermann, A.L.; Wunderlich, C.M.; Wunderlich, F.T. Obesity-Induced TNFalpha and IL-6 Signaling: The Missing Link between Obesity and Inflammation-Driven Liver and Colorectal Cancers. Cancers 2018, 11, 24.
- Barcellos-de-Souza, P.; Comito, G.; Pons-Segura, C.; Taddei, M.L.; Gori, V.; Becherucci, V.; Bambi, F.; Margheri, F.; Laurenzana, A.; Del Rosso, M.; et al. Mesenchymal Stem Cells are Recruited and Activated into Carcinoma-Associated Fibroblasts by Prostate Cancer Microenvironment-Derived TGF-beta1. Stem Cells 2016, 34, 2536–2547.
- Xu, W.; Qian, J.; Zeng, F.; Li, S.; Guo, W.; Chen, L.; Li, G.; Zhang, Z.; Wang, Q.J.; Deng, F.; et al. Protein Kinase Ds Promote Tumor Angiogenesis through Mast Cell Recruitment and Expression of Angiogenic Factors in Prostate Cancer Microenvironment. J. Exp. Clin. Cancer Res. 2019, 38, 114.
- Zhou, H.; Liu, T.; Wang, Z. Analysis of Non-Small Cell Lung Cancer Microenvironment Indicates Preponderance of T Cell Exhaustion Marker Expression. Exp. Cell Res. 2017, 360, 205–209.
- Kwiecien, I.; Stelmaszczyk-Emmel, A.; Polubiec-Kownacka, M.; Dziedzic, D.; Domagala-Kulawik, J. Elevated Regulatory T Cells, Surface and Intracellular CTLA-4 Expression and Interleukin-17 in the Lung Cancer Microenvironment in Humans. Cancer Immunol. Immunother. 2017, 66, 161–170.
- Faget, J.; Groeneveld, S.; Boivin, G.; Sankar, M.; Zangger, N.; Garcia, M.; Guex, N.; Zlobec, I.; Steiner, L.; Piersigilli, A.; et al. Neutrophils and Snail Orchestrate the Establishment of a Pro-tumor Microenvironment in Lung Cancer. Cell Rep. 2017, 21, 3190–3204.
- Chen, P.C.; Cheng, H.C.; Wang, J.; Wang, S.W.; Tai, H.C.; Lin, C.W.; Tang, C.H. Prostate Cancer-Derived CCN3 Induces M2 Macrophage Infiltration and Contributes to Angiogenesis in Prostate Cancer Microenvironment. Oncotarget 2014, 5, 1595–1608.
- Mantovani, A.; Germano, G.; Marchesi, F.; Locatelli, M.; Biswas, S.K. Cancer-Promoting Tumor-Associated Macrophages: New Vistas and Open Questions. Eur. J. Immunol. 2011, 41, 2522–2525.
- Mantovani, A.; Schioppa, T.; Porta, C.; Allavena, P.; Sica, A. Role of Tumor-Associated Macrophages in Tumor Progression and Invasion. Cancer Metastasis Rev. 2006, 25, 315–322.
- Puig-Kroger, A.; Sierra-Filardi, E.; Dominguez-Soto, A.; Samaniego, R.; Corcuera, M.T.; Gomez-Aguado, F.; Ratnam, M.; Sanchez-Mateos, P.; Corbi, A.L. Folate Receptor Beta is Expressed by Tumor-Associated Macrophages and Constitutes a Marker for M2 Anti-Inflammatory/Regulatory Macrophages. Cancer Res. 2009, 69, 9395–9403.
- De Robertis, M.; Massi, E.; Poeta, M.L.; Carotti, S.; Morini, S.; Cecchetelli, L.; Signori, E.; Fazio, V.M. The AOM/DSS murine model for the study of colon carcinogenesis: From pathways to diagnosis and therapy studies. J. Carcinog. 2011, 10, 9.
- Dong, P.; Ma, L.; Liu, L.; Zhao, G.; Zhang, S.; Dong, L.; Xue, R.; Chen, S. CD86(+)/CD206(+), Diametrically Polarized Tumor-Associated Macrophages, Predict Hepatocellular Carcinoma Patient Prognosis. Int. J. Mol. Sci. 2016, 17, 320.
- Gabhann, J.; Hams, E.; Smith, S.; Wynne, C.; Byrne, J.C.; Brennan, K.; Spence, S.; Kissenpfennig, A.; Johnston, J.A.; Fallon, P.G.; et al. Btk Regulates Macrophage Polarization in Response to Lipopolysaccharide. PLoS ONE 2014, 9, e85834.
- Singh, S.P.; Dammeijer, F.; Hendriks, R.W. Role of Bruton’s Tyrosine Kinase in B Cells and Malignancies. Mol. Cancer 2018, 17, 57.
- Park, H.; Wahl, M.I.; Afar, D.E.; Turck, C.W.; Rawlings, D.J.; Tam, C.; Scharenberg, A.M.; Kinet, J.P.; Witte, O.N. Regulation of Btk Function by a Major Autophosphorylation Site within the SH3 Domain. Immunity 1996, 4, 515–525.
- Rawlings, D.J.; Scharenberg, A.M.; Park, H.; Wahl, M.I.; Lin, S.; Kato, R.M.; Fluckiger, A.-C.; Witte, O.N.; Kinet, J.-P. Activation of BTK by a Phosphorylation Mechanism Initiated by SRC Family Kinases. Science 1996, 271, 822–825.
- Solvason, N.; Wu, W.W.; Kabra, N.; Lund-Johansen, F.; Roncarolo, M.G.; Behrens, T.W.; Grillot, D.A.; Nunez, G.; Lees, E.; Howard, M.; et al. Transgene Expression of bcl-xL Permits Anti-Immunoglobulin (Ig)-Induced Proliferation in Xid B Cells. J. Exp. Med. 1998, 187, 1081–1091.
- Craxton, A.; Jiang, A.; Kurosaki, T.; Clark, E.A. Syk and Bruton’s Tyrosine Kinase are Required for B Cell Antigen Receptor-Mediated Activation of the Kinase Akt. J. Biol. Chem. 1999, 274, 30644–30650.
- Okada, T.; Ngo, V.N.; Ekland, E.H.; Forster, R.; Lipp, M.; Littman, D.R.; Cyster, J.G. Chemokine Requirements for B Cell Entry to Lymph Nodes and Peyer’s Patches. J. Exp. Med. 2002, 196, 65–75.
- De Gorter, D.J.; Beuling, E.A.; Kersseboom, R.; Middendorp, S.; van Gils, J.M.; Hendriks, R.W.; Pals, S.T.; Spaargaren, M. Bruton’s Tyrosine Kinase and Phospholipase Cgamma2 Mediate Chemokine-Controlled B Cell Migration and Homing. Immunity 2007, 26, 93–104.
- Alugupalli, K.R.; Akira, S.; Lien, E.; Leong, J.M. MyD88-and Bruton’s Tyrosine Kinase-Mediated Signals are Essential for T Cell-Independent Pathogen-Specific IgM Responses. J. Immunol. 2007, 178, 3740–3749.
- Mitsuiki, N.; Yang, X.; Bartol, S.J.; Grosserichter-Wagener, C.; Kosaka, Y.; Takada, H.; Imai, K.; Kanegane, H.; Mizutani, S.; van der Burg, M.; et al. Mutations in Bruton’s Tyrosine Kinase Impair IgA Responses. Int. J. Hematol. 2015, 101, 305–313.
- Ng, Y.S.; Wardemann, H.; Chelnis, J.; Cunningham-Rundles, C.; Meffre, E. Bruton’s Tyrosine Kinase is Essential for Human B Cell Tolerance. J. Exp. Med. 2004, 200, 927–934.
- Rajaiya, J.; Hatfield, M.; Nixon, J.C.; Rawlings, D.J.; Webb, C.F. Bruton’s Tyrosine Kinase Regulates Immunoglobulin Promoter Activation in Association with the Transcription Factor Bright. Mol. Cell Biol. 2005, 25, 2073–2084.
- Mundy-Bosse, B.L.; Lesinski, G.B.; Jaime-Ramirez, A.C.; Benninger, K.; Khan, M.; Kuppusamy, P.; Guenterberg, K.; Kondadasula, S.V.; Chaudhury, A.R.; La Perle, K.M.; et al. Myeloid-Derived Suppressor Cell Inhibition of the IFN Response in Tumor-Bearing Mice. Cancer Res. 2011, 71, 5101–5110.
- Gunderson, A.J.; Kaneda, M.M.; Tsujikawa, T.; Nguyen, A.V.; Affara, N.I.; Ruffell, B.; Gorjestani, S.; Liudahl, S.M.; Truitt, M.; Olson, P.; et al. Bruton Tyrosine Kinase-Dependent Immune Cell Cross-talk Drives Pancreas Cancer. Cancer Discov. 2016, 6, 270–285.
- Das, S.; Bar-Sagi, D. BTK Signaling Drives CD1d(hi)CD5(+) Regulatory B-Cell Differentiation to Promote Pancreatic Carcinogenesis. Oncogene 2019, 38, 3316–3324.
- Kawakami, Y.; Inagaki, N.; Salek-Ardakani, S.; Kitaura, J.; Tanaka, H.; Nagao, K.; Kawakami, Y.; Xiao, W.; Nagai, H.; Croft, M.; et al. Regulation of Dendritic Cell Maturation and Function by Bruton’s Tyrosine Kinase via IL-10 and Stat3. Proc. Natl. Acad. Sci. USA 2006, 103, 153–158.
- Natarajan, G.; Oghumu, S.; Terrazas, C.; Varikuti, S.; Byrd, J.C.; Satoskar, A.R. A Tec Kinase BTK Inhibitor Ibrutinib Promotes Maturation and Activation of Dendritic Cells. Oncoimmunology 2016, 5, e1151592.
- Johnson, A.R.; Kohli, P.B.; Katewa, A.; Gogol, E.; Belmont, L.D.; Choy, R.; Penuel, E.; Burton, L.; Eigenbrot, C.; Yu, C.; et al. Battling Btk Mutants With Noncovalent Inhibitors That Overcome Cys481 and Thr474 Mutations. ACS Chem. Biol. 2016, 11, 2897–2907.
- Wu, J.; Liu, C.; Tsui, S.T.; Liu, D. Second-Generation Inhibitors of Bruton Tyrosine Kinase. J. Hematol. Oncol. 2016, 9, 80.
- Brown, J.R. Ibrutinib (PCI-32765), the First BTK (Bruton’s Tyrosine Kinase) Inhibitor in Clinical Trials. Curr. Hematol. Malig. Rep. 2013, 8, 1–6.
- Burger, J.A.; Buggy, J.J. Bruton Tyrosine Kinase Inhibitor Ibrutinib (PCI-32765). Leuk. Lymphoma 2013, 54, 2385–2391.
- Sivina, M.; Kreitman, R.J.; Arons, E.; Ravandi, F.; Burger, J.A. The Bruton Tyrosine Kinase Inhibitor Ibrutinib (PCI-32765) Blocks Hairy Cell Leukaemia Survival, Proliferation and B Cell Receptor Signalling: A New Therapeutic Approach. Br. J. Haematol. 2014, 166, 177–188.
- Honigberg, L.A.; Smith, A.M.; Sirisawad, M.; Verner, E.; Loury, D.; Chang, B.; Li, S.; Pan, Z.; Thamm, D.H.; Miller, R.A.; et al. The Bruton Tyrosine Kinase Inhibitor PCI-32765 Blocks B-Cell Activation and is Efficacious in Models of Autoimmune Disease and B-Cell Malignancy. Proc. Natl. Acad. Sci. USA 2010, 107, 13075–13080.
- Herman, S.E.; Gordon, A.L.; Hertlein, E.; Ramanunni, A.; Zhang, X.; Jaglowski, S.; Flynn, J.; Jones, J.; Blum, K.A.; Buggy, J.J.; et al. Bruton Tyrosine Kinase Represents a Promising Therapeutic Target for Treatment of Chronic Lymphocytic Leukemia and is Effectively Targeted by PCI-32765. Blood 2011, 117, 6287–6296.
- De Rooij, M.F.; Kuil, A.; Geest, C.R.; Eldering, E.; Chang, B.Y.; Buggy, J.J.; Pals, S.T.; Spaargaren, M. The Clinically Active BTK Inhibitor PCI-32765 Targets B-Cell Receptor-and Chemokine-Controlled Adhesion and Migration in Chronic Lymphocytic Leukemia. Blood 2012, 119, 2590–2594.
- Ponader, S.; Chen, S.S.; Buggy, J.J.; Balakrishnan, K.; Gandhi, V.; Wierda, W.G.; Keating, M.J.; O’Brien, S.; Chiorazzi, N.; Burger, J.A.; et al. The Bruton Tyrosine Kinase Inhibitor PCI-32765 Thwarts Chronic Lymphocytic Leukemia Cell Survival and Tissue Homing in Vitro and in Vivo. Blood 2012, 119, 1182–1189.
- Thorp, B.C.; Badoux, X. Atrial Fibrillation as a Complication of Ibrutinib Therapy: Clinical Features and Challenges of Management. Leuk. Lymphoma. 2018, 59, 311–320.
- Kriegsmann, K.; Kriegsmann, M.; Witzens-Harig, M. Acalabrutinib, A Second-Generation Bruton’s Tyrosine Kinase Inhibitor. Recent Results Cancer Res. 2018, 212, 285–294.
- Wu, J.; Zhang, M.; Liu, D. Acalabrutinib (ACP-196): A Selective Second-Generation BTK Inhibitor. J. Hematol. Oncol. 2016, 9, 21.
- Byrd, J.C.; Harrington, B.; O’Brien, S.; Jones, J.A.; Schuh, A.; Devereux, S.; Chaves, J.; Wierda, W.G.; Awan, F.T.; Brown, J.R.; et al. Acalabrutinib (ACP-196) in Relapsed Chronic Lymphocytic Leukemia. N. Engl. J. Med. 2016, 374, 323–332.
- Heather, B.K.H.; Gardner, L.; Raquel, I.; Ahmed, H.; Allard, K.; Bart, V.L.; Cheryl, A.; London, J.C.B.; Amy, J.J.; William, C.K. ACP-196: A Second Generation Btk Inhibitor Demonstrates Biological Activity in a Canine Model of B-Cell Non-Hodgkin Lymphoma. In Proceedings of the 105th Annual Meeting of the American Association for Cancer Research, San Diego, CA, USA, 5–9 April 2014; Volume 74.
- Niemann, C.U.; Montraveta, A.; Herman, S.E.M.; Ingallinera, T.; Barf, T.; Colomer, D.; Wiestner, A. The Novel Bruton’s Tyrosine Kinase Inhibitor ACP-196 Shows in Vivo Efficacy Against Human Chronic Lymphocytic Leukemia Cells Xenografted to the NSG Mouse Model. Cancer Res. 2014, 74, 2624.
- Herman, S.E.; Sun, X.; McAuley, E.M.; Hsieh, M.M.; Pittaluga, S.; Raffeld, M.; Liu, D.; Keyvanfar, K.; Chapman, C.M.; Chen, J.; et al. Modeling Tumor-Host Interactions of Chronic Lymphocytic Leukemia in Xenografted Mice to Study Tumor Biology and Evaluate Targeted Therapy. Leukemia 2013, 27, 2311–2321.
- Herman, S.E.M.; Montraveta, A.; Niemann, C.U.; Mora-Jensen, H.; Gulrajani, M.; Krantz, F.; Mantel, R.; Smith, L.L.; McClanahan, F.; Harrington, B.K.; et al. The Bruton Tyrosine Kinase (BTK) Inhibitor Acalabrutinib Demonstrates Potent On-Target Effects and Efficacy in Two Mouse Models of Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2017, 23, 2831–2841.
- Johnson, A.J.; Lucas, D.M.; Muthusamy, N.; Smith, L.L.; Edwards, R.B.; De Lay, M.D.; Croce, C.M.; Grever, M.R.; Byrd, J.C. Characterization of the TCL-1 Transgenic Mouse as a Preclinical Drug Development Tool for Human Chronic Lymphocytic Leukemia. Blood 2006, 108, 1334–1338.
- Skarzynski, M.; Niemann, C.U.; Lee, Y.S.; Martyr, S.; Maric, I.; Salem, D.; Stetler-Stevenson, M.; Marti, G.E.; Calvo, K.R.; Yuan, C.; et al. Interactions between Ibrutinib and Anti-CD20 Antibodies: Competing Effects on the Outcome of Combination Therapy. Clin. Cancer Res. 2016, 22, 86–95.
- Varikuti, S.; Singh, B.; Volpedo, G.; Ahirwar, D.K.; Jha, B.K.; Saljoughian, N.; Viana, A.G.; Verma, C.; Hamza, O.; Halsey, G.; et al. Ibrutinib Treatment Inhibits Breast Cancer Progression and Metastasis by Inducing Conversion of Myeloid-Derived Suppressor Cells to Dendritic Cells. Br. J. Cancer 2020, 122, 1005–1013.
- Hong, D.; Rasco, D.; Veeder, M.; Luke, J.J.; Chandler, J.; Balmanoukian, A.; George, T.J.; Munster, P.; Berlin, J.D.; Gutierrez, M.; et al. A Phase 1b/2 Study of the Bruton Tyrosine Kinase Inhibitor Ibrutinib and the PD-L1 Inhibitor Durvalumab in Patients with Pretreated Solid Tumors. Oncology 2019, 97, 102–111.
- Overman, M.; Javle, M.; Davis, R.E.; Vats, P.; Kumar-Sinha, C.; Xiao, L.; Mettu, N.B.; Parra, E.R.; Benson, A.B.; Lopez, C.D.; et al. Randomized Phase II Study of the Bruton Tyrosine Kinase Inhibitor Acalabrutinib, Alone or with Pembrolizumab in Patients with Advanced Pancreatic Cancer. J. Immunother. Cancer 2020, 8.
- Tempero, M.; Oh, D.Y.; Tabernero, J.; Reni, M.; Van Cutsem, E.; Hendifar, A.; Waldschmidt, D.T.; Starling, N.; Bachet, J.B.; Chang, H.M.; et al. Ibrutinib in Combination with Nab-Paclitaxel and Gemcitabine for First-Line Treatment of Patients with Metastatic Pancreatic Adenocarcinoma: Phase III RESOLVE Study. Ann. Oncol. 2021, 32, 600–608.
- Zhang, T.; Harrison, M.R.; O’Donnell, P.H.; Alva, A.S.; Hahn, N.M.; Appleman, L.J.; Cetnar, J.; Burke, J.M.; Fleming, M.T.; Milowsky, M.I.; et al. A Randomized Phase 2 Trial of Pembrolizumab Versus Pembrolizumab and Acalabrutinib in Patients with Platinum-Resistant Metastatic Urothelial Cancer. Cancer 2020, 126, 4485–4497.

