 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | wei wang | -- | 2967 | 2022-10-20 04:08:15 | | | |
| 2 | Beatrix Zheng | Meta information modification | 2967 | 2022-10-21 02:59:16 | | | | |
| 3 | wei wang | Meta information modification | 2967 | 2022-10-31 10:09:14 | | | | |
| 4 | Beatrix Zheng | Meta information modification | 2967 | 2022-11-02 03:06:18 | | |
Video Upload Options
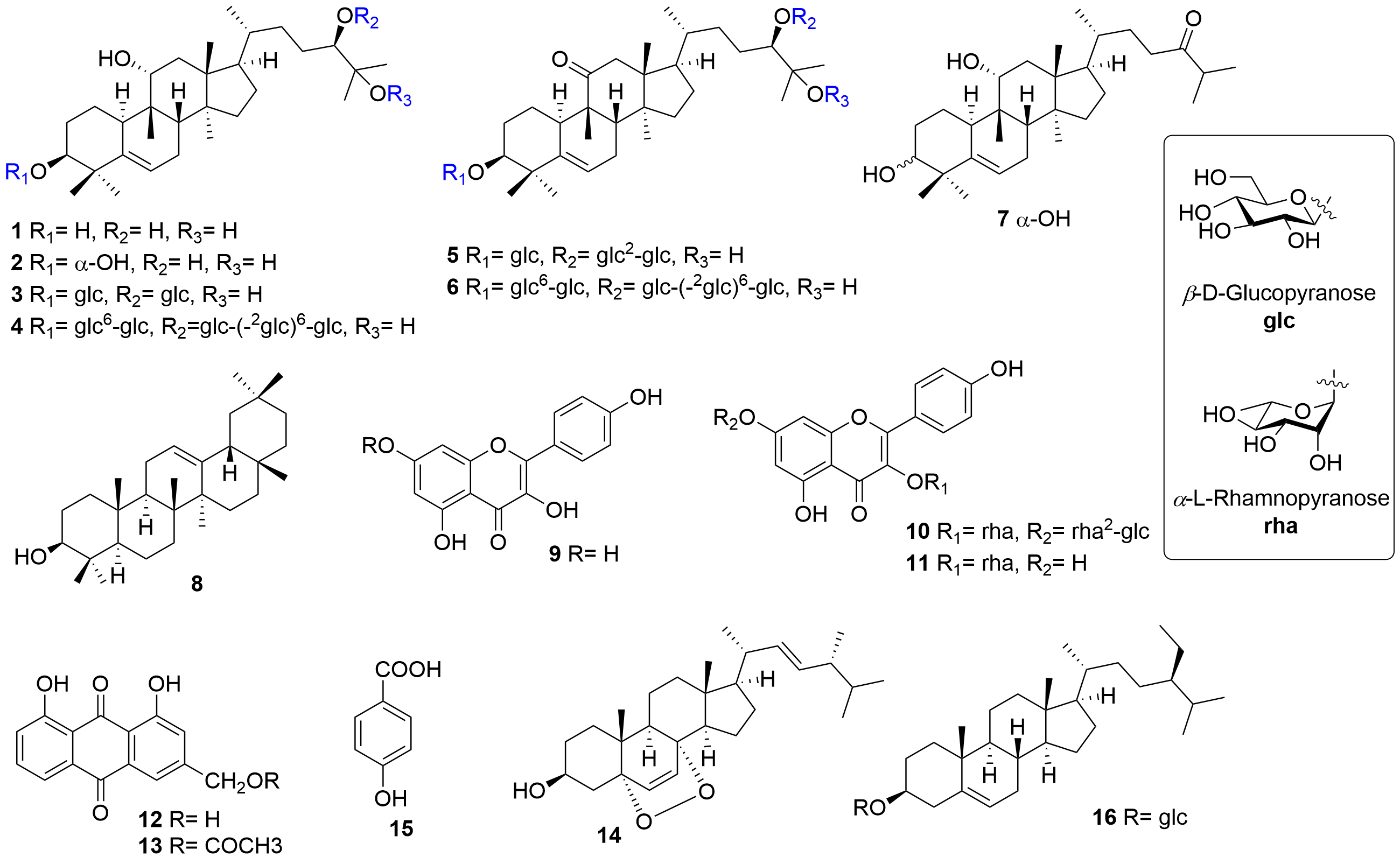
Siraitia grosvenorii (Swingle) C. Jeffrey ex Lu et Z. Y. Zhang is a unique economic and medicinal plant of Cucurbitaceae in Southern China. For hundreds of years, Chinese people have used the fruit of S. grosvenorii as an excellent natural sweetener and traditional medicine for lung congestion, sore throat, and constipation. It is one of the first species in China to be classified as a medicinal food homology, which has received considerable attention as a natural product with high development potential. Various natural products, such as triterpenoids, flavonoids, amino acids, and lignans, have been released from this plant by previous phytochemical studies. Pharmacological research of the fruits of S. grosvenorii has attracted extensive attention, and an increasing number of extracts and compounds have been demonstrated to have antitussive, expectorant, antiasthmatic, antioxidant, hypoglycemic, immunologic, hepatoprotective, antibacterial, and other activities.
1. Antitussive, Expectorant and Anti-Asthmatic Activities
2. Antioxidant
3. Antidiabetic and Antihyperlipidemic Activies
4. Anti-Inflammatory
5. Liver Protection
6. Antibacterial and Antiviral Activities
7. Miscellaneous Activities
 ) has comprehensive anti-cancer activities. The natural sweet compound mogroside V (4) could inhibit the proliferation and survival of pancreatic cancer cells by focusing on multiple biological targets. In both in vitro and in vivo models of pancreatic cancer, mogroside V (4) has tumor growth inhibitory activity by promoting apoptosis and cell cycle arrest in pancreatic cancer cells (PANC-1 cells), possibly through modulation of the STAT3 signalling pathway, promoting cell proliferation (CCND1, CCNE1 and CDK2), while also upregulating cell cycle inhibitors (CDKN1A and CDKN1B) [35]. Mogroside V (4) inhibited hypoglycemic-induced lung cancer cell migration and invasion by reversing EMT and disrupting the cytoskeleton. In lung cancer cells cultured under hypoglycemic conditions, the metastatic efficiency of mogroside V (4) was compared with normoglycemia, which reversed hyperglycemia-induced invasion and migration by upregulating E-Cadherin expression and downregulating N-Cadherin, Vimentin, and Snail expression. Meanwhile, the expression levels of Rho A, Rac1, Cdc42 and p-PAK1 protein were decreased in a dose-dependent manner [36]. After injection of Aβ1-42, mice showed a distinct increase in escape latency and a distinct decrease in the number of times they crossed the target and time spent in the target quadrant. Mogrol (1) could significantly alleviate memory impairment caused by Aβ1-42, inhibit microglia overactivation and prevent hippocampus apoptosis, downregulate the high expression of IL1β, IL-6, NF-kB p65, TNF-a induced by Aβ1–42, reduce the neuroinflammation [37]. Likewise, this plant plays a role in anti-aging. Rats fed with S. grosvenorii showed a slower ageing process. The experimental group maintained a stationary LSK state, decreased ROS level, augmented hematopoietic stem cells, and reduced the number of β-gal positive cells associated with aging, thus lowering the expression of aging-related proteins and slowing the aging process in rats [38].
) has comprehensive anti-cancer activities. The natural sweet compound mogroside V (4) could inhibit the proliferation and survival of pancreatic cancer cells by focusing on multiple biological targets. In both in vitro and in vivo models of pancreatic cancer, mogroside V (4) has tumor growth inhibitory activity by promoting apoptosis and cell cycle arrest in pancreatic cancer cells (PANC-1 cells), possibly through modulation of the STAT3 signalling pathway, promoting cell proliferation (CCND1, CCNE1 and CDK2), while also upregulating cell cycle inhibitors (CDKN1A and CDKN1B) [35]. Mogroside V (4) inhibited hypoglycemic-induced lung cancer cell migration and invasion by reversing EMT and disrupting the cytoskeleton. In lung cancer cells cultured under hypoglycemic conditions, the metastatic efficiency of mogroside V (4) was compared with normoglycemia, which reversed hyperglycemia-induced invasion and migration by upregulating E-Cadherin expression and downregulating N-Cadherin, Vimentin, and Snail expression. Meanwhile, the expression levels of Rho A, Rac1, Cdc42 and p-PAK1 protein were decreased in a dose-dependent manner [36]. After injection of Aβ1-42, mice showed a distinct increase in escape latency and a distinct decrease in the number of times they crossed the target and time spent in the target quadrant. Mogrol (1) could significantly alleviate memory impairment caused by Aβ1-42, inhibit microglia overactivation and prevent hippocampus apoptosis, downregulate the high expression of IL1β, IL-6, NF-kB p65, TNF-a induced by Aβ1–42, reduce the neuroinflammation [37]. Likewise, this plant plays a role in anti-aging. Rats fed with S. grosvenorii showed a slower ageing process. The experimental group maintained a stationary LSK state, decreased ROS level, augmented hematopoietic stem cells, and reduced the number of β-gal positive cells associated with aging, thus lowering the expression of aging-related proteins and slowing the aging process in rats [38].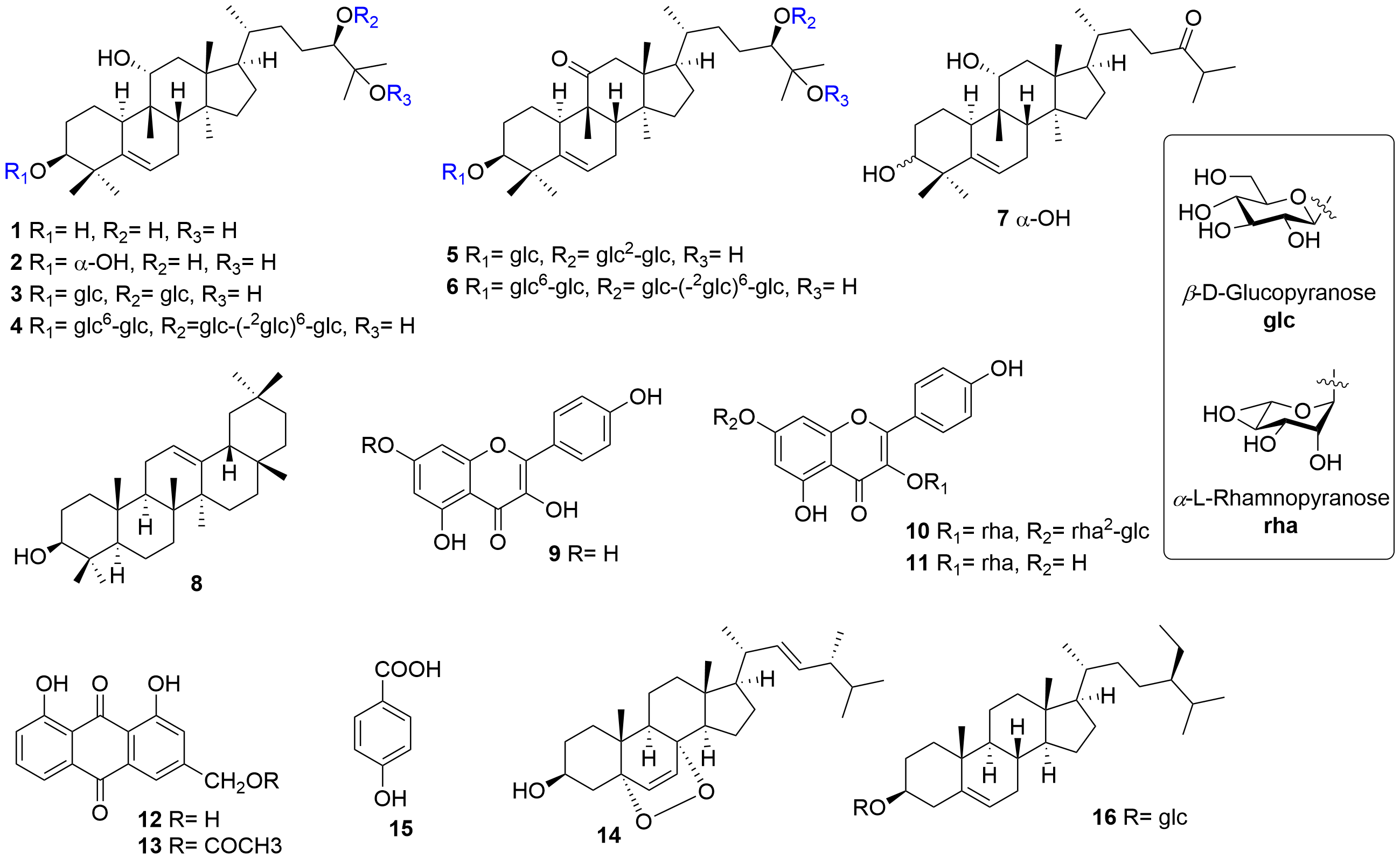
8. Summary of Pharmacologic Activities
| Activities | Detail | Extracts/Compounds | Concentration/Dose | In Vivo/In Vitro | Ref. |
|---|---|---|---|---|---|
| Antitussive, expectorant and anti-asthmatic activities | increased the secretion of phenolsulfonphthalein in mice and the excretion of sputum in rats | SGA | 4000 and 8000 mg/kg | in vivo | [1] |
| reduced Th2 cytokines (IL-4, IL-5, and IL-13) and increased the Th1cytokine IFN-γ | SGRE | 200 mg/kg | in vivo | [2] | |
| enhanced sputum secretion in mice and ciliary cell motility in the frog respiratory tract | mogrosides | 50, 100, and 200 mg/kg | in vivo | [3] | |
| Antioxidant | decrease ROS in oxide injury PC12 cells and decrease apoptotic and necrotic cells | SGP | 0.5, 1.0, 1.5, 2.0 mg/mL | in vitro | [4] |
| reduced ROS levels and MDA content, increase SOD, GSH-Px and CAT activities | mogroside V | 30, 60, and 90 µg/mL | in vitro | [6] | |
| scavenging of -OH | mogroside V | PC12 cells EC50 = 48.44 μg/mL |
in vitro | [5] | |
| scavenging of O2 and H2O2, inhibited -OH induced DNA damage | 11-oxomogroside V | PC12 cells EC50 = 4.79 μg/mL, EC50 = 16.52 μg/mL, EC50 = 3.09 μg/mL | in vitro | [5] | |
| inhibited BSA glycation | MGE | 500 μg/mL | in vitro | [7] | |
| decrease the levels of inflammatory cytokines and oxidative stress-related biomarkers | mogroside IIIE | MPC-5 cells 1, 10, and 50 μM | in vitro | [8] | |
| inhibited the reduction of HB and LD and inhibited the increase of MDA, promoted the synthesis of HB and the clearance of LD | SGE | 1500 mg/kg | in vivo | [9] | |
| Hypoglycemic | increase AMPK phosphorylation | mogrol | HepG2 cell line 1, 10, and 20 μM | in vivo | [11] |
| increase AMPK phosphorylation | 3-hydroxymogrol | HepG2 cell line 1, 10, and 20 μM |
in vivo | [11] | |
| increase AMPK phosphorylation | 3-hydroxy-25-dehydroxy-24-oxo-mogrol | HepG2 cell line 4 μM |
in vivo | [11] | |
| downregulated mRNA levels of hepatic gluconeogenic and lipogenic genes, upregulated fat oxidation-associated genes | MGE | 300 mg/kg | in vivo | [12] | |
| Immunology and anti-inflammatory | increased filaggrin expression and reduced DfE induced phosphorylation of ERK, JNK and p38 | NHGR | 200 or 400 mg/kg | in vivo | [13] |
| enhanced monocyte phagocytosis in hydrocortisone injured | SGA | 2500 and 5000 mg/kg | in vivo | [14] | |
| decreased expression of IL-1b, IL-6, and TNF-a |
mogroside IIIE | GDM model 20.0 mg/kg | in vivo | [15] | |
| inhibited LPS-induced inflammatory |
mogroside IIIE | RAW264.7 cells 10 μM |
in vitro | [15] | |
| inhibited LPS-induced inflammatory |
mogroside IIIE | RAW264.7 cells 1, 10 or 50 μM |
in vitro | [17] | |
| reduced the OVA-induced activation of NF-κB |
mogroside V | 2, 5, and 10 mg/kg | in vivo | [21] | |
| inhibited LPS-induced inflammatory |
mogroside V | BV-2 cells 6.25, 12.5 and 25 μM |
in vitro | [23] | |
| inhibited the IL-9/IL-9R/calcium overload/cathepsin B activation/trypsinogen activation pathway | mogroside IIE | AR42J cells 5, 10 and 20 μM |
in vitro | [24] | |
| Liver protection | inhibited reactive oxygen species production and upregulated sequestosome-1 (SQSTM1, p62) expression |
mogroside V | 200, 400, and 800 mg/kg | in vivo | [25] |
| activated AMPK ameliorates HFD-induced hepatic steatosis | mogroside V | LO2 cells 15, 30, 60, and 120 μM |
in vitro | [26] | |
| Antibacterial and anti-viral | inhibitory effects against gram-positive bacteria | grosvenorine | MIC less than 70 μg/mL | in vitro | [27] |
| inhibitory effects against gram-positive bacteria | kaempferitrin | MIC less than 70 μg/mL | in vitro | [27] | |
| inhibitory effects against gram-positive bacteria and MSSA | kaempferol | MIC less than 70 μg/mL | in vitro | [27] | |
| inhibitory effects against MSSA and MRSA | afzelin | MIC less than 70 μg/mL | in vitro | [27] | |
| inhibitory effects against Streptococcus mutans, Actinobacillus actinobacillus, Clostridium sclerotiorum, and Candida albicans | β-amyrin | MIC = 48.80, >100, 48.40, and >100 μg/mL | in vitro | [29] | |
| inhibitory effects against Streptococcus mutans, Actinobacillus actinobacillus, Clostridium sclerotiorum, and Candida albicans | ergosterol peroxide | MIC = 4.88, 48.80, 48.80, and 12.20 μg/mL | in vitro | [29] | |
| inhibitory effects against Streptococcus mutans, Actinobacillus actinobacillus, Clostridium sclerotiorum, and Candida albicans | aloe-emodin | MIC = 1.22, 6.10, 12.20, and 6.10 μg/mL | in vitro | [29] | |
| inhibitory effects against Streptococcus mutans, Actinobacillus actinobacillus, Clostridium sclerotiorum, and Candida albicans | aloe-emodin acetate | MIC = 6.10, 12.20, >100, and 6.10 μg/mL | in vitro | [29] | |
| inhibitory effects against Streptococcus mutans, Actinobacillus actinobacillus, Clostridium sclerotiorum, and Candida albicans | 4-hydroxybenzoic acid | MIC = 12.20, >100, 12.20, and 12.20 μg/mL | in vitro | [29] | |
| inhibitory effects against Escherichia coli bacterial biofilms | different ethanol-eluting parts of S. grosvenorii | MIC = 55.58, 78.32, and 87.62% | in vitro | [28] | |
| regulation of VEGF | mogroside V | - | in vitro | [33] | |
| Miscellaneous activities | inhibited expression of the STAT3 pathway | mogroside V | PANC-1 cells 10, 100, and 250 μM |
in vivo and in vitro | [35] |
| inhibited expression levels of Rho A, Rac1, Cdc42 and p-PAK1 |
mogroside V | A549 and H1299 cells 0–50 μM |
in vitro | [36] | |
| inhibited expression of IL1β, IL-6, NF-κB p65, TNF-a induced by Aβ1–42 | mogrol | 20, 40, 80 mg/kg | in vivo | [37] | |
| decrease the intercellular levels of ROS | extract of S. grosvenorii | 200 mg/kg | in vivo | [38] |
References
- Zhou, X.X.; Song, J.S. Study on the pharmacological function of fruit and extracts from momordica grosvenorii. Chin. Ach. Tradit. Chin. Med. 2004, 22, 1723–1724.
- Sung, Y.Y.; Kim, S.H.; Yuk, H.J.; Yang, W.K.; Lee, Y.M.; Son, E.J.; Kim, D.S. Siraitia grosvenorii residual extract attenuates ovalbumin-induced lung inflammation by down-regulating IL-4, IL-5, IL-13, IL-17, and MUC5AC expression in mice. Phytomedicine 2019, 61, 152835.
- Wang, T.; Huang, Z.H.; Jiang, Y.M.; Zhou, S.; Su, L.; Jiang, S.Y.; Liu, G.Q. Studies on the pharmaeological profile of mogroside. Chin. Tradit. Herb. Drugs 1999, 30, 914–916.
- Zhu, Y.M.; Pan, L.C.; Zhang, L.J.; Yin, Y.; Zhu, Z.Y.; Sun, H.Q.; Liu, C.Y. Chemical structure and antioxidant activity of a polysaccharide from Siraitia grosvenorii. Int. J. Biol. Macromol. 2020, 165, 1900–1910.
- Chen, W.J.; Wang, J.; Qi, X.Y.; Xie, B.J. The antioxidant activities of natural sweeteners, mogrosides, from fruits of Siraitia grosvenorii. Int. J. Food Sci. Nutr. 2007, 58, 548–556.
- Mo, Q.T.; Fu, H.; Zhao, D.; Zhang, J.C.; Wang, C.; Wang, D.; Li, M. Protective effects of mogroside v on oxidative stress induced by H2O2 in skin fibroblasts. Drug Des., Dev. Ther. 2021, 15, 4901–4909.
- Liu, H.S.; Wang, C.C.; Qi, X.Y.; Zou, J.; Sun, Z.D. Antiglycation and antioxidant activities of mogroside extract from Siraitia grosvenorii (swingle) fruits. J. Food Sci. Technol. 2018, 55, 1880–1888.
- Xue, W.; Mao, J.H.; Chen, Q.J.; Ling, W.D.; Sun, Y.Q. Mogroside IIIE alleviates high glucose-induced inflammation, oxidative stress and apoptosis of podocytes by the activation of AMPK/sirt1 signaling pathway. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 3821–3830.
- Yao, J.W.; Yang, Y.L.; Tang, H.; Zhou, L.; Ding, X.S. Impact of momordica extracts on spon perform ance and free radical metabolism in heart muscle of mice in trainin. J. Beijing Sport Univ. 2009, 67–69.
- Boy, H.; Rutilla, A.; Santos, K.A.; Ty, A.; Yu, A.I.; Mahboob, T.; Tangpoong, J.; Nissapatorn, V. Recommended medicinal plants as source of natural products: A review. Digital Chin. Med. 2018, 1, 131–142.
- Chen, X.B.; Zhuang, J.J.; Liu, J.H.; Lei, M.; Ma, L.; Chen, J.; Shen, X.; Hu, L.H. Potential ampk activators of cucurbitane triterpenoids from Siraitia grosvenorii swingle. Bioorg. Med. Chem. 2011, 19, 5776–5781.
- Liu, H.S.; Qi, X.Y.; Yu, K.K.; Lu, A.J.; Lin, K.F.; Zhu, J.J.; Zhang, M.; Sun, Z.D. AMPK activation is involved in hypoglycemic and hypolipidemic activities of mogroside-rich extract from Siraitia grosvenorii (swingle) fruits on high-fat diet/streptozotocin-induced diabetic mice. Food Funct. 2019, 10, 151–162.
- Sung, Y.Y.; Yuk, H.J.; Yang, W.K.; Kim, S.H.; Kim, D.S. Siraitia grosvenorii residual extract attenuates atopic dermatitis by regulating immune dysfunction and skin barrier abnormality. Nutrients 2020, 12, 3638.
- Wang, Q.; Y, L.A.; Li, X.P. Pharmacological effects of S. grosvenorii fruit. China J. Chin. Mater Med. 1999, 24, 425–428.
- Zou, C.L.; Zhang, Q.Q.; Zhang, S.H. Mogroside IIIE attenuates gestational diabetes mellitus through activating of ampk signaling pathway in mice. J. Pharmacol. Sci 2018, 138, 161–166.
- Tao, L.J.; Yang, J.Y.; Cao, F.Y.; Xie, H.F.; Zhang, M.; Gong, Y.Q.; Zhang, C.F. Mogroside IIIE, a novel anti-fibrotic compound, reduces pulmonary fibrosis through toll-like receptor 4 pathways. J. Pharmacol. Exp. Ther. 2017, 361, 268–279.
- Tao, L.J.; Cao, F.Y.; Xu, G.H.; Xie, H.F.; Zhan, M.; Zhang, C.F. Mogroside IIIE attenuates LPS-induced acute lung injury in mice partly through regulation of the tlr4/MAPK/NF-κB axis via ampk activation. Phytother. Res. 2017, 31, 1097–1106.
- Han, B.Y.; Michael, R.; Epperly, W.; Cao, S.N.; Goff, J. Improved longevity of hematopoiesis in long-term bone marrow cultures and reduced irradiation-induced pulmonary fibrosis in toll-like receptor-4 deletion recombinant-negative mice. Vivo 2015, 28, 441–448.
- Bhattacharyya, S.; Kelley, K.; Melichian, D.S.; Tamaki, Z.; Fang, F.; Su, Y.Y.; Feng, G.; Pope, R.M.; Budinger, G.R.S.; Mutlu, G.M. Toll-like receptor 4 signaling augments transforming growth factor-β responses: A novel mechanism for maintaining and amplifying fibrosis in scleroderma. Am. J. Pathol. 2013, 182, 192–205.
- Bueno, G.B.; Caso, J.R.; Madrigal, J.; Leza, J.C. Innate immune receptor toll-like receptor 4 signalling in neuropsychiatric diseases. Neurosci. Biobehav. Rev. 2016, 134–147.
- Song, J.L.; Qian, B.; Pan, C.L.; Lv, F.F.; Wang, H.P.; Gao, Y.; Zhou, Y.Y. Protective activity of mogroside v against ovalbumin-induced experimental allergic asthma in kunming mice. J. Food Biochem. 2019, 43, e12973.
- Carty, M.; Bowie, A.G. Evaluating the role of toll-like receptors in diseases of the central nervous system. Biochem. Pharmacol. 2011, 81, 825–837.
- Liu, Y.Y.; Zhang, B.X.; Liu, J.H.; Qiao, C.Y.; Xue, N.Y.; Lv, H.M.; Li, S.Z. Mogroside V alleviates lipopolysaccharide-induced neuroinflammation via inhibition of tlr4-Myd88 and activation of akt/ampk-nrf2 signaling pathway. J. Evid.-Based Complementary Altern. Med. 2021, 2021, 5521519.
- Xiao, J.; Huang, K.; Lin, H.M.; Xia, Z.J.; Zhang, J.; Li, D.; Jin, J.F. Mogroside IIE inhibits digestive enzymes via suppression of interleukin 9/interleukin 9 receptor signalling in acute pancreatitis. Front. Pharmacol. 2020, 11, 859.
- Zhang, X.B.; Song, Y.F.; Ding, Y.P.; Wang, W.; Liao, L.; Zhong, J.; Sun, P.B.; Lei, F.; Zhang, Y.O.; Xie, W.D. Effects of mogrosides on high-fat-diet-induced obesity and nonalcoholic fatty liver disease in mice. Molecules 2018, 23, 1894.
- Li, L.H.; Zheng, W.F.; Wang, C.; Qi, J.M.; Li, H.B. Mogroside V protects against hepatic steatosis in mice on a high-fat diet and lo2 cells treated with free fatty acids via AMPK activation. Evid. Based Complementary Altern. Med. 2020, 2020, 7826874.
- Wang, M.Y.; Xing, S.H.; Luu, T.; Fan, M.; Li, X.B. The gastrointestinal tract metabolism and pharmacological activities of grosvenorine, a major and characteristic flavonoid in the fruits of Siraitia grosvenorii. Chem. Biodiversity 2016, 12, 1652–1664.
- Aguero, M.B.; Gonzalez, M.; Lima, B.; Svetaz, L.; Sanchez, M.; Zacchino, S.; Feresin, G.E.; Schmeda-Hirschmann, G.; Palermo, J.; Wunderlin, D. Argentinean propolis from zuccagnia punctata cav. (Caesalpinieae) exudates: Phytochemical characterization and antifungal activity. J. Agric. Food Chem. 2010, 58, 194–201.
- Zheng, Y.; Wen, H.; Yoo, J.G.; Ebersole, J.L.; Huang, C.B. Antibacterial compounds from Siraitia grosvenorii leaves. Nat. Prod. Res 2011, 25, 890–897.
- Wang, H.Y.; Wang, T.; Li, H.Y.; Xu, D.D. Selecting active constituents of siraitiae fructus on inhibition of escherichia coli bacterial biofilms. Chin. J. Exp. Tradit. Med. Formulae 2016, 22, 51–54.
- Aslan, A.; Aslan, C.; Zolbanin, N.M.; Jafari, R. Acute respiratory distress syndrome in COVID-19: Possible mechanisms and therapeutic management. Pneumonia (Nathan) 2021, 13, 14.
- Yarza, R.; Bover, M.; Paredes, D.; López-López, F.; Jara-Casas, D.; Castelo-Loureiro, A.; Baena, J.; Mazarico, J.M.; Folgueira, M.D.; Meléndez-Carmona, M.Á.; et al. SARS-COV-2 infection in cancer patients undergoing active treatment: Analysis of clinical features and predictive factors for severe respiratory failure and death. Eur. J. Cancer 2020, 135, 242–250.
- Li, Y.; Chen, Y.; Wei, M.; Wei, C. Preclinical in silico evidence indicates the pharmacological targets and mechanisms of mogroside V in patients with ovarian cancer and coronavirus disease 2019. Front. Endocrinol. (Lausanne) 2022, 13, 845404.
- Bortolotti, D.; Simioni, C.; Neri, L.M.; Rizzo, R.; Semprini, C.M.; Occhionorelli, S.; Laface, I.; Sanz, J.M.; Schiuma, G.; Rizzo, S.; et al. Relevance of vegf and cd147 in different SARS-COV-2 positive digestive tracts characterized by thrombotic damage. FASEB J. 2021, 35, e21969.
- Liu, C.; Dai, L.H.; Dou, D.Q.; Ma, L.Q.; Sun, Y.X. A natural food sweetener with anti-pancreatic cancer properties. Oncogenesis 2016, 5, e217.
- Chen, J.; Jiao, D.M.; Li, Y.; Jiang, C.Y.; Chen, Q.Y. Mogroside v inhibits hyperglycemia-induced lung cancer cells metastasis through reversing emt and damaging cytoskeleton. Curr. Cancer Drug Targets 2019, 19, 885–895.
- Chen, G.L.; Liu, C.H.; Meng, G.L.; Zhang, C.T.; Chen, F.; Tang, S.S.; Hong, H.; Zhang, C.F. Neuroprotective effect of mogrol against aβ1-42-induced memory impairment neuroinflammation and apoptosis in mice. J. Pharm. Pharmacol. 2019, 71, 869–877.
- Bai, L.; Shi, G.Y.; Yang, Y.J.; Chen, W.; Zhang, L.F. Anti-aging effect of siraitia grosuenorii by enhancement of hematopoietic stem cell function. Am. J. Chin. Med. 2016, 803–815.



