 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Chetan Keswani | -- | 3012 | 2022-10-19 12:33:31 | | | |
| 2 | Catherine Yang | -1 word(s) | 3011 | 2022-10-20 03:24:19 | | |
Video Upload Options
Abiotic stress factors are considered a serious threat to various growth parameters of crop plants. Stressors such as drought, salinity, and heavy metals (HMs) hamper the chlorophyll content in plants, resulting in low photosynthesis, hinder the integrity of cell membranes, reduce biomass, and overall growth and development of crops which ultimately results in the sharp decline of crop yield. Under such stressful conditions, various strategies are employed to overcome hazardous effects. Application of Zinc (Zn) or Selenium (Se) in different forms is an effective way to alleviate the abiotic stresses in plants. Zn and Se play a pivotal role in enhancing the chlorophyll level to improve photosynthesis, reducing oxidative stress by limiting reactive oxygen species (ROS) production, controlling HMs absorption by plant roots and their accumulation in the plant body, maintaining homeostasis, and alleviating all the detrimental effects caused by abiotic stress factors.
1. Zinc (Zn) and Selenium (Se) Absorption and Transport in Plants

2. Importance of Zinc and Selenium and the Effects of Their Deficiency on Plants


3. Process of Zn and Se Application, Its Downstream Effects on Plants
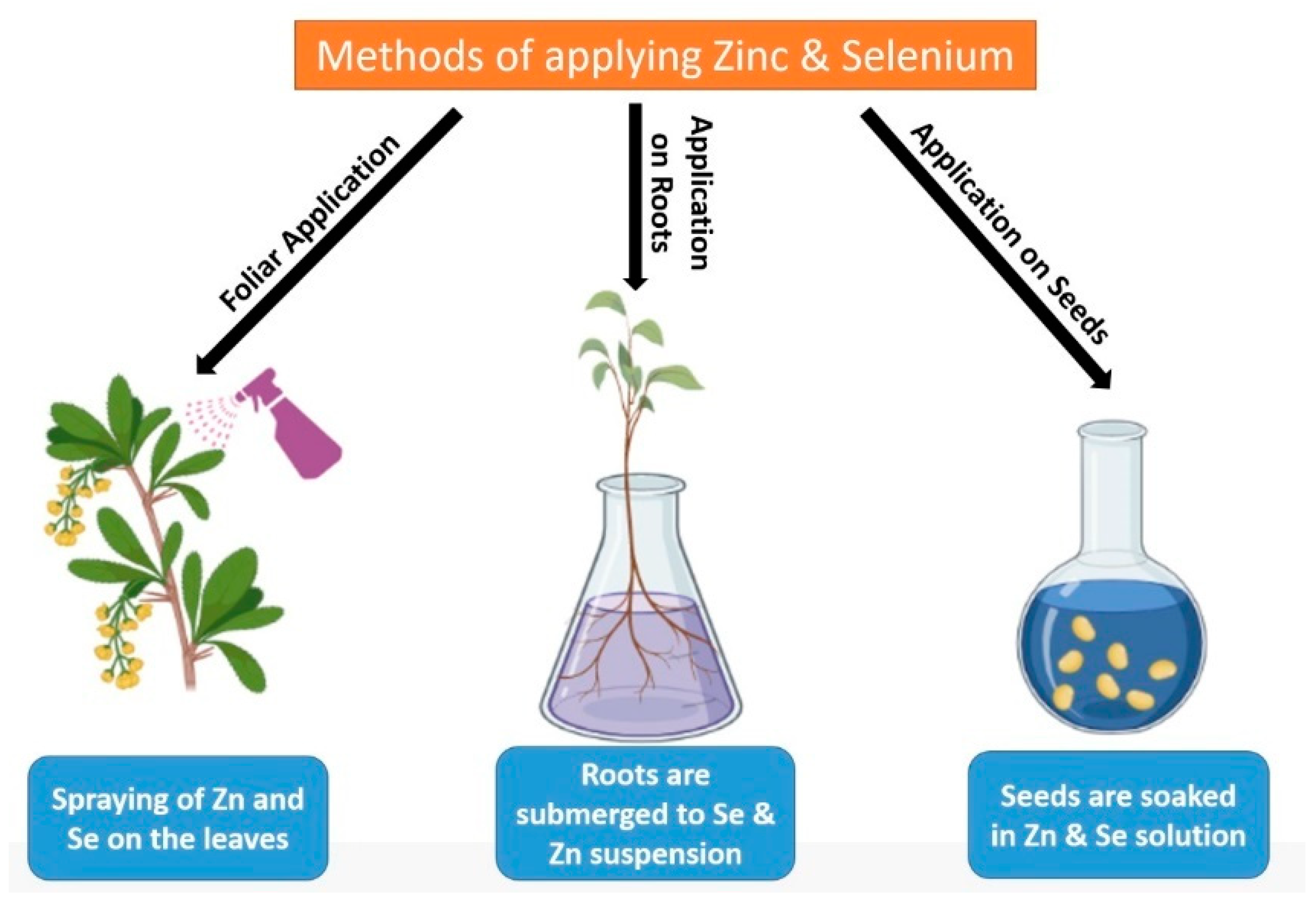
| Name of Plant. | The Part on Which Zn Is Applied | Utilities as a Result of Zn Application | References |
|---|---|---|---|
| Maize | Foliar application | Increases starch content Improves crop yield |
[35] |
| Mungbean | Foliar application | Increases the growth and productivity of the crop | [38] |
| Maize and Wheat | Foliar application | Enhances the yield of both the grains | [34] |
| French Bean | Foliar application | Improves in the physiological traits Enhances crop quality Improves productivity of grains |
[39] |
| Maize | Soil | Advances crop yield | [41] |
| Wheat | Soil | Increases in grain productivity | [42] |
| Wheat | Foliar + soil | remarkable boost in the yield | [9] |
| Bread wheat | Foliar + soil | Improves the growth rate Enhances crop productivity Increases Zn content in wheat |
[10] |
| Garden pansy | Foliar application of Se in the form of sodium selenate | Increases fresh weight by 25.10% Increases dry weight by 25.41% |
[41] |
| Rice | Foliar application of Se in the form of selenate and selenite | Enhances the productivity of rice grains | [37] |
| Rice | Soil application of Se | Produces more shoot dry matter | [37] |
| Oat | Foliar + soil application of Se in the form of Se fertilizer | Improves Se transport Enhances crop yield |
[42] |
| Atractylodes macrocephala | Foliar application of Se | Increases the growth and survival rate Enhances crop yield |
[43] |
4. Role of Zn and Se in Plants under Salinity Stress
References
- Vadlamudi, K.; Upadhyay, H.; Singh, A.; Reddy, M. Influence of Zinc Application in Plant Growth: An Overview. Eur. J. Mol. Clin. Med. 2020, 7, 2321–2327.
- Gupta, N.; Ram, H.; Kumar, B. Mechanism of Zinc Absorption in Plants: Uptake, Transport, Translocation and Accumulation. Rev. Environ. Sci. Biotechnol. 2016, 15, 89–109.
- Silva, V.M.; Rimoldi Tavanti, R.F.; Gratão, P.L.; Alcock, T.D.; dos Reis, A.R. Selenate and Selenite Affect Photosynthetic Pigments and ROS Scavenging through Distinct Mechanisms in Cowpea (Vigna unguiculata (L.) Walp) Plants. Ecotoxicol. Environ. Saf. 2020, 201, 110777.
- Palmer, C.M.; Guerinot, M.L. Facing the Challenges of Cu, Fe and Zn Homeostasis in Plants. Nat. Chem. Biol. 2009, 5, 333–340.
- Demidchik, V.; Davenport, R.J.; Tester, M. Nonselective Cation Channels in Plants. Annu. Rev. Plant Biol. 2002, 53, 67–107.
- Sondergaard, T.E.; Schulz, A.; Palmgren, M.G. Energization of Transport Processes in Plants. Roles of the Plasma Membrane H+-ATPase. Plant Physiol. 2004, 136, 2475–2482.
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium Speciation from Food Source to Metabolites: A Critical Review. Anal. Bioanal. Chem. 2006, 385, 1304–1323.
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 2074.
- Harris, J.; Schneberg, K.A.; Pilon-Smits, E.A.H. Sulfur-Selenium-Molybdenum Interactions Distinguish Selenium Hyperaccumulator Stanleya Pinnata from Non-Hyperaccumulator Brassica Juncea (Brassicaceae). Planta 2014, 239, 479–491.
- Mazej, D.; Osvald, J.; Stibilj, V. Selenium Species in Leaves of Chicory, Dandelion, Lamb’s Lettuce and Parsley. Food Chem. 2008, 107, 75–83.
- Chattha, M.U.; Hassan, M.U.; Khan, I.; Chattha, M.B.; Mahmood, A.; Chattha, M.U.; Nawaz, M.; Subhani, M.N.; Kharal, M.; Khan, S. Biofortification of Wheat Cultivars to Combat Zinc Deficiency. Front. Plant Sci. 2017, 8, 281.
- Hassan, M.U.; Chattha, M.U.; Ullah, A.; Khan, I.; Qadeer, A.; Aamer, M.; Khan, A.U.; Nadeem, F.; Khan, T.A. Agronomic Biofortification to Improve Productivity and Grain Zn Concentration of Bread Wheat. Int. J. Agric. Biol. 2019, 21, 615–620.
- Hasanuzzaman, M.; Nahar, K.; García-Caparrós, P.; Parvin, K.; Zulfiqar, F.; Ahmed, N.; Fujita, M. Selenium Supplementation and Crop Plant Tolerance to Metal/Metalloid Toxicity. Front. Plant Sci. 2022, 12, 792770.
- Mousavi, S.R.; Galavi, M.; Ahmadvand, G. Effect of Zinc and Manganese Foliar Application on Yield, Quality and Enrichment on Potato (Solanum tuberosum L.). Asian J. Plant Sci. 2007, 6, 1256–1260.
- Zafar, S.; Ashraf, M.Y.; Saleem, M. Shift in Physiological and Biochemical Processes in Wheat Supplied with Zinc and Potassium under Saline Condition. J. Plant Nutr. 2018, 41, 19–28.
- Sharma, P.N.; Kumar, N.; Bisht, S.S. Effect of Zinc Deficiency on Chlorophyll Content, Photosynthesis and Water Relations of Cauliflower Plants. Photosynth. Prague 1994, 30, 353–359.
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H. Selenium Promotes the Growth and Photosynthesis of Tomato Seedlings under Salt Stress by Enhancing Chloroplast Antioxidant Defense System. J. Plant Growth Regul. 2014, 33, 671–682.
- Al-Zahrani, H.S.; Alharby, H.F.; Hakeem, K.R.; Rehman, R.U. Exogenous Application of Zinc to Mitigate the Salt Stress in Vigna radiata (L.) Wilczek—Evaluation of Physiological and Biochemical Processes. Plants 2021, 10, 1005.
- Hussein, M.M.; Abou-Baker, N.H. The Contribution of Nano-Zinc to Alleviate Salinity Stress on Cotton Plants. R. Soc. Open Sci. 2018, 5, 171809.
- Hassan, M.; Aamer, M.; Chattha, M.; Tang, H.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The Critical Role of Zinc in Plants Facing the Drought Stress. Agriculture 2020, 10, 396.
- ul Hassan, Z.; Ali, S.; Rizwan, M.; Hussain, A.; Akbar, Z.; Rasool, N.; Abbas, F. Role of Zinc in Alleviating Heavy Metal Stress. In Essential Plant Nutrients: Uptake, Use Efficiency, and Management; Naeem, M., Ansari, A.A., Gill, S.S., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 351–366.
- Morkunas, I.; Woźniak, A.; Mai, V.C.; Rucińska-Sobkowiak, R.; Jeandet, P. The Role of Heavy Metals in Plant Response to Biotic Stress. Mol. J. Synth. Chem. Nat. Prod. Chem. 2018, 23, 2320.
- Hafeez, B.; Khanif, Y.M.; Saleem, M. Role of Zinc in Plant Nutrition-A Review. Am. J. Exp. Agric. 2013, 3, 374.
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers; Macmillan Publishing Company: New York, NY, USA, 1985.
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press, Elsevier Ltd.: Cambridge, MA, USA, 1995.
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2000.
- Dang, H.; Li, R.; Sun, Y.; Zhang, X.; Li, Y. Absorption, Accumulation and Distribution of Zinc in Highly-Yielding Winter Wheat. Agric. Sci. China 2010, 9, 965–973.
- Disante, K.B.; Fuentes, D.; Cortina, J. Response to Drought of Zn-Stressed Quercus Suber L. Seedlings. Environ. Exp. Bot. 2011, 70, 96–103.
- Lerdau, M. Mineral Nutrition of Plants: Principles and Perspectives. Second Edition. By Emanuel Epstein and Arnold J Bloom. Q. Rev. Biol. 2005, 80, 359.
- Alloway, B.J. Micronutrients and Crop Production: An Introduction. In Micronutrient Deficiencies in Global Crop Production; Alloway, B.J., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–39.
- Welch, R.M. The Impact of Mineral Nutrients in Food Crops on Global Human Health. Plant Soil 2002, 247, 83–90.
- Snowball, K.; Robson, A.D. Symptoms of Nutrient Deficiencies: Lupins; Soil Science and Plant Nutrition, School of Agriculture, University of Western Australia: Nedlands, Australia, 1986.
- Neue, H.U.; Lantin, R.S. Micronutrient Toxicities and Deficiencies in Rice. In Soil Mineral Stresses: Approaches to Crop Improvement; Yeo, A.R., Flowers, T.J., Eds.; Monographs on Theoretical and Applied Genetics; Springer: Berlin/Heidelberg, Germany, 1994; pp. 175–200.
- Van Breemen, N.; Quijano, C.C.; Sen, L.N. Zinc Deficiency in Wetland Rice Along a Toposequence of Hydromorphic Soils in the Philippines: I. Soil Conditions and Hydrology. Plant Soil 1980, 57, 203–214.
- Yoshida, S.; Tanaka, A. Zinc Deficiency of the Rice Plant in Calcareous Soils. Soil Sci. Plant Nutr. 1969, 15, 75–80.
- Fageria, N.K. Dry Matter Yield and Nutrient Uptake by Lowland Rice at Different Growth Stages. J. Plant Nutr. 2004, 27, 947–958.
- Cakmak, I. Tansley Review No. 111: Possible Roles of Zinc in Protecting Plant Cells from Damage by Reactive Oxygen Species. New Phytol. 2000, 146, 185–205.
- Grzebisz, W.; Wronska, M.; Diatta, J.B.; Szczepaniak, W. Effect of Zinc Foliar Application at an Early Stage of Maize Growth on Patterns of Nutrients and Dry Matter Accumulation by the Canopy. Part II. Nitrogen Uptake and Dry Matter Accumulation Patterns. J. Elem. 2008, 13, 29–40.
- Leach, K.A.; Hameleers, A. The Effects of a Foliar Spray Containing Phosphorus and Zinc on the Development, Composition and Yield of Forage Maize. Grass Forage Sci. 2001, 56, 311–315.
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Boldrin, K.V.F.; Ávila, F.W.; Guilherme, L.R.G. Soil and Foliar Application of Selenium in Rice Biofortification. J. Food Compos. Anal. 2013, 31, 238–244.
- Javadi, F.; Kalatejari, S.; Diyanat, M. Effect of Foliar or Soil Application of Selenium on Some Morphological and Physiological Traits of Garden Pansy (Viola x Wittrockiana Gams) Grown under Salinity Stress. Acta Agric. Slov. 2020, 115, 357–368.
- Li, J.; Yang, W.; Guo, A.; Yang, S.; Chen, J.; Qiao, Y.; Anwar, S.; Wang, K.; Yang, Z.; Gao, Z.; et al. Combined Foliar and Soil Selenium Fertilizer Improves Selenium Transport and the Diversity of Rhizosphere Bacterial Community in Oats. Environ. Sci. Pollut. Res. Int. 2021, 28, 64407–64418.
- Zhou, W.; Duan, Y.; Zhang, Y.; Wang, H.; Huang, D.; Zhang, M. Effects of Foliar Selenium Application on Growth and Rhizospheric Soil Micro-Ecological Environment of Atractylodes Macrocephala Koidz. S. Afr. J. Bot. 2021, 137, 98–109.
- Thalooth, A.; Tawfik, M.; Mohamed, H. A Comparative Study on the Effect of Foliar Application of Zinc, Potassium and Magnesium on Growth, Yield and Some Chemical Constituents of Mungbean Plants Grown under Water Stress Conditions. World J. Agric. Sci. 2006, 2, 37–46.
- Yadavi, A.; Aboueshaghi, R.; Movahhedi-Dehnavi, M.; Balouchi, H. Effect of micronutrients foliar application on grain qualitative characteristics and some physiological traits of bean (Phaseolus vulgaris L.) Under drought stress. Indian J. Fundam. Appl. Life Sci. 2014, 4, 124–131.
- Cakmak, I.; Pfeiffer, W.H.; McClafferty, B. Review: Biofortification of Durum Wheat with Zinc and Iron. Cereal Chem. 2010, 87, 10–20.
- Ehsanullah; Tariq, A.; Randhawa, M.A.; Anjum, S.A.; Nadeem, M.; Naeem, M. Exploring the Role of Zinc in Maize (Zea mays L.) through Soil and Foliar Application. Univers. J. Agric. Res. 2015, 3, 69–75.
- Khan, H.; Maitlo, A.A. Yield and Micronutrients Content of Bread Wheat (Triticum aestivum L.) under a Multinutrient Fertilizer–Hal-Tonic. Int. J. Agric. Biol. 2006, 8, 5.
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops–What Is the Cost? New Phytol. 2015, 208, 668–673.
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High Concentrations of Na+ and Cl– Ions in Soil Solution Have Simultaneous Detrimental Effects on Growth of Faba Bean under Salinity Stress. J. Exp. Bot. 2010, 61, 4449–4459.
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous Selenium Pretreatment Protects Rapeseed Seedlings from Cadmium-Induced Oxidative Stress by Upregulating Antioxidant Defense and Methylglyoxal Detoxification Systems. Biol. Trace Elem. Res. 2012, 149, 248–261.
- Grattan, S.R.; Grieve, C.M. Mineral Element Acquisition and Growth Response of Plants Grown in Saline Environments. Agric. Ecosyst. Environ. 1992, 38, 275–300.
- Parker, D.R.; Aguilera, J.J.; Thomason, D.N. Zinc-Phosphorus Interactions in Two Cultivars of Tomato (Lycopersicon esculentum L.) Grown in Chelator-Buffered Nutrient Solutions. Plant Soil 1992, 143, 163–177.
- Arough, Y.K.; Sharifi, R.S.; Sedghi, M.; Barmaki, M. Effect of Zinc and Bio Fertilizers on Antioxidant Enzymes Activity, Chlorophyll Content, Soluble Sugars and Proline in Triticale Under Salinity Condition. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 116–124.
- Upadhyaya, H.; Shome, S.; Tewari, S.; Bhattacharya, M.K.; Panda, S. Effect of Zn Nano-Particles on Growth Responses of Rice; Mc Graw Hill PVT.: Delhi, India, 2015.
- Torabian, S.; Zahedi, M.; Khoshgoftarmanesh, A. Effect of Foliar Spray of Zinc Oxide on Some Antioxidant Enzymes Activity of Sunflower under Salt Stress. J. Agric. Sci. Technol. 2016, 18, 1013–1025.
- Babaeian, M.; Piri, I.; Tavassoli, A.; Esmaeilian, Y.; Gholami, H. Effect of Water Stress and Micronutrients (Fe, Zn and Mn) on Chlorophyll Fluorescence, Leaf Chlorophyll Content and Sunflower Nutrient Uptake in Sistan Region. Afr. J. Agric. Res. 2011, 6, 3526–3531.
- Alharby, H.; Hasanuzzaman, M.; Al-Zahrani, H.; Hakeem, K. Exogenous Selenium Mitigates Salt Stress in Soybean by Improving Growth, Physiology, Glutathione Homeostasis and Antioxidant Defense. Phyton-Int. J. Exp. Bot. 2021, 90, 373–388.
- El-Badri, A.M.; Batool, M.; Wang, C.; Hashem, A.M.; Tabl, K.M.; Nishawy, E.; Kuai, J.; Zhou, G.; Wang, B. Selenium and Zinc Oxide Nanoparticles Modulate the Molecular and Morpho-Physiological Processes during Seed Germination of Brassica Napus under Salt Stress. Ecotoxicol. Environ. Saf. 2021, 225, 112695.


