 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dawei Xue | -- | 2617 | 2022-08-30 16:06:15 | | | |
| 2 | Peter Tang | Meta information modification | 2617 | 2022-08-31 03:01:57 | | |
Video Upload Options
Plant lesion mutation usually refers to the phenomenon of cell death in green tissues before senescence in the absence of external stress, and such mutants also show enhanced resistance to some plant pathogens. The occurrence of lesion mimic mutants in rice is affected by gene mutation, reactive oxygen species accumulation, an uncontrolled programmed cell death system, and abiotic stress.
1. Introduction
2. Discovery, Excavation, and Classification of Rice Lesion Mimic Mutants
3. Mechanism of Rice Lesion Occurrence
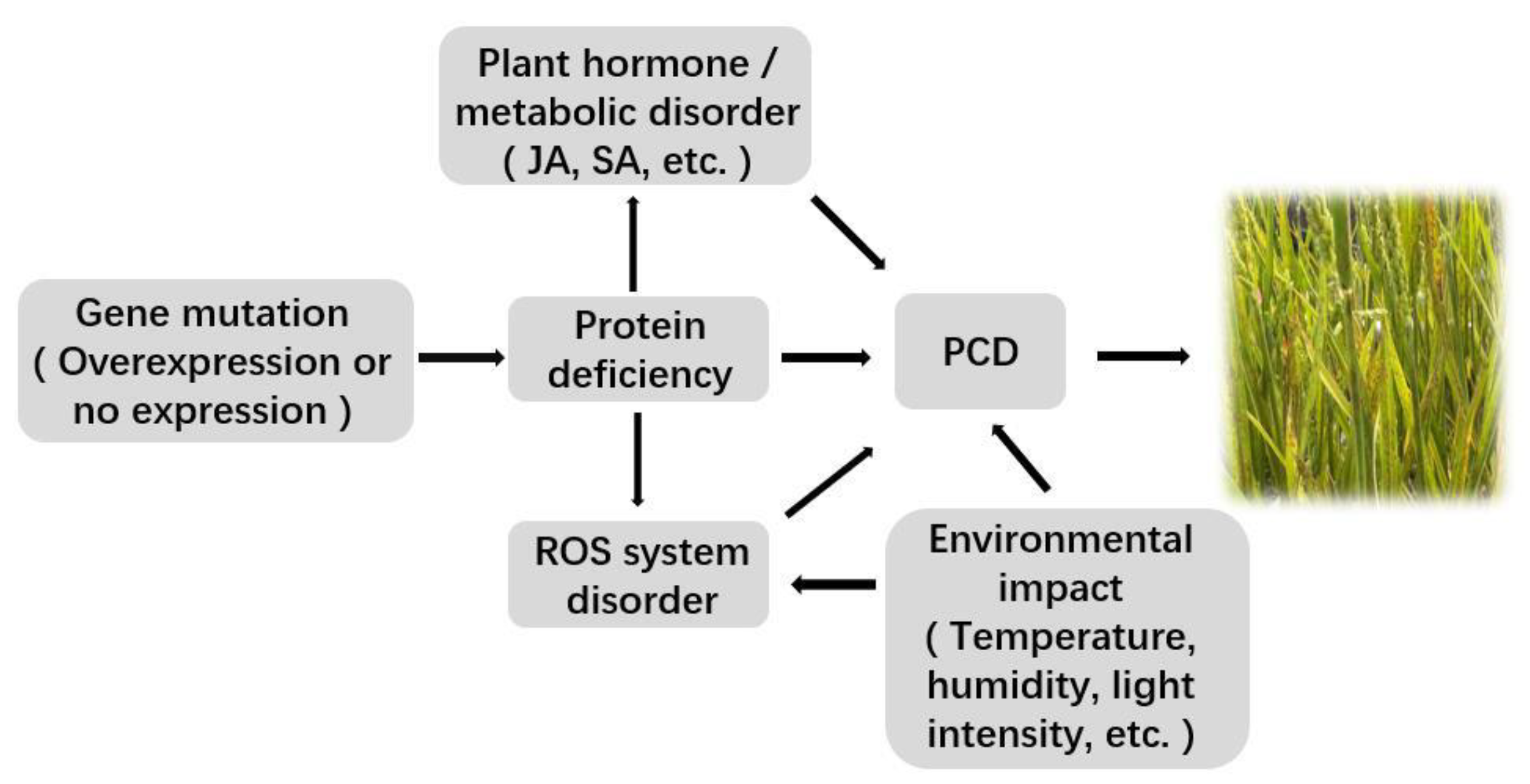
3.1. Related Gene Mutation or Expression Change
3.2. Accumulation of ROS
3.3. Effects of Plant Hormones
3.4. Disorder of Plant Metabolic Pathways
3.5. Uncontrolled PCD
3.6. Influence of the External Environment
4. Identification, Cloning, and Functional Analysis of Rice Lesion Mimic Genes
|
Mutant Gene |
Mutant Type |
Accession Number |
Protein Function Analysis |
Reference |
|---|---|---|---|---|
|
SPL7 |
RILM |
Os05g05304 |
Heat shock transcription factor |
[7] |
|
SPL11 |
RILM |
Os12g05700 |
E3 ubiquitin ligase |
[28] |
|
LSD1 |
VILM |
Os08g01595 |
C2C2 zinc finger protein |
[31] |
|
OsNPR1(NH1) |
VILM |
Os01g01943 |
Transcriptional coactivators |
[34] |
|
SPL18(OsAT1) |
WLLM |
Os10g01956 |
Acyltransferase |
[14] |
|
OsPti1a |
VILM |
Os05g01358 |
Rice protein kinase |
[35] |
|
XB15 |
VILM |
Os03g60650 |
Protein phosphatase |
[36] |
|
OsACDR1 (OsEDR1,SPL3) |
RILM |
Os03g01601 |
Mitogen-activated protein kinase |
[29] |
|
OsSSI2 |
RILM |
Os01g09199 |
Fatty acid dehydrogenase |
[22] |
|
OsPLDβ1 |
VILM |
Os10g38060 |
Phospholipase |
[37] |
|
SPL28 |
RILM |
Os01g07036 |
Subunits of grid-related receptor protein complexes |
[26] |
|
OsSL(ELL1) |
RILM |
Os12g02680 |
Cytochrome P450 monooxygenase |
|
|
RLIN1 |
WLLM |
Os04g06108 |
Porphyrin III oxidase |
[40] |
|
GF14e |
WLLM |
Os02g05803 |
14-3-3 protein |
[30] |
|
NOE1 |
RILM |
Os03g03910 |
Catalase |
[31] |
|
NLS1 |
RILM |
Os11g14380 |
CC-NB-LRR type R protein |
[13] |
|
SPL5 |
RILM |
Os07g02037 |
Splicing factor 3b subunit |
[41] |
|
OsHPL3 |
VILM |
Os02g01102 |
Hydroperoxide lyase |
|
|
OsLMS |
WLLM |
Os02g06390 |
RNA binding protein |
[43] |
|
CslF6 |
VILM |
Os08g01605 |
Cellulose-like synthase F |
[44] |
|
RLS1 |
VILM |
Os02g10900 |
NB-ARM domain protein |
[45] |
|
FGL |
VILM |
Os10g35370 |
OsPORB protein, involved in cytochrome synthesis |
[25] |
|
SPL29 |
WLLM |
Os08g02069 |
Acetylglucosamine pyrophosphatase |
[23] |
|
LMR |
RILM |
Os06g01300 |
Adenosine triphosphatase |
[46] |
|
EBR1 |
VILM |
Os05g19970 |
E3 ubiquitin ligase in RING domain |
[47] |
|
OsPLS1 |
RILM |
Os06g45120 |
Vacuolar proton ATPase subunit |
[48] |
|
SPL32 |
VILM |
Os07g06584 |
Ferroxin-dependent glutamate synthase |
[49] |
|
SPL33 |
WLLM |
Os01g01166 |
eEF1A-like protein |
[50] |
|
OsCUL3a |
VILM |
Os02g07460 |
Cullin protein |
[51] |
|
LML1 |
WLLM |
Os04g06599 |
Eukaryotic Release Factor 1 Protein |
[52] |
|
SDS |
VILM |
Os01g57480 |
SD-1 receptor-like kinase |
[53] |
|
SPL35 |
WLLM |
Os03g02050 |
CUE domain protein |
[54] |
|
SPL40 |
WLLM |
Os05g03120 |
Ribosome structural components |
[55] |
|
LMM24 |
VILM |
Os03g24930 |
receptor-like cytoplasmic kinase |
[56] |
|
OsRLR1 |
RILM |
Os10g07978 |
CC-NB-LRR protein |
[57] |
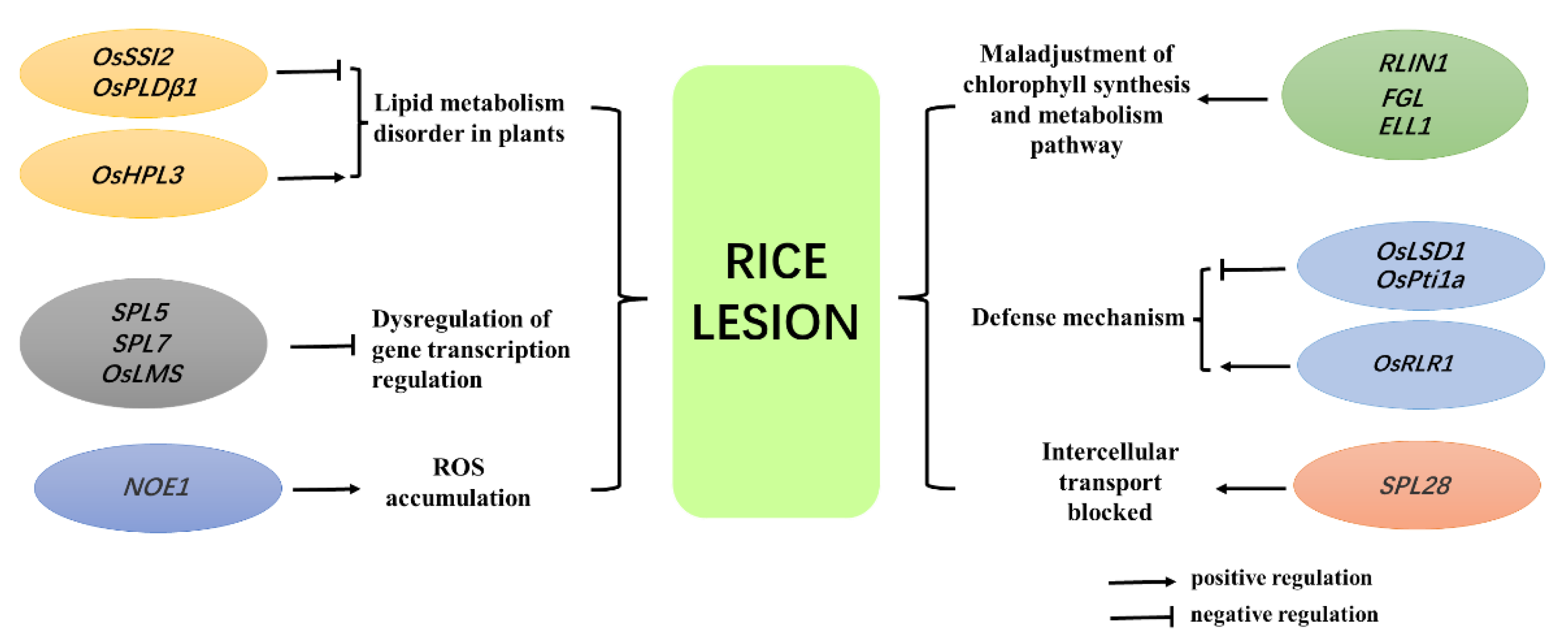
The lesion mimic mutants can be divided into whole life lesion mimics (WLLMs), vegetative initiation lesion mimics (VILMs), and reproductive initiation lesion mimics (RILMs) according to the occurrence period of the lesion mimic mutations.
5. Disease Resistance of Rice Lesion Mimic Mutants
References
- Jiao, R.; Xu, N.; Hu, J.; Song, Z.L.; Hu, J.Q.; Rao, Y.C.; Wang, Y.X. Research progress on traits and molecular mechanisms of rice lesion mimic mutants. Chin. J. Rice Sci. 2018, 32, 285–295.
- Matin, M.N.; Pandeya, D.; Baek, K.H.; Dong, S.L.; Lee, J.H.; Kang, H.; Kang, S.G. Phenotypic and genotypic analysis of rice lesion mimic mutants. Plant Pathol. J. 2010, 26, 159–169.
- Xia, C.Y.; Yan, B.Y.; Cai, Y.F.; Wang, H.H.; Gong, Y.H. Advances in programmed cell death in plants. Lett. Biotechnol. 2008, 19, 296–298.
- Huang, Q.N.; Yang, Y.; Shi, Y.F.; Chen, J.; Wu, J.L. Research progress on spot leaf variation in rice. Chin. J. Rice Sci. 2010, 24, 108–115.
- McGrann, G.R.; Steed, A.; Burt, C.; Paul, N.; Brown, J.K. Differential effects of lesion mimic mutants in barley on disease development by facultative pathogens. J. Exp. Bot. 2015, 66, 3417–3428.
- Liu, G.; Wang, L.; Zhou, Z.; Leung, H.; Wang, G.L.; He, C. Physical mapping of a rice lesion mimic gene, Spl1, to a 70-kb segment of rice chromosome 12. Mol. Genet. Genom. 2004, 272, 108–115.
- Yamanouchi, U.; Yano, M.; Lin, H.; Ashikari, M.; Yamada, K. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc. Natl. Acad. Sci. USA 2002, 99, 7530–7535.
- Hoang, T.V.; Vo, K.T.X.; Rahman, M.M.; Choi, S.H.; Jeon, J.S. Heat stress transcription factor OsSPL7 plays a critical role in reactive oxygen species balance and stress responses in rice. Plant Sci. 2019, 289, 110273.
- Bai, J.T.; Zhu, X.D.; Wang, Q.; Zhang, J.; Chen, H.Q.; Dong, G.J.; Zhu, L.; Zheng, H.K.; Xie, Q.J.; Nian, J.Q.; et al. Rice TUTOU1 encodes a suppressor of camp receptor-like protein that is important for actin organization and panicle development. Plant Physiol. 2015, 169, 1179.
- Wu, C.; Bordeos, A.; Madamba, M.R.; Baraoidan, M.; Ramos, M.; Wang, G.L.; Leach, J.E.; Leung, H. Rice lesion mimic mutants with enhanced resistance to diseases. Mol. Genet. Genom. 2008, 279, 605–619.
- Wang, J.J.; Zhu, X.D.; Wang, L.Y.; Zhang, L.H.; Xue, Q.Z.; He, Z.H. Physiological and genetic analysis of rice lesion mimic mutants. J. Plant Physiol. Mol. Biol. 2004, 30, 331–338.
- Wang, Z.H.; Jia, Y.L. Induction and preliminary analysis of rice lesion mutant lmm1. J. Nucl. Agric. Sci. 2006, 20, 255–258.
- Tang, J.; Zhu, X.; Wang, Y.; Liu, L.; Xu, B.; Li, F.; Fang, J.; Chu, C. Semi-dominant mutations in the CC-NB-LRR-type R gene, NLS1, lead to constitutive activation of defense responses in rice. Plant J. 2011, 66, 996–1007.
- Mori, M.; Tomita, C.; Sugimoto, K.; Hasegawa, M.; Hayashi, N.; Dubouzet, J.G.; Ochiai, H.; Sekimoto, H.; Hirochika, H.; Kikuchi, S. Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol. Biol. 2007, 63, 847–860.
- Shirano, Y.; Kachroo, P.; Shah, J.; Klessig, D.F. A gain-of-function mutation in an Arabidopsis Toll Interleukin1 receptor-nucleotide binding site-leucine-rich repeat type R gene triggers defense responses and results in enhanced disease resistance. Plant Cell 2002, 14, 3149–3162.
- Danon, A.; Miersch, O.; Felix, G.; Camp, R.G.; Apei, K. Concurrent activation of cell death-regulating signaling pathways by singlet oxygen in Arabidopsis thaliana. Plant J. 2005, 41, 68–80.
- Jin, B.; Zhou, X.; Jiang, B.; Gu, Z.; Zhang, P.; Qian, Q.; Chen, X.; Ma, B. Transcriptome profiling of the spl5 mutant reveals that SPL5 has a negative role in the biosynthesis of serotonin for rice disease resistance. Rice 2015, 8, 18.
- Lin, A.; Wang, Y.; Tang, J.; Xue, P.; Li, C.; Liu, L.; Hu, B.; Yang, F.; Loake, G.J.; Chu, C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol. 2011, 158, 451–464.
- Dong, X. SA, JA, ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1998, 1, 316–323.
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14 (Suppl. 1), S153–S164.
- Shen, X.; Liu, H.; Yuan, B.; Li, X.; Xu, C.; Wang, S. OsEDR1 negatively regulates rice bacterial resistance via activation of ethylene biosynthesis. Plant Cell Environ. 2011, 34, 179–191.
- Jiang, C.J.; Shimono, M.; Maeda, S.; Inoue, H.; Mori, M.; Hasegawa, M.; Sugano, S.; Takatsuji, H. Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol. Plant-Microbe Interact. 2009, 22, 820–829.
- Tu, R.; Wang, H.; Liu, Q.; Wang, D.; Zhou, X.; Xu, P.; Zhang, Y.; Wu, W.; Chen, D.; Cao, L. Characterization and genetic analysis of the oshpl3 rice lesion mimic mutant showing spontaneous cell death and enhanced bacterial blight resistance. Plant Physiol. Biochem. 2020, 154, 94–104.
- Wang, Z.; Wang, Y.; Hong, X.; Hu, D.; Liu, C.; Yang, J.; Li, Y.; Huang, Y.; Feng, Y.; Gong, H. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defence responses in rice. J. Exp. Bot. 2015, 66, 973–987.
- Sakuraba, Y.; Rahman, M.L.; Cho, S.; Kim, Y.; Koh, H.; Yoo, S.; Paek, N. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J. 2013, 74, 122–133.
- Qiao, Y.; Jiang, W.; Lee, J.; Park, B.S.; Choi, M.S.; Piao, R.; Woo, M.O.; Roh, J.H.; Han, L.; Paek, N.C. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol. 2010, 185, 258–274.
- Broker, L.E.; Kruyt, F.A.; Giaccone, G. Cell Death Independent of Caspases: A Review. Clin. Cancer Res. 2005, 11, 3155–3162.
- Zeng, L.R.; Qu, S.; Bordeos, A.; Yang, C.; Baraoidan, M.; Yan, H.; Xie, Q.; Nahm, B.H.; Leung, H.; Wang, G.L. Spotted leaf 11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell 2004, 16, 2795–2808.
- Ma, H.; Li, j.; Ma, L.; Xue, Y.; Yin, P.; Xiao, J.; Wang, S. Pathogen-inducible OsMPKK10.2-OsMPK6 cascade phosphorylates the Raf-like kinase OsEDR1 and inhibits its scaffold function to promote rice disease resistance. Mol. Plant 2021, 14, 620–632.
- Manosalva, P.M.; Bruce, M.; Leach, J.E. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance. Plant J. 2011, 68, 777–787.
- Wang, L.; Pei, Z.; Tian, Y.; He, C. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol. Plant-Microbe Interact. 2005, 18, 375–384.
- Xia, S.S.; Cui, Y.; Li, F.F.; Tan, J.; Ling, Y.H. Phenotypic identification and gene mapping of premature senescence mutant lmps1 in rice. Acta Agron. Sin. 2019, 45, 46–54.
- Wang, J.J.; Zhang, L.X.; Wang, L.Y.; Zhang, L.H.; Zhu, C.N.; He, Z.H.; Jin, Q.S.; Fan, H.H.; Xin, Y. Induced responses of rice Lesion Resembling Disease mutants to light and temperature. Sci. Agric. Sin. 2010, 43, 2039–2044.
- Yuan, Y.; Zhong, S.; Li, Q.; Zhu, Z.; Lou, Y.; Wang, L.; Wang, J.; Wang, M.; Li, Q.; Yang, D.; et al. Functional analysis of rice NPR1-like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol. J. 2007, 5, 313–324.
- Takahashi, A.; Agrawal, G.K.; Yamazaki, M.; Onosato, K.; Miyao, A.; Kawasaki, T.; Shimamoto, K.; Hirochika, H. Rice Pti1a negatively regulates RAR1-dependent defense responses. Plant Cell 2007, 19, 2940–2951.
- Park, C.J.; Peng, Y.; Chen, X.; Dardick, C.; Ruan, D.; Bart, R.; Canlas, P.E.; Ronald, P.C. Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity. PLoS Biol. 2008, 6, e231.
- Yamaguchi, T.; Kuroda, M.; Yamakawa, H.; Ashizawa, H.; Hirayae, K.; Kurimoto, L.; Shinya, T.; Shibuya, N. Suppression of a phospholipase D gene, OsPLDβ1, activates defense responses and increases disease resistance in rice. Plant Physiol. 2009, 150, 308–319.
- Fujiwara, T.; Maisonneuve, S.; Isshiki, M.; Mizutani, M.; Chen, L.; Wong, H.L.; Kawasaki, T.; Shimamoto, K. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J. Biol. Chem. 2010, 285, 11308–11313.
- Cui, Y.; Peng, Y.; Zhang, Q.; Xia, S.; Ruan, B.; Xu, Q.; Yu, X.; Zhou, T.; Liu, H.; Zeng, D.; et al. Disruption of EARLY LESION LEAF 1, encoding a cytochrome P450 monooxygenase, induces ROS accumulation and cell death in rice. Plant J. 2021, 105, 942–956.
- Sun, C.; Liu, L.; Tang, J.; Lin, A.; Zhang, F.; Fang, J.; Zhang, G.; Chu, C. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J. Genet. Genom. 2011, 38, 29–37.
- Chen, X.; Hao, L.; Pan, J.; Zheng, X.; Jiang, G.; Yang, J.; Gu, Z.; Qian, Q.; Zhai, W.; Ma, B. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol. Breed. 2012, 30, 939–949.
- Liu, X.; Li, F.; Tang, J.; Wang, W.; Zhang, F.; Wang, G.; Chu, J.; Yan, C.; Wang, T.; Chu, C.; et al. Activation of the Jasmonic Acid Pathway by Depletion of the Hydroperoxide Lyase OsHPL3 Reveals Crosstalk between the HPL and AOS Branches of the Oxylipin Pathway in Rice. PLoS ONE 2012, 7, e50089.
- Undan, J.R.; Tamiru, M.; Abe, A.; Yoshida, K.; Kosugi, S.; Takagi, H.; Yoshida, K.; Kanzaki, H.; Saitoh, H.; Fekih, R.; et al. Mutation in OsLMS, a gene encoding a protein with two double-stranded RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L.). Genes Genet. Syst. 2012, 87, 169–179.
- Vega-Sánchez, M.E.; Verhertbruggen, Y.; Christensen, U.; Chen, X.; Sharma, V.; Varanasi, P.; Jobling, S.A.; Talbot, M.; White, R.G.; Joo, M.; et al. Loss of Cellulose synthase-like F6 function affects mixed-linkage glucan deposition, cell wall mechanical properties, and defense responses in vegetative tissues of rice. Plant Physiol. 2012, 159, 56–69.
- Jiao, B.B.; Wang, J.J.; Zhu, X.D.; Zeng, L.J.; Li, Q.; He, Z.H. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice. Mol. Plant 2012, 5, 205–217.
- Fekih, R.; Tamiru, M.; Kanzaki, H.; Abe, A.; Yoshida, K.; Kanzaki, E.; Saitoh, H.; Takagi, H.; Natsume, S.; Undan, J.R.; et al. The rice (Oryza sativa L.) LESION MIMIC RESEMBLING, which encodes an AAA-type ATPase, is implicated in defense response. Mol. Genet. Genom. 2015, 290, 611–622.
- You, Q.; Zhai, K.; Yang, D.; Yang, W.; Wu, J.; Liu, J.; Pan, W.; Wang, J.; Zhu, A.; Jian, Y.; et al. An E3 Ubiquitin Ligase-BAG Protein Module Controls Plant Innate Immunity and Broad-Spectrum Disease Resistance. Cell Host Microbe 2016, 20, 758–769.
- Yang, X.; Gong, P.; Li, K.; Huang, F.; Cheng, F.; Pan, G. A single cytosine deletion in the OsPLS1 gene encoding vacuolar-type H+-ATPase subunit A1 leads to premature leaf senescence and seed dormancy in rice. J. Exp. Bot. 2016, 67, 2761–2776.
- Sun, L.; Wang, Y.; Liu, L.L.; Wang, C.; Gan, T.; Zhang, Z.; Wang, Y.; Wang, D.; Niu, M.; Long, W.; et al. Isolation and characterization of a spotted leaf 32 mutant with early leaf senescence and enhanced defense response in rice. Sci. Rep. 2017, 7, 41846.
- Wang, S.; Lei, C.; Wang, J.; Ma, J.; Tang, S.; Wang, C.; Zhao, K.; Tian, P.; Zhang, H.; Qi, C.; et al. SPL33, encoding an eEF1A-like protein, negatively regulates cell death and defense responses in rice. J. Exp. Bot. 2017, 68, 899–913.
- Liu, Q.; Ning, Y.; Zhang, Y.; Yu, N.; Zhao, C.; Zhan, X.; Wu, W.; Chen, D.; Wei, X.; Wang, G.L.; et al. OsCUL3a negatively regulates cell death and immunity by degrading OsNPR1 in rice. Plant Cell 2017, 29, 345–359.
- Qin, P.; Fan, S.; Deng, L.; Zhong, G.; Zhang, S.; Li, M.; Chen, W.; Wang, G.; Tu, B.; Wang, Y.; et al. LML1, encoding a conserved eukaryotic release factor 1 protein, regulates cell death and pathogen resistance by forming a conserved complex with SPL33 in rice. Plant Cell Physiol. 2018, 59, 887–902.
- Fan, J.; Bai, P.; Ning, Y.; Wang, J.; Shi, X.; Xiong, Y.; Zhang, K.; He, F.; Zhang, C.; Wang, R.; et al. The Monocot-Specific Receptor-like Kinase SDS2 controls cell death and immunity in rice. Cell Host Microbe 2018, 23, 498–510.e5.
- Ma, J.; Wang, Y.; Ma, X.; Meng, L.; Jing, R.; Wang, F.; Wang, S.; Cheng, Z.; Zhang, X.; Jiang, L.; et al. Disruption of gene SPL35, encoding a novel CUE domain-containing protein, leads to cell death and enhanced disease response in rice. Plant Biotechnol. J. 2019, 17, 1679–1693.
- Sathe, A.P.; Su, X.; Chen, Z.; Chen, T.; Wei, X.; Tang, S.; Zhang, X.B.; Wu, J.L. Identification and characterization of a spotted-leaf mutant spl40 with enhanced bacterial blight resistance in rice. Rice 2019, 12, 68.
- Zhang, Y.; Liu, Q.; Zhang, Y.; Chen, Y.; Yu, N.; Cao, Y.; Zhan, X.; Cheng, S.; Cao, L. LMM24 Encodes Receptor-Like Cytoplasmic Kinase 109, which regulates cell death and defense responses in rice. Int. J. Mol. Sci. 2019, 20, 3243.
- Du, D.; Zhang, C.; Xing, Y.; Lu, X.; Cai, L.; Yun, H.; Zhang, Q.; Zhang, Y.; Chen, X.; Liu, M.; et al. The CC-NB-LRR OsRLR1 mediates rice disease resistance through interaction with OsWRKY19. Plant Biotechnol. J. 2021, 19, 1052–1064.


