Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Duy T. Nguyen | -- | 3079 | 2022-07-04 14:19:02 | | | |
| 2 | Beatrix Zheng | -25 word(s) | 3054 | 2022-07-05 03:34:17 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Nguyen, D.T.; Ogando-Rivas, E.; Liu, R.; Wang, T.; Rubin, J.; Jin, L.; Tao, H.; Sawyer, W.W.; Nguyen, T.Y.N.; Mendez-Gomez, H.R.; et al. CAR T Cell Locomotion in Solid Tumor Microenvironment. Encyclopedia. Available online: https://encyclopedia.pub/entry/24794 (accessed on 14 June 2026).
Nguyen DT, Ogando-Rivas E, Liu R, Wang T, Rubin J, Jin L, et al. CAR T Cell Locomotion in Solid Tumor Microenvironment. Encyclopedia. Available at: https://encyclopedia.pub/entry/24794. Accessed June 14, 2026.
Nguyen, Duy T., Elizabeth Ogando-Rivas, Ruixuan Liu, Theodore Wang, Jacob Rubin, Linchun Jin, Haipeng Tao, William W. Sawyer, Tran Yen Nhi Nguyen, Hector R. Mendez-Gomez, et al. "CAR T Cell Locomotion in Solid Tumor Microenvironment" Encyclopedia, https://encyclopedia.pub/entry/24794 (accessed June 14, 2026).
Nguyen, D.T., Ogando-Rivas, E., Liu, R., Wang, T., Rubin, J., Jin, L., Tao, H., Sawyer, W.W., Nguyen, T.Y.N., Mendez-Gomez, H.R., Cascio, M., Mitchell, D.A., Huang, J., Sawyer, W.G., Sayour, E.J., & Castillo, P. (2022, July 04). CAR T Cell Locomotion in Solid Tumor Microenvironment. In Encyclopedia. https://encyclopedia.pub/entry/24794
Nguyen, Duy T., et al. "CAR T Cell Locomotion in Solid Tumor Microenvironment." Encyclopedia. Web. 04 July, 2022.
Copy Citation
The promising outcomes of chimeric antigen receptor (CAR) T cell therapy in hematologic malignancies potentiates its capability in the fight against many cancers. Nevertheless, this immunotherapy modality needs significant improvements for the treatment of solid tumors. Researchers have incrementally identified limitations and constantly pursued better CAR designs. However, even if CAR T cells are armed with optimal killer functions, they must overcome and survive suppressive barriers imposed by the tumor microenvironment (TME). The ability of CAR T cells to efficiently migrate to the tumor site, infiltrate suppressive barriers, and survive the harsh TME represents a crucial prerequisite for carrying out the anti-tumor function.
CAR T cells
solid tumors
T cell migration
trafficking
tumor microenvironment
immunotherapy
adoptive T cell therapy
3D in vitro models
1. The Immunosuppressive Tumor Microenvironment and Physical Barriers Favorable to Resilient Tumor Growth
Circulating chimeric antigen receptor (CAR) T cells must gain sufficient traction and adhesion against receptors on the endothelial wall to initiate extravasation (Figure 1). However, secretion of angiogenic factors VEGF and bFGF during tumorigenesis leads to insufficient expression of adhesion molecules—such as intercellular adhesion molecule 1 (ICAM-1), 2 (ICAM-2), and vascular cell adhesion molecule 1 (VCAM-1)—on endothelial cells which prevents efficient T cell engagement [1]. Adhesion molecules expressed on both immune cells and endothelial walls modulate interactions necessary for transendothelial migration (TEM). Although mechanisms of TEM have been reported extensively, identification of a comprehensible approach to enhance the extravasation of T cells remains elusive. Instead, T cells have been engineered to target tumor-associated endothelial receptors such as VEGFR2 to disrupt tumor-supporting vasculature, enhance T cell trafficking, and deprive the tumor of nutrient and oxygen supplies [2][3]. This approach revealed the efficient killing of in vitro tumor spheroids [3] and prolonged survival in animal models [2][4][5].
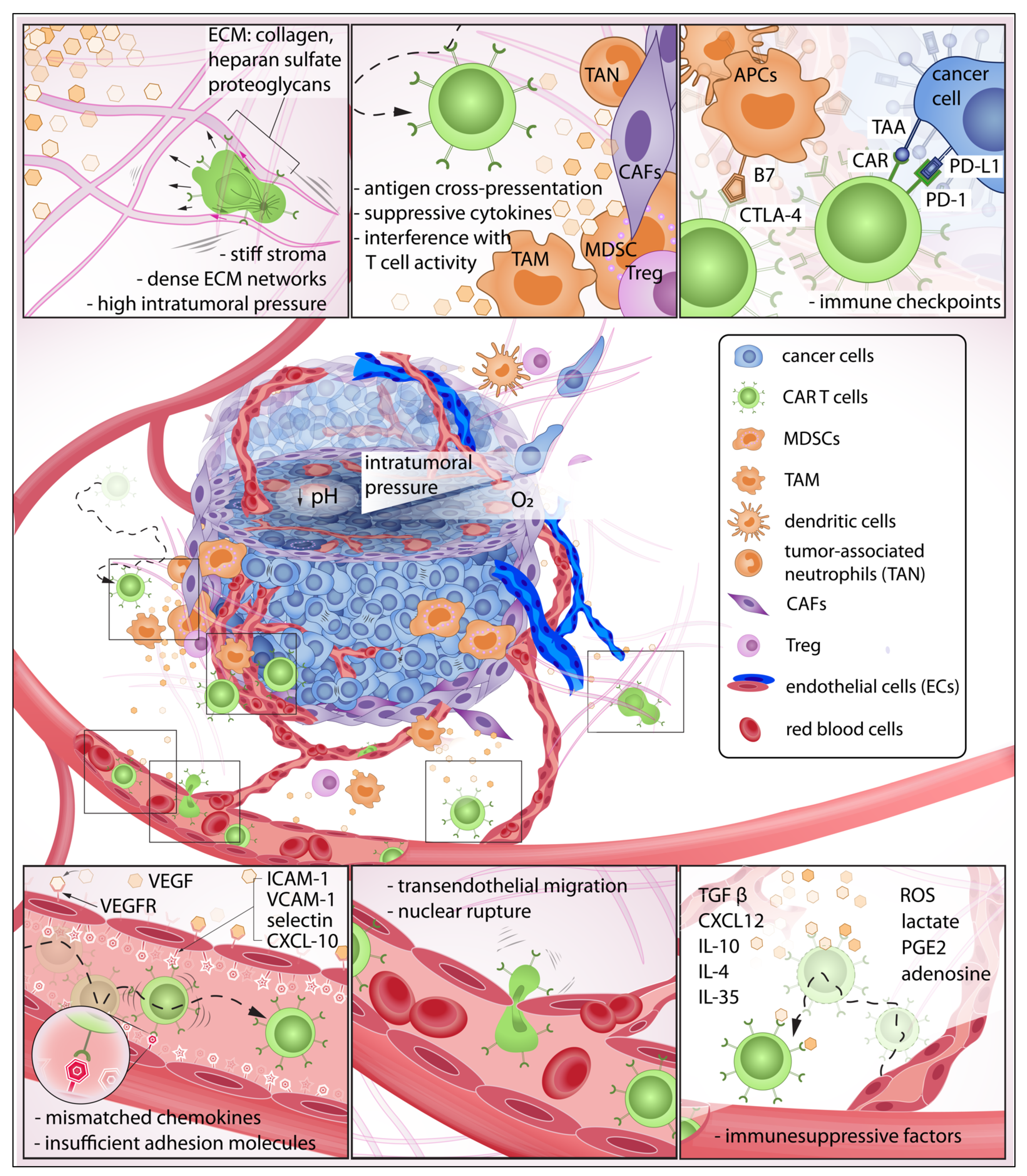
Figure 1. The exhausting journey of CAR T cells in the tumor microenvironment (TME): In this journey, CAR T cells must be able to detect chemokines at the tumor site, effectively roll and adhere to the blood vessel wall, initiate transendothelial migration, and invade the tumor stroma. Here, the immune cells must overcome various biochemical and physical barriers to then encounter pro-tumor cells that suppress CAR T cell activity. Some CAR T cells may eventually make contact with target cancer cells that express abundant immune checkpoints, further reducing anti-tumor function.
CAR T cells must traverse the endothelial junction and navigate through the tumor stroma that is inherently armed with abundant immunosuppressive factors to reach target cancer cells. The TME has long been identified as an active contributor to cancer progression [6]. In this neoplastic microenvironment, different cell types interact with each other and the surrounding ECM to orchestrate a constantly evolving habitat that is favorable to resilient tumor growth [7][8][9] and deleterious to immune function [10][11][12][13] (Figure 2). For instance, CAR T cells—upon successful infiltration of the TME—demonstrated a rapid loss of function [14]. Infiltrating CAR T isolated from xenograft tumors and cultured in vitro for 24 h regained killing ability superior to those freshly isolated [14]. This emphasizes the immunosuppressive influences of the TME that suppress CAR T anti-tumor activity. Furthermore, the stromal, immune, and cancer cells constantly remodel the TME in response to the surrounding biochemical and biophysical cues [7][15][16][17][18] (Figure 2). Unlike hematologic malignancies—which can be freely accessed by CAR T cells—solid tumors have a complex three-dimensional structure wherein malignant cells are more difficult to access [19][20][21][22]. One hallmark of tumor progression is the elevated stiffening of the ECM due to increased matrix crosslinking, collagen deposition, and fiber alignment which regulate cell migration, proliferation, and apoptosis via mechanotransduction [23][24][25][26][27]. The stiffening effect is often caused by collagen deposition and crosslinking, mediated by increased secretion of lysyl oxidase (LOX) [18][28] and overproduction of other ECM components such as heparan sulfate proteoglycans (HSPGs) [29][30]. Due to this physical constraint, primary tumor growth and cell migration within the TME are greatly dependent on matrix metalloproteinases (MMP) and heparinase enzymes to degrade and reorganize the crosslinked networks [31][32][33][34][35][36]. However, T cells do not often secrete enzymes to degrade ECM, but rather choose the path of least resistance [37] or follow contact guidance imposed by architectural features of the surrounding ECM [38][39]. Therefore, densely packed and oriented stromal fibers have been reported to impose significant challenges to T cell infiltration [40].
Figure 2. Overview of the immunosuppressive tumor microenvironment (TME) and important contributors to tumor progression: Within the TME, there is a dynamic relationship between peritumoral and intratumoral components that include immune cells, cancer cells, and the stromal elements often exhibiting context-dependent functionality. Different components of the TME carry out pro- or anti-tumor functions, and such polarizations are caused by reciprocal interactions with physical and biochemical cues from the surroundings. The polarized functions are not distinctive but rather heterogenous contributing to an immunosuppressive TME [6][7][8][16][17][23][27][28][34][36][41][42][43][44][45][46][47][48][49][50][51][52][53][54][55][56][57][58][59][60][61][62][63].
Tumor growth is critically dependent on angiogenesis [15][64][65] which is promoted in an MMP–dependent manner [31]. Given nutrient support from neovessel networks, the growing tumor continues to modify its microenvironment through elevation of growth-induced solid stress [66][67], interstitial fluid pressure [68][69], enzymatic secretion [28][31], and cellular contractility onto the ECM [56][59]. One example of ECM modification is the physical alignment of collagen fibers due to cell-induced strain stiffening [16][57][70] from cancer cells and their stromal neighbors such as cancer-associated fibroblasts (CAFs) [16][52][55]. This reorganization further stiffens the TME, suppresses immune activity and mobility [71][72], and promotes local invasion of cancer cells along the aligned fibers [43][61][71][73][74][75][76]. In addition, intratumoral pressure caused by growth-induced solid stress and interstitial fluid pressure obstruct delivery of therapeutic agents and trafficking of immune cells to the solid tumors [77]. Elevated interstitial fluid pressure—reported in most solid cancers [78]—is mainly caused by leaky blood vessels, impaired lymphatic transport, and hyaluronan swelling [60][79]. The fluid pressure contributes to an increasing growth-induced solid stress and presents a non-trivial barrier to immune infiltration and uptake of therapeutic agents [78][80]. An in vitro study demonstrated that an interstitial fluid pressure ≥ 1 KPa—simulated by hydrostatic pressure—is sufficient to physically hindered antigen-specific T cell infiltration into tumor site [81]. Furthermore, growth-induced solid stress (≥2 kPa) —while impeding proliferation of cancer cells [66][82] —can collapse blood vessels [60] and contribute to vasculature abnormalities and subsequent increase in interstitial pressure [59]. The reciprocal relationship further escalates total intratumoral pressure, exacerbates hypoxia, and increases pH and metabolic waste accumulation. TME determinants—such as hypoxia [83] and TGFβ [51]—stimulate the production of ECM [11][53][84][85] and LOX [86] to further reinforce the physical barriers that impede CAR T infiltration [21][87]. The feedback loop continually contributes an immunosuppressive TME responsible for tumor pathogenic traits, its progression [28][88][89][90], subtle microevolution [91][92][93][94][95], and metastasis [61][62][63][96][97][98][99]. To overcome these challenges, therapeutic agents for vascular normalization [100] and degradation of ECM components—such as hyaluronidases [101], collagens [102], and LOX inhibition [103]—have been developed to alleviate intratumoral pressure and improve perfusion transport of therapeutic agents and immune cell infiltration [104].
2. Immunosuppressive Chemokines: The Invisible Barrier against CAR T Infiltration
The recruitment of CAR T cells to the TME, upon systemic or regional administration, depends largely on efficient chemotactic migration. Without biochemical hints, the probability that CAR T cells localize and migrate to a solid tumor would be low. Within a TME, the local accumulation of suppressive chemokines and cytokines—such as CXCL12 [105][106], TGFβ [51][107], IL-10 [108][109][110], IL-4 [111], IL-35 [112][113] and other factors (e.g., reactive oxygen species (ROS), lactate [114], prostaglandin E2 (PGE2), and adenosine [115])—constitute a first major barrier of immune suppression. The ability to sense and migrate towards a tumor-specific site is a critical prerequisite to immune extravasation and infiltration; however, as a physiological protective mechanism, tumors produce insufficient chemoattractant ligands for T cells. Therefore, there have been significant efforts to increase CAR T chemotaxis and extravasation that endow the local delivery of favorable chemokines and cytokines (e.g., CXCL11 [116], RANTES, and IL-15 [117]); these efforts include the use of intratumorally delivered oncolytic virus to improve CAR T cell recruitment and anti-tumor activity with resulting better survival [116][117][118]. Alternatively, there have been promising strategies that employ CAR T cells as cytokine carriers to promote infiltration and maintenance of a healthy TILs population. CAR T cells—engineered to secrete proinflammatory cytokines, IL-12 [4][119], IL-18 [120], IL-7 [121], and CCL19 [122] —have been reported to augment autocrine stimulation and potentiate paracrine signaling advantageous for survival, persistence, and recruitment of endogenous immune cells.
To enhance locomotion in the TME, CAR T cells are engineered to express receptors specific to tumor-derived chemokines. This approach has shown efficient chemotactic migration and demonstrated remarkable tumor regression in both in vitro and in vivo models. T cells transduced with a retroviral vector to express CXCR2, a receptor of chemokine CXCL1, were able to enhance direct migration toward tumor-derived chemokine [123]. In another study, CCR2—a receptor of chemokine CCL2, co-expressed with a chimeric antigen receptor targeting tumor antigen GD2 [124] (expressed on neuroblastoma) and mesothelin [125] (expressed on malignant pleural mesotheliomas)—demonstrated a significant increase (more than 10-fold) in homing and anti-tumor activity. The researchers' group, Jin et al., has developed CD70-directed CXCR1 or CXCR2-modified CAR T cells that can co-opt IL-8 with robust antitumor activity and long-lasting immunologic memory against glioblastoma, ovarian, and pancreatic cancer xenograft models [126]. This approach has received FDA IND approval soon to be evaluated in a first clinical trial in humans [126][127]. Furthermore, CAR T cells have been designed to co-express dominant-negative receptors of factors (e.g., TGFβ [128], PD-L1 [129]) to resist immune-inhibitory signals in the TME or co-express inverted cytokine receptor [111][130] to reverse suppressive cytokine functions and stimulate anti-tumor activity. In a preclinical study for breast cancer, CAR T cells targeting MUC1 co-expressed with an inverted cytokine receptor comprised of an IL-4 exodomain linked to an IL-7 endodomain reversed inhibitory signals from IL-4 present in the TME and demonstrated durable T cells infiltration and memory formation [131].
3. Pro-Tumor Stromal and Immune Cells Are Active Contributors to Immune Suppression
Once CAR T cells invade chemorepellent and physical barriers of the TME, they encounter stromal and immune cell populations that often exhibit context-dependent functionality. Cancer-associated fibroblasts (CAFs) are present in all solid tumors and represent a major component of the reactive tumor stroma integral to the development and maintenance of a complex TME [132]. CAFs are responsible for abundant production of ECM, crosslinking enzymes, and cytokines that fortify the tumor and constitute a complex and heterogenous barrier against immune attacks [133]. CAFs are major contributors to the production of immunosuppressive cytokines, such as TGFβ, which have also been shown to interfere with T cell migration and infiltration into tumors [51][134]. CAFs have been reported to cross-present antigen and kill CD8+T cells in an antigen-dependent manner via PD-L2 and FASL expression [135] and recruit other inhibitory immune cells to participate in suppressing anti-tumor function [49]. Direct immunotherapeutic attenuation of CAF activity shows promising improvement of T cell trafficking and infiltration [136]. Kakarla et al. developed CAR T cells targeting fibroblast activation protein (FAP), demonstrating a significant reduction in FAP-positive stromal cells and tumor growth in murine models. A combined application of FAP-CAR T and tumor-antigen CAR T cells resulted in anti-tumor activity superior to treatment with either CAR T alone [137]. Despite generally being known for pro-tumor function, the role of CAFs in tumor progression is not totally understood. Depletion of CAFs accelerated pancreatic ductal adenocarcinoma progression and reduced survival in transgenic mice models [138]. Although each cellular component of a TME exhibits a supportive role in tumor progression, their existing anti-tumor functions further complicate therapeutic strategies.
Tumor initiation and progression lead to inflammatory reactions that trigger recruitment and repair mechanisms by innate and adaptive immune response [139]. In the complex TME, cancer, stromal, and immune cell populations continuously evolve. The pro-tumor immune subsets within the TME generally comprise tumor-associated macrophages (TAM) [44][140], myeloid-derived suppressor cells (MDSCs), tumor-associated neutrophils (TAN), and regulatory T cells (Treg) [42][50][141]. Macrophages and dendritic cells (DCs) are important myeloid cells of the innate immune system. TAM often adopt an M2- macrophage phenotype [142] which is anti-inflammatory and pro-tumorigenic [143][144]. TAMs secrete CCL2, which recruits and activates additional TAMs. M2 TAMs have revealed pro-tumor functions and immune suppressive effects via secretion of epidermal and angiogenic factors, IL10 and TGFβ [145]. CAR T cells targeting M2-like TAMs in murine ovarian carcinoma, colon adenocarcinoma, and melanoma models reprogrammed the TME with enrichment of pro-inflammatory monocytes and activated CD8+T cells [146].
MDSCs, a heterogeneous population of immature myeloid cells, are recruited into the tumor by various chemokines CCL1, CCL2, CCL5, or CXCL5 [141][147]. MDSCs can be divided into two major subtypes: granulocytic and monocytic. Granulocytic MDSCs secrete reactive oxygen species (ROS), whereas monocytic MDSCs produce increased levels of nitric oxide derivatives resulting in decreased T cell immune responses [48][148]. A MUC1-directed CAR T cell approach concomitantly targeting tumor necrosis factor-related apoptosis-inducing ligand receptor 2 (TR2) expressed on MDSCs demonstrated enhanced antitumor tumor activity in breast cancers enriched with MDSCs and TME remodeling [149]. In addition, there is a growing body of data on the anti-antitumor immunity of TANs. The role of TANs seems to vary based on the type of solid tumors [150]. It has been described that TANs recruit MDSCs, TAMs, and Tregs to TME through secretion of IL-4, IL-10, arginine-1, and ROS which have inhibitory effects on cytotoxic T cells [150].
Tregs are cells born in the thymus and express CD4+ FoxP3. Tregs within the TME can act antagonistically to effector T cells by secreting a number of cytokines (e.g., IL10, IL35, TGFβ), upregulating cytotoxic T lymphocyte antigen 4 (CTLA4), and inhibiting CD80/CD86 co-stimulatory pathways, by competing for binding of IL-2 [151]. An agonist antibody specific against glucocorticoid-induced TNFR-related receptor (GITR) decreased Treg mediated immunosuppression which correlated with augmented anti-glioblastoma immune response [152]. Once in the TME, CAR T cells must overcome checkpoint inhibitory signals. Checkpoints (e.g., PD-L1, GITR) are physiological brakes that prevent autoimmunity [146][147][148]. However, cancer cells abuse such mechanisms to avoid immune surveillance, and the complex TME further supports cancer cells by employing multifactorial and reciprocal pathways advantageous to immune suppression. For instance, IFNγ release during CAR T cell activation and killing of tumor cells induces upregulation of immune checkpoint proteins (e.g., PD-L1) which attenuate the anti-tumor function of cytotoxic T cells [153]. On the other hand, cancer-associated cells can be reprogrammed to target cancer cells. Reinhard et al. revealed nanoparticles delivering RNA encoding for claudin 6—a developmental antigen—can transduce antigen presenting cells to express the immunological targets and enhance CAR T expansion and tumor regression in a mouse model [154].
4. Physical Confinement and Mechanical Properties of Cell Nucleus Modulate T Cell Locomotion
T cell locomotion within the TME is an important aspect of immuno-oncology and has been extensively studied [38][39][155][156]. T cells are capable of dynamically adapting to various modes of migration depending on ECM composition within the TME [38][156][157]. However, the mechanism behind this adaptation has not been clearly explained. A better understanding of T cell migration will help enhance CAR T trafficking to solid tumors. The modes of cell migration are often regulated by the density of binding sites on substrate [158], proteolytic activity [159], actomyosin contractility [38][160], microtubule stability [39], and geometrical and mechanical properties of the confined surroundings [39][41][155][157][161]. T cells are fast migrators, for they need rapid scanning abilities to carry out efficient immunological response. Two-photon microscopy imaging of intact mouse lymph nodes revealed that T cells can migrate at an average speed of 10 µm min−1 and peak at 25 µm min−1 [162]. Unlike cancer and stromal cells that actively secrete protease enzymes to degrade the ECM [47] and migrate via focal adhesion mediated attachment, T cells do not need mature focal adhesions and prefer to explore the TME via paths of least resistance [156][163][164]. In other words, leukocytes probe their surroundings and selectively squeeze through accessible pores without proteolytically breaking down ECM to create paths for migration. Although capable of integrin-dependent migration, T cells only form short-lived adhesion complexes to facilitate rapid detachment and engagement onto the next location [165]. The most common mode of T cell migration is amoeboid which employs coordinated membrane extrusion, cell contractility, and contact-induced traction in confined spaces to move forward [156][165]. Amoeboid migration is critically dependent on cortical contractility regulated via the Rho/ROCK pathway and myosin II activity [38]. It has been reported that inducing actomyosin contractility by increasing myosin II or activation of the Rho/ROCK pathway is sufficient to transform cell mode of migration into amoeboid [160][166][167]. Besides cell contractility, proper geometrical confinement is an essential determinant of amoeboid migration [157]. T cells protrude oscillatory leading-edge membrane blebs into narrow pores, probe for potential paths, and generate contraction-mediated retrograde actin flow beneath the cell membranes to transmit equal and opposite frictional forces onto the substrate to drive the cell forward [38][156].
The compliance of the cells can better facilitate constricted migration [43]. The deformability of a cell depends significantly on the mechanical property of its nucleus [168][169]. During transendothelial migration through tight spaces, CAR T may be subjected to nuclear rupture which could severely impact cell function [170]. Major determinants of nuclear stiffness are lamin A/C content and chromatin decondensation which are mechanically regulated by force transmission between cell–cell and cell–matrix interaction via cytoskeleton and LINC complex [58]. Lamin A is known to be responsible for the viscous portion of the nucleus which leads to plastic deformation post-migration, whereas lamin B is responsible for the elastic portion promoting shape recovery after deformation [171][172]. Although high lamin A expression impedes migration through small pores (<3 µm in dia.), it promotes cell survival against stress-induced apoptosis by upregulation of DNA damage repair protein, HSP90 [173][174].
Furthermore, cells are well-known mechanosensors [24][58][175][176][177][178][179] and can alter their mechanical properties and mode of migration according to the stiffness of local surroundings [39][43][45][46]. On a soft substrate, lamin A/C phosphorylation is increased; whereas on stiff substrate, myosin II-generated tension promotes dephosphorylation and stabilization of lamin A/C levels which in turn enhance nuclear stiffness [180]. Moreover, mechanotransduction is a key regulator in T cell activation [181][182] and cytotoxic activity [179]. In the context of T cell migration, the mechanical stiffness of the substrate is important for durotaxis and contact guidance-directed locomotion in 3D [39]. T cells apply traction forces (100 pN) via TCR to physically probe the rigidity of their surroundings [177] and migrate along with the aligned ECM fibers [183]. Although matrix optimal confinement and alignment could induce rapid T cell amoeboid migration, there is a limit to which cells are physically trapped and suppressed. For instance, high-density collagen deposition and crosslinking have been shown to suppress proliferation and anti-tumor activity of infiltrating CD8+ T cells in triple-negative breast cancer tissue explants [72]. Another study shows that a densely packed and oriented stromal fibers at the perivascular niche and around the tumor impose a significant challenge to T cell infiltration [40]. Recently, Caruana et al. developed CAR T cells expressing heparinase to degrade HSPGs and promote tumor infiltration. In xenografted mouse models, the strategy improved T cell capability to degrade ECM, infiltrate, and carry out antitumor activity [35].
References
- Bellone, M.; Calcinotto, A. Ways to enhance lymphocyte trafficking into tumors and fitness of tumor infiltrating lymphocytes. Front. Oncol. 2013, 3, 231.
- Chinnasamy, D.; Yu, Z.; Theoret, M.R.; Zhao, Y.; Shrimali R, R.K.; Morgan, R.A.; Feldman, S.A.; Restifo, N.P.; Rosenberg, S.A. Gene therapy using genetically modified lymphocytes targeting VEGFR-2 inhibits the growth of vascularized syngenic tumors in mice. J. Clin. Investig. 2010, 120, 3953–3968.
- Englisch, A.; Altvater, B.; Kailayangiri, S.; Hartmann, W.; Rossig, C. VEGFR2 as a target for CAR T cell therapy of Ewing sarcoma. Pediatr. Blood Cancer 2020, 67, e28313.
- Chinnasamy, D.; Yu, Z.; Kerkar, S.P.; Zhang, L.; Morgan, R.A.; Restifo, N.P.; Rosenberg, S.A. Local delivery of interleukin-12 using T cells targeting VEGF receptor-2 eradicates multiple vascularized tumors in mice. Clin. Cancer Res. 2012, 18, 1672–1683.
- Chinnasamy, D.; Tran, E.; Yu, Z.; Morgan, R.A.; Restifo, N.P.; Rosenberg, S.A. Simultaneous targeting of tumor antigens and the tumor vasculature using t lymphocyte transfer synergize to induce regression of established tumors in mice. Cancer Res. 2013, 73, 3371–3380.
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674.
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437.
- Kai, F.B.; Drain, A.P.; Weaver, V.M. The Extracellular Matrix Modulates the Metastatic Journey. Dev. Cell 2019, 49, 332–346.
- Napoli, M.; Flores, E.R. Beware of thy neighbor: Senescent cancer cells feast on adjacent ells to persist. J. Cell Biol. 2019, 218, 3535–3536.
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200.
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: Implications for fibrotic diseases and cancer. Dis. Model. Mech. 2011, 4, 165–178.
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801.
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67.
- Moon, E.K.; Wang, L.C.; Dolfi, D.V.; Wilson, C.B.; Ranganathan, R.; Sun, J.; Kapoor, V.; Scholler, J.; Puré, E.; Milone, M.C.; et al. Multifactorial T-cell hypofunction that is reversible can limit the efficacy of chimeric antigen receptor-transduced human T cells in solid tumors. Clin. Cancer Res. 2014, 20, 4262–4273.
- Gustave, I.; Albini, A. Contribution to Tumor Angiogenesis From innate immune Cells within the Tumor Microenvironment: Implications for immunotherapy. Front. Immunol. 2018, 9, 527.
- Van Helvert, S.; Friedl, P. Strain Stiffening of Fibrillar Collagen during Individual and Collective Cell Migration Identified by AFM Nanoindentation. ACS Appl. Mater. Interfaces 2016, 8, 21946–21955.
- Ahmadzadeh, H.; Webster, M.R.; Behera, R.; Jimenez, A.M.; Wirtz, D. Modeling the two-way feedback between contractility and matrix realignment reveals a nonlinear mode of cancer cell invasion. Proc. Natl. Acad. Sci. USA 2017, 114, E1617–E1626.
- Butcher, D.T.; Alliston, T.; Weaver, V.M. A tense situation: Forcing tumour progression. Nat. Rev. Cancer 2009, 9, 108–122.
- D’Aloia, M.M.; Zizzari, I.G.; Sacchetti, B.; Pierelli, L.; Alimandi, M. CAR-T cells: The long and winding road to solid tumors review-article. Cell Death Dis. 2018, 9, 282.
- Li, J.; Li, W.; Huang, K.; Zhang, Y.; Kupfer, G.; Zhao, Q. Chimeric antigen receptor T cell (CAR-T) immunotherapy for solid tumors: Lessons learned and strategies for moving forward. J. Hematol. Oncol. 2018, 11, 22.
- Yong, C.S.M.; Dardalhon, V.; Devaud, C.; Taylor, N.; Darcy, P.K.; Kershaw, M.H. CAR T-cell therapy of solid tumors. Immunol. Cell Biol. 2017, 95, 356–363.
- Sterner, R.C.; Sterner, R.M. CAR-T cell therapy: Current limitations and potential strategies. Blood Cancer J. 2021, 11, 69.
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.T.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix Crosslinking Forces Tumor Progression by Enhancing Integrin Signaling. Cell 2009, 139, 891–906.
- Coste, B.; Xiao, B.; Santos, J.S.; Syeda, R.; Grandl, J.; Spencer, K.S.; Kim, S.E.; Schmidt, M.; Mathur, J.; Dubin, A.E.; et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 2012, 483, 176–181.
- Zanconato, F.; Cordenonsi, M.; Piccolo, S. YAP/TAZ at the Roots of Cancer. Cancer Cell 2016, 29, 783–803.
- Chin, L.K.; Xia, Y.; Discher, D.E.; Janmey, P.A. Mechanotransduction in cancer. Curr. Opin. Chem. Eng. 2016, 11, 77–84.
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54.
- Wei, B.; Zhou, X.; Liang, C.; Zheng, X.; Lei, P.; Fang, J.; Han, X.; Wang, L.; Qi, C.; Wei, H. Human colorectal cancer progression correlates with LOX-induced ECM stiffening. Int. J. Biol. Sci. 2017, 13, 1450–1457.
- Su, G.; Meyer, K.; Nandini, C.D.; Qiao, D.; Salamat, S.; Friedl, A. Glypican-1 is frequently overexpressed in human gliomas and enhances FGF-2 signaling in glioma cells. Am. J. Pathol. 2006, 168, 2014–2026.
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031.
- Bordeleau, F.; Mason, B.N.; Lollis, E.M.; Mazzola, M.; Zanotelli, M.R.; Somasegar, S.; Califano, J.P.; Montague, C.; LaValley, D.J.; Huynh, J.; et al. Matrix stiffening promotes a tumor vasculature phenotype. Proc. Natl. Acad. Sci. USA 2017, 114, 492–497.
- Mcmahon, M.; Ye, S.; Pedrina, J.; Dlugolenski, D.; Stambas, J. Extracellular Matrix Enzymes and Immune Cell Biology. Front. Mol. Biosci. 2021, 8, 703868.
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 ( MMP-9 ) and its inhibitors in cancer: A minireview. Eur. J. Med. Chem. 2020, 194, 112260.
- Kumar, S.; Das, A.; Barai, A.; Sen, S. MMP Secretion Rate and Inter-invadopodia Spacing Collectively Govern Cancer Invasiveness. Biophysj 2018, 114, 650–662.
- Caruana, I.; Savoldo, B.; Hoyos, V.; Weber, G.; Liu, H.; Kim, E.S.; Ittmann, M.M.; Marchetti, D.; Dotti, G. Heparanase promotes tumor infiltration and antitumor activity of CAR-redirected T lymphocytes. Nat. Med. 2015, 21, 524–529.
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzyme Inhib. Med. Chem. 2016, 31, 177–183.
- Renkawitz, J.; Kopf, A.; Stopp, J.; de Vries, I.; Driscoll, M.K.; Merrin, J.; Hauschild, R.; Welf, E.S.; Danuser, G.; Fiolka, R.; et al. Nuclear positioning facilitates amoeboid migration along the path of least resistance. Nature 2019, 568, 546–550.
- Krummel, M.F.; Friedman, R.S.; Jacobelli, J. Modes and mechanisms of T cell motility: Roles for confinement and Myosin-IIA. Curr. Opin. Cell Biol. 2014, 30, 9–16.
- Tabdanov, E.D.; Rodríguez-Merced, N.J.; Cartagena-Rivera, A.X.; Puram, V.V.; Callaway, M.K.; Ensminger, E.A.; Pomeroy, E.J.; Yamamoto, K.; Lahr, W.S.; Webber, B.R.; et al. Engineering T cells to enhance 3D migration through structurally and mechanically complex tumor microenvironments. Nat. Commun. 2021, 12, 1–17.
- Salmon, H.; Franciszkiewicz, K.; Damotte, D.; Dieu-Nosjean, M.C.; Validire, P.; Trautmann, A.; Mami-Chouaib, F.; Donnadieu, E. Matrix architecture defines the preferential localization and migration of T cells into the stroma of human lung tumors. J. Clin. Investig. 2012, 122, 899–910.
- Fanfone, D.; Wu, Z.; Mammi, J.; Berthenet, K.; Neves, D.; Weber, K.; Halaburkova, A.; Virard, F.; Bunel, F.; Jamard, C.; et al. Confined migration promotes cancer metastasis through resistance to anoikis and increased invasiveness. Elife 2022, 11, e73150.
- De Cicco, P.; Ercolano, G.; Ianaro, A. The New Era of Cancer Immunotherapy: Targeting Myeloid-Derived Suppressor Cells to Overcome Immune Evasion. Front. Immunol. 2020, 11, 1680.
- Rianna, C.; Radmacher, M.; Kumar, S.; Discher, D. Direct evidence that tumor cells soften when navigating confined spaces. Mol. Biol. Cell 2020, 31, 1726–1734.
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 2006, 124, 263–266.
- Bufi, N.; Saitakis, M.; Dogniaux, S.; Buschinger, O.; Bohineust, A.; Richert, A.; Maurin, M.; Hivroz, C.; Asnacios, A. Human primary immune cells exhibit distinct mechanical properties that are modified by inflammation. Biophys. J. 2015, 108, 2181–2190.
- Wullkopf, L.; West, A.K.V.; Leijnse, N.; Cox, T.R.; Madsen, C.D.; Oddershede, L.B.; Erler, J.T. Cancer cells’ ability to mechanically adjust to extracellular matrix stiffness correlates with their invasive potential. Mol. Biol. Cell 2018, 29, 2378–2385.
- Dhar, M.; Lam, J.N.; Walser, T.; Dubinett, S.M.; Rettig, M.B.; Di Carlo, D. Functional profiling of circulating tumor cells with an integrated vortex capture and single-cell protease activity assay. Proc. Natl. Acad. Sci. USA 2018, 115, 9986–9991.
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174.
- Xiang, H.; Ramil, C.P.; Hai, J.; Zhang, C.; Wang, H.; Watkins, A.A.; Afshar, R.; Georgiev, P.; Sze, M.A.; Song, X.S.; et al. Cancer-associated fibroblasts promote immunosuppression by inducing ROS-generating monocytic MDSCs in lung squamous cell carcinoma. Cancer Immunol. Res. 2020, 8, 436–450.
- Tamura, R.; Tanaka, T.; Akasaki, Y.; Murayama, Y.; Yoshida, K.; Sasaki, H. The role of vascular endothelial growth factor in the hypoxic and immunosuppressive tumor microenvironment: Perspectives for therapeutic implications. Med. Oncol. 2020, 37, 2.
- Tauriello, D.V.F.; Palomo-Ponce, S.; Stork, D.; Berenguer-Llergo, A.; Badia-Ramentol, J.; Iglesias, M.; Sevillano, M.; Ibiza, S.; Cañellas, A.; Hernando-Momblona, X.; et al. TGFβ drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 2018, 554, 538–543.
- Deleon-pennell, K.Y.; Barker, T.H.; Lindsey, M.L. Fibroblasts: The arbiters of extracellular matrix remodeling. Matrix Biol. 2020, 91–92, 1–7.
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136.
- Gubin, M.M.; Zhang, X.; Schuster, H.; Caron, E.; Ward, J.P.; Noguchi, T.; Ivanova, Y.; Hundal, J.; Arthur, C.D.; Krebber, W.J.; et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 2014, 515, 577–581.
- Jang, I.; Beningo, K.A. Integrins, CAFs and mechanical forces in the progression of cancer. Cancers 2019, 11, 721.
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell 2005, 8, 241–254.
- Kim, J.; Feng, J.; Jones, C.A.R.; Mao, X.; Sander, L.M.; Levine, H.; Sun, B. Stress-induced plasticity of dynamic collagen networks. Nat. Commun. 2017, 8.
- Swift, J.; Discher, D.E. The nuclear lamina is mechano-responsive to ECM elasticity in mature tissue. J. Cell Sci. 2014, 127, 3005–3015.
- Northcott, J.M.; Dean, I.S.; Mouw, J.K.; Weaver, V.M. Feeling Stress: The Mechanics of Cancer Progression and Aggression. Front. Cell Dev. Biol. 2018, 6, 17.
- Stylianopoulos, T.; Martin, J.D.; Snuderl, M.; Mpekris, F.; Jain, S.R.; Jain, R.K. Co-evolution of solid stress and interstitial fluid pressure in tumors during progression: Implications for vascular collapse. Cancer Res. 2013, 73, 3833–3841.
- Goetz, J.G.; Minguet, S.; Navarro-Lérida, I.; Lazcano, J.J.; Samaniego, R.; Calvo, E.; Tello, M.; Osteso-Ibáñez, T.; Pellinen, T.; Echarri, A.; et al. Biomechanical remodeling of the microenvironment by stromal caveolin-1 favors tumor invasion and metastasis. Cell 2011, 146, 148–163.
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770.
- Zuazo-gaztelu, I.; Casanovas, O. Unraveling the Role of Angiogenesis in Cancer ecosystems. Front. Oncol. 2018, 8, 248.
- Forster, J.C.; Harriss-phillips, W.M.; Douglass, M.J.J.; Bezak, E. A review of the development of tumor vasculature and its effects on the tumor microenvironment. Hypoxia 2017, 5, 21–32.
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 2, 213–219.
- Cheng, G.; Tse, J.; Jain, R.K.; Munn, L.L. Micro-environmental mechanical stress controls tumor spheroid size and morphology by suppressing proliferation and inducing apoptosis in cancer cells. PLoS ONE 2009, 4, e4632.
- Nia, H.T.; Liu, H.; Seano, G.; Datta, M.; Jones, D.; Rahbari, N.; Incio, J.; Chauhan, V.P.; Jung, K.; Martin, J.D.; et al. Solid stress and elastic energy as measures of tumour mechanopathology. Nat. Biomed. Eng. 2016, 1, 0004.
- Hofmann, M.; Guschel, M.; Bernd, A.; Bereiter-Hahn, J.; Kaufmann, R.; Tandi, C.; Helge, W.; Kippenberger, S. Lowering of tumor interstitial fluid pressure reduces tumor cell proliferation in a xenograft tumor model. Neoplasia 2006, 8, 89–95.
- Rofstad, E.K.; Galappathi, K.; Mathiesen, B.S. Tumor Interstitial Fluid Pressure-A Link between Tumor Hypoxia, Microvascular Density, and Lymph Node Metastasis. Neoplasia 2014, 16, 586–594.
- Rens, E.G.; Merks, R.M.H. Cell Contractility Facilitates Alignment of Cells and Tissues to Static Uniaxial Stretch. Biophys. J. 2017, 112, 755–766.
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 2006, 4, 1–15.
- Kuczek, D.E.; Larsen, A.M.H.; Thorseth, M.L.; Carretta, M.; Kalvisa, A.; Siersbæk, M.S.; Simões, A.M.C.; Roslind, A.; Engelholm, L.H.; Noessner, E.; et al. Collagen density regulates the activity of tumor-infiltrating T cells. J. Immunother. Cancer 2019, 7, 68.
- Riching, K.M.; Cox, B.L.; Salick, M.R.; Pehlke, C.; Riching, A.S.; Ponik, S.M.; Bass, B.R.; Crone, W.C.; Jiang, Y.; Weaver, A.M.; et al. 3D collagen alignment limits protrusions to enhance breast cancer cell persistence. Biophys. J. 2015, 107, 2546–2558.
- Gocheva, V.; Wang, H.W.; Gadea, B.B.; Shree, T.; Hunter, K.E.; Garfall, A.L.; Berman, T.; Joyce, J.A. IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev. 2010, 24, 241–255.
- Erdogan, B.; Ao, M.; White, L.M.; Means, A.L.; Brewer, B.M.; Yang, L.; Washington, M.K.; Shi, C.; Franco, O.E.; Weaver, A.M.; et al. Cancer-associated fibroblasts promote directional cancer cell migration by aligning fibronectin. J. Cell Biol. 2017, 216, 3799–3816.
- Koorman, T.; Jansen, K.A.; Khalil, A.; Haughton, P.D.; Visser, D.; Rätze, M.A.K.; Haakma, W.E.; Sakalauskaitè, G.; van Diest, P.J.; de Rooij, J.; et al. Spatial collagen stiffening promotes collective breast cancer cell invasion by reinforcing extracellular matrix alignment. Oncogene 2022.
- Jain, R.K. Delivery of molecular and cellular medicine to solid tumors. Adv. Drug Deliv. Rev. 2012, 64, 353–365.
- Heldin, C.H.; Rubin, K.; Pietras, K.; Östman, A. High interstitial fluid pressure—An obstacle in cancer therapy. Nat. Rev. Cancer 2004, 4, 806–813.
- Provenzano, P.P.; Hingorani, S.R. Hyaluronan, fluid pressure, and stromal resistance in pancreas cancer. Br. J. Cancer 2013, 108, 1–8.
- Libutti, S.K.; Tamarkin, L.; Nilubol, N. Targeting the invincible barrier for drug delivery in solid cancers: Interstitial fluid pressure. Oncotarget 2018, 9, 35723–35725.
- Chen, S.C.; Wu, P.C.; Wang, C.Y.; Kuo, P.L. Evaluation of cytotoxic T lymphocyte-mediated anticancer response against tumor interstitium-simulating physical barriers. Sci. Rep. 2020, 10, 13662.
- Helmlinger, G.; Netti, P.A.; Lichtenbeld, H.C.; Melder, R.J.; Jain, R.K. Solid stress inhibits the growth of multicellular tumor spheroids. Nat. Biotechnol. 1997, 15, 778–783.
- Chen, A.; Sceneay, J.; Gödde, N.; Kinwel, T.; Ham, S.; Thompson, E.W.; Humbert, P.O.; Möller, A. Intermittent hypoxia induces a metastatic phenotype in breast cancer. Oncogene 2018, 37, 4214–4225.
- Lewis, C.; Murdoch, C. Macrophage responses to hypoxia: Implications for tumor progression and anti-cancer therapies. Am. J. Pathol. 2005, 167, 627–635.
- Winkler, J.; Werb, Z.; Metcalf, K.J. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120.
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFβ in the tumour microenvironment. Nat. Rev. Cancer 2013, 13, 788–799.
- Martinez, M.; Moon, E.K. CAR T cells for solid tumors: New strategies for finding, infiltrating, and surviving in the tumor microenvironment. Front. Immunol. 2019, 10, 1–21.
- Gargalionis, A.N.; Basdra, E.K.; Papavassiliou, A.G. Mechanosignalling in tumour progression. J. Cell. Mol. Med. 2018, 22, 704–705.
- Chaudhuri, O.; Koshy, S.T.; Branco Da Cunha, C.; Shin, J.W.; Verbeke, C.S.; Allison, K.H.; Mooney, D.J. Extracellular matrix stiffness and composition jointly regulate the induction of malignant phenotypes in mammary epithelium. Nat. Mater. 2014, 13, 970–978.
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406.
- Crotti, S.; Piccoli, M.; Rizzolio, F.; Giordano, A.; Nitti, D.; Agostini, M. Extracellular Matrix and Colorectal Cancer: How Surrounding Microenvironment Affects Cancer Cell Behavior? J. Cell. Physiol. 2017, 232, 967–975.
- Moriggi, M.; Giussani, M.; Torretta, E.; Capitanio, D.; Sandri, M.; Leone, R.; De Palma, S.; Vasso, M.; Vozzi, G.; Tagliabue, E.; et al. ECM Remodeling in Breast Cancer with Different Grade: Contribution of 2D-DIGE Proteomics. Proteomics 2018, 18, e1800278.
- Ingber, D.E. Mechanical signaling and the cellular response to extracellular matrix in angiogenesis and cardiovascular physiology. Circ. Res. 2002, 91, 877–887.
- Tuxhorn, J.A.; Mcalhany, S.J.; Dang, T.D.; Ayala, G.E.; Rowley, D.R. Stromal Cells Promote Angiogenesis and Growth of Human Prostate Tumors in a Differential Reactive Stroma ( DRS ) Xenograft Model 1. Cancer Res. 2002, 62, 3298–3307.
- Gupta, M.K.; Qin, R. Mechanism and its regulation of tumor-induced angiogenesis. World J. Gastroenterol. WJG 2003, 9, 1144–1155.
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009.
- Kim, J.; Zheng, Y.; Alobaidi, A.A.; Nan, H.; Tian, J.; Jiao, Y.; Sun, B. Collective ECM remodeling organizes 3D collective cancer invasion. arXiv 2019, arXiv:1903.03290.
- Chaudhuri, P.K.; Low, B.C.; Lim, C.T. Mechanobiology of Tumor Growth. Chem. Rev. 2018, 118, 6499–6515.
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular Matrix in the Tumor Microenvironment and Its Impact on Cancer Therapy. Front. Mol. Biosci. 2020, 6, 160.
- Huang, Y.; Yuan, J.; Righi, E.; Kamoun, W.S.; Ancukiewicz, M.; Nezivar, J.; Santosuosso, M.; Martin, J.D.; Martin, M.R.; Vianello, F.; et al. Vascular normalizing doses of antiangiogenic treatment reprogram the immunosuppressive tumor microenvironment and enhance immunotherapy. Proc. Natl. Acad. Sci. USA 2012, 109, 17561–17566.
- Whatcott, C.J.; Han, H.; Posner, R.G.; Hostetter, G.; Von Hoff, D.D. Targeting the tumor microenvironment in cancer: Why hyaluronidase deserves a second look. Cancer Discov. 2011, 1, 291–296.
- Venning, F.A.; Wullkopf, L.; Erler, J.T. Targeting ECM disrupts cancer progression. Front. Oncol. 2015, 5, 224.
- Barry-Hamilton, V.; Spangler, R.; Marshall, D.; McCauley, S.; Rodriguez, H.M.; Oyasu, M.; Mikels, A.; Vaysberg, M.; Ghermazien, H.; Wai, C.; et al. Allosteric inhibition of lysyl oxidase-like-2 impedes the development of a pathologic microenvironment. Nat. Med. 2010, 16, 1009–1017.
- Lanitis, E.; Irving, M.; Coukos, G. Targeting the tumor vasculature to enhance T cell activity. Curr. Opin. Immunol. 2015, 33, 55–63.
- Feig, C.; Jones, J.O.; Kraman, M.; Wells, R.J.B.; Deonarine, A.; Chan, D.S.; Connell, C.M.; Roberts, E.W.; Zhao, Q.; Caballero, O.L.; et al. Targeting CXCL12 from FAP-expressing carcinoma-associated fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 20212–20217.
- Zboralski, D.; Hoehlig, K.; Eulberg, D.; Frömming, A.; Vater, A. Increasing tumor-infiltrating T cells through inhibition of CXCL12 with NOX-A12 synergizes with PD-1 blockade. Cancer Immunol. Res. 2017, 5, 950–956.
- Mariathasan, S.; Turley, S.J.; Nickles, D.; Castiglioni, A.; Yuen, K.; Wang, Y.; Kadel, E.E.; Koeppen, H.; Astarita, J.L.; Cubas, R.; et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 2018, 554, 544–548.
- Ito, T.; Hanabuchi, S.; Wang, Y.H.; Park, W.R.; Arima, K.; Bover, L.; Qin, F.X.F.; Gilliet, M.; Liu, Y.J. Two Functional Subsets of FOXP3+ Regulatory T Cells in Human Thymus and Periphery. Immunity 2008, 28, 870–880.
- Hombach, A.A.; Heiders, J.; Foppe, M.; Chmielewski, M.; Abken, H. OX40 costimulation by a chimeric antigen receptor abrogates CD28 and IL-2 induced IL-10 secretion by redirected CD4+ T cells. Oncoimmunology 2012, 1, 458–466.
- Sato, T.; Terai, M.; Tamura, Y.; Alexeev, V.; Mastrangelo, M.J.; Selvan, S.R. Interleukin 10 in the tumor microenvironment: A target for anticancer immunotherapy. Immunol. Res. 2011, 51, 170–182.
- Wang, Y.; Jiang, H.; Luo, H.; Sun, Y.; Shi, B.; Sun, R.; Li, Z. An IL-4/21 Inverted Cytokine Receptor Improving CAR-T Cell Potency in Immunosuppressive Solid-Tumor Microenvironment. Front. Immunol. 2019, 10, 1691.
- Liu, K.; Huang, A.; Nie, J.; Tan, J.; Xing, S.; Qu, Y.; Jiang, K. IL-35 Regulates the Function of Immune Cells in Tumor Microenvironment. Front. Immunol. 2021, 12, 683332.
- Heim, L.; Kachler, K.; Siegmund, R.; Trufa, D.I.; Mittler, S.; Geppert, C.I.; Friedrich, J.; Rieker, R.J.; Sirbu, H.; Finotto, S. Increased expression of the immunosuppressive interleukin-35 in patients with non-small cell lung cancer. Br. J. Cancer 2019, 120, 903–912.
- Morrot, A.; da Fonseca, L.M.; Salustiano, E.J.; Gentile, L.B.; Conde, L.; Filardy, A.A.; Franklim, T.N.; da Costa, K.M.; Freire-de-Lima, C.G.; Freire-de-Lima, L. Metabolic symbiosis and immunomodulation: How tumor cell-derived lactate may disturb innate and adaptive immune responses. Front. Oncol. 2018, 8, 81.
- Vigano, S.; Alatzoglou, D.; Irving, M.; Ménétrier-Caux, C.; Caux, C.; Romero, P.; Coukos, G. Targeting adenosine in cancer immunotherapy to enhance T-Cell function. Front. Immunol. 2019, 10, 925.
- Moon, E.K.; Wang, L.C.S.; Bekdache, K.; Lynn, R.C.; Lo, A.; Thorne, S.H.; Albelda, S.M. Intra-tumoral delivery of CXCL11 via a vaccinia virus, but not by modified T cells, enhances the efficacy of adoptive T cell therapy and vaccines. Oncoimmunology 2018, 7, e1395997.
- Nishio, N.; Diaconu, I.; Liu, H.; Cerullo, V.; Caruana, I.; Hoyos, V.; Bouchier-Hayes, L.; Savoldo, B.; Dotti, G. Armed oncolytic virus enhances immune functions of chimeric antigen receptor-modified T cells in solid tumors. Cancer Res. 2014, 74, 5195–5205.
- McGrath, K.; Dotti, G. Combining oncolytic viruses with chimeric antigen receptor T cell therapy. Hum. Gene Ther. 2021, 32, 150–157.
- Koneru, M.; Purdon, T.J.; Spriggs, D.; Koneru, S.; Brentjens, R.J. IL-12 secreting tumor-targeted chimeric antigen receptor T cells eradicate ovarian tumors in vivo. Oncoimmunology 2015, 4, e994446.
- Hu, B.; Ren, J.; Luo, Y.; Keith, B.; Young, R.M.; Scholler, J.; Zhao, Y.; June, C.H. Augmentation of Antitumor Immunity by Human and Mouse CAR T Cells Secreting IL-18. Cell Rep. 2017, 20, 3025–3033.
- Shum, T.; Omer, B.; Tashiro, H.; Kruse, R.L.; Wagner, D.L.; Parikh, K.; Yi, Z.; Sauer, T.; Liu, D.; Parihar, R.; et al. Constitutive signaling from an engineered IL7 receptor promotes durable tumor elimination by tumor-redirected T cells. Cancer Discov. 2017, 7, 1238–1247.
- Adachi, K.; Kano, Y.; Nagai, T.; Okuyama, N.; Sakoda, Y.; Tamada, K. IL-7 and CCL19 expression in CAR-T cells improves immune cell infiltration and CAR-T cell survival in the tumor. Nat. Biotechnol. 2018, 36, 346–351.
- Kershaw, M.H.; Wang, G.; Westwood, J.A.; Pachynski, R.K.; Tiffany, H.L.; Marincola, F.M.; Wang, E.; Young, H.A.; Murphy, P.M.; Hwu, P. Redirecting migration of T cells to chemokine secreted from tumors by genetic modification with CXCR2. Hum. Gene Ther. 2002, 13, 1971–1980.
- Craddock, J.A.; Lu, A.; Bear, A.; Pule, M.; Brenner, M.K.; Rooney, C.M.; Foster, A.E. Enhanced tumor trafficking of GD2 chimeric antigen receptor T cells by expression of the chemokine receptor CCR2b. J. Immunother. 2010, 33, 780–788.
- Moon, E.K.; Carpenito, C.; Sun, J.; Wang, L.C.S.; Kapoor, V.; Predina, J.; Powell, D.J.; Riley, J.L.; June, C.H.; Albelda, S.M. Expression of a functional CCR2 receptor enhances tumor localization and tumor eradication by retargeted human T cells expressing a mesothelin-specific chimeric antibody receptor. Clin. Cancer Res. 2011, 17, 4719–4730.
- Jin, L.; Tao, H.; Karachi, A.; Long, Y.; Hou, A.Y.; Na, M.; Dyson, K.A.; Grippin, A.J.; Deleyrolle, L.P.; Zhang, W.; et al. CXCR1- or CXCR2-modi fi ed CAR T cells co-opt IL- 8 for maximal antitumor ef fi cacy in solid tumors. Nat. Commun. 2019, 10, 4016.
- Jin, L.; Ge, H.; Long, Y.; Yang, C.; Chang, Y.E.; Mu, L.; Sayour, E.J.; De Leon, G.; Wang, Q.J.; Yang, J.C.; et al. CD70, a novel target of CAR T-cell therapy for gliomas. Neuro. Oncol. 2018, 20, 55–65.
- Zhang, L.; Yu, Z.; Muranski, P.; Palmer, D.C.; Restifo, N.P.; Rosenberg, S.A.; Morgan, R.A. Inhibition of TGF-β signaling in genetically engineered tumor antigen-reactive T cells significantly enhances tumor treatment efficacy. Gene Ther. 2013, 20, 575–580.
- Chen, N.; Morello, A.; Tano, Z.; Adusumilli, P.S. CAR T-cell intrinsic PD-1 checkpoint blockade: A two-in-one approach for solid tumor immunotherapy. Oncoimmunology 2017, 6, e1273302.
- Mohammed, S.; Sukumaran, S.; Bajgain, P.; Watanabe, N.; Heslop, H.E.; Rooney, C.M.; Brenner, M.K.; Fisher, W.E.; Leen, A.M.; Vera, J.F. Improving Chimeric Antigen Receptor-Modified T Cell Function by Reversing the Immunosuppressive Tumor Microenvironment of Pancreatic Cancer. Mol. Ther. 2017, 25, 249–258.
- Bajgain, P.; Tawinwung, S.; D’Elia, L.; Sukumaran, S.; Watanabe, N.; Hoyos, V.; Lulla, P.; Brenner, M.K.; Leen, A.M.; Vera, J.F. CAR T cell therapy for breast cancer: Harnessing the tumor milieu to drive T cell activation. J. Immunother. Cancer 2018, 6, 1–13.
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 86.
- Liu, T.; Zhou, L.; Li, D.; Andl, T.; Zhang, Y. Cancer-associated fibroblasts build and secure the tumor microenvironment. Front. Cell Dev. Biol. 2019, 7, 60.
- Harryvan, T.J.; Verdegaal, E.M.E.; Hardwick, J.C.H.; Hawinkels, L.J.A.C.; van der Burg, S.H. Targeting of the cancer-associated fibroblast—t-cell axis in solid malignancies. J. Clin. Med. 2019, 8, 1989.
- Lakins, M.A.; Ghorani, E.; Munir, H.; Martins, C.P.; Shields, J.D. Cancer-associated fibroblasts induce antigen-specific deletion of CD8 + T Cells to protect tumour cells. Nat. Commun. 2018, 9, 948.
- Lo, A.; Wang, L.C.S.; Scholler, J.; Monslow, J.; Avery, D.; Newick, K.; O’Brien, S.; Evans, R.A.; Bajor, D.J.; Clendenin, C.; et al. Tumor-promoting desmoplasia is disrupted by depleting FAP-expressing stromal cells. Cancer Res. 2015, 75, 2800–2810.
- Kakarla, S.; Chow, K.K.H.; Mata, M.; Shaffer, D.R.; Song, X.T.; Wu, M.F.; Liu, H.; Wang, L.L.; Rowley, D.R.; Pfizenmaier, K.; et al. Antitumor effects of chimeric receptor engineered human T cells directed to tumor stroma. Mol. Ther. 2013, 21, 1611–1620.
- Özdemir, B.C.; Pentcheva-Hoang, T.; Carstens, J.L.; Zheng, X.; Wu, C.C.; Simpson, T.R.; Laklai, H.; Sugimoto, H.; Kahlert, C.; Novitskiy, S.V.; et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell 2014, 25, 719–734.
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284.
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455.
- Law, A.M.K.; Valdes-mora, F.; Gallego-ortega, D. Myeloid-Derived Suppressor Cells as a Therapeutic. Cells 2020, 27, 561.
- Pollard, J.W. Trophic macrophages in development and disease. Nat. Rev. Immunol. 2009, 9, 259–270.
- Qian, B.-Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51.
- Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Bronte, V. Coordinated regulation of myeloid cells by tumours. Nat. Rev. Immunol. 2012, 12, 253–268.
- Sica, A.; Saccani, A.; Bottazzi, B.; Polentarutti, N.; Vecchi, A.; Van Damme, J.; Mantovani, A. Autocrine Production of IL-10 Mediates Defective IL-12 Production and NF-κB Activation in Tumor-Associated Macrophages. J. Immunol. 2000, 164, 762–767.
- Rodriguez-Garcia, A.; Lynn, R.C.; Poussin, M.; Eiva, M.A.; Shaw, L.C.; O’Connor, R.S.; Minutolo, N.G.; Casado-Medrano, V.; Lopez, G.; Matsuyama, T.; et al. CAR-T cell-mediated depletion of immunosuppressive tumor-associated macrophages promotes endogenous antitumor immunity and augments adoptive immunotherapy. Nat. Commun. 2021, 12, 877.
- Wang, G.; Lu, X.; Dey, P.; Deng, P.; Wu, C.C.; Jiang, S.; Fang, Z.; Zhao, K.; Konaparthi, R.; Hua, S.; et al. Targeting YAP-dependent MDSC infiltration impairs tumor progression. Cancer Discov. 2016, 6, 80–95.
- Alban, T.J.; Bayik, D.; Otvos, B.; Rabljenovic, A.; Leng, L.; Jia-Shiun, L.; Roversi, G.; Lauko, A.; Momin, A.A.; Mohammadi, A.M.; et al. Glioblastoma Myeloid-Derived Suppressor Cell Subsets Express Differential Macrophage Migration Inhibitory Factor Receptor Profiles That Can Be Targeted to Reduce Immune Suppression. Front. Immunol. 2020, 11, 1191.
- Nalawade, S.A.; Shafer, P.; Bajgain, P.; McKenna, M.K.; Ali, A.; Kelly, L.; Joubert, J.; Gottschalk, S.; Watanabe, N.; Leen, A.; et al. Selectively targeting myeloid-derived suppressor cells through TRAIL receptor 2 to enhance the efficacy of CAR T cell therapy for treatment of breast cancer. J. Immunother. Cancer 2021, 9, e003237.
- Shaul, M.E.; Fridlender, Z.G. Tumour-associated neutrophils in patients with cancer. Nat. Rev. Clin. Oncol. 2019, 16, 601–620.
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression—Implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371.
- Amoozgar, Z.; Kloepper, J.; Ren, J.; Tay, R.E.; Kazer, S.W.; Kiner, E.; Krishnan, S.; Posada, J.M.; Ghosh, M.; Mamessier, E.; et al. Targeting Treg cells with GITR activation alleviates resistance to immunotherapy in murine glioblastomas. Nat. Commun. 2021, 12, 2582.
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264.
- Reinhard, K.; Rengstl, B.; Oehm, P.; Michel, K.; Billmeier, A.; Hayduk, N.; Klein, O.; Kuna, K.; Ouchan, Y.; Wöll, S.; et al. An RNA vaccine drives expansion and efficacy of claudin-CAR-T cells against solid tumors. Science 2020, 367, 446–453.
- Sadjadi, Z.; Zhao, R.; Hoth, M.; Qu, B.; Rieger, H. Migration of Cytotoxic T Lymphocytes in 3D Collagen Matrices. Biophys. J. 2020, 119, 2141–2152.
- Wolf, K.; Müller, R.; Borgmann, S.; Bröcker, E.B.; Friedl, P. Amoeboid shape change and contact guidance: T-lymphocyte crawling through fibrillar collagen is independent of matrix remodeling by MMPs and other proteases. Blood 2003, 102, 3262–3269.
- Jacobelli, J.; Friedman, R.S.; Conti, M.A.; Lennon-Dumenil, A.M.; Piel, M.; Sorensen, C.M.; Adelstein, R.S.; Krummel, M.F. Confinement-optimized three-dimensional T cell amoeboid motility is modulated via myosin IIA-regulated adhesions. Nat. Immunol. 2010, 11, 953–961.
- Kawauchi, T. Cell adhesion and its endocytic regulation in cell migration during neural development and cancer metastasis. Int. J. Mol. Sci. 2012, 13, 4564–4590.
- Petrie, R.J.; Harlin, H.M.; Korsak, L.I.T.; Yamada, K.M. Activating the nuclear piston mechanism of 3D migration in tumor cells. J. Cell Biol. 2017, 216, 93–100.
- Ruprecht, V.; Wieser, S.; Callan-Jones, A.; Smutny, M.; Morita, H.; Sako, K.; Barone, V.; Ritsch-Marte, M.; Sixt, M.; Voituriez, R.; et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell 2015, 160, 673–685.
- Yamada, K.M.; Sixt, M. Mechanisms of 3D cell migration. Nat. Rev. Mol. Cell Biol. 2019, 20, 738–752.
- Miller, M.J.; Wei, S.H.; Parker, I.; Cahalan, M.D. Two-photon imaging of lymphocyte motility and antigen response in intact lymph node. Science 2002, 296, 1869–1873.
- Friedl, P.; Entschladen, F.; Conrad, C.; Niggemann, B.; Zänker, K.S. CD4+ T lymphocytes migrating in three-dimensional collagen lattices lack focal adhesions and utilize β1 integrin-independent strategies for polarization, interaction with collagen fibers and locomotion. Eur. J. Immunol. 1998, 28, 2331–2343.
- Paluch, E.K.; Aspalter, I.M.; Sixt, M. Focal Adhesion-Independent Cell Migration. Annu. Rev. Cell Dev. Biol. 2016, 32, 469–490.
- Lämmermann, T.; Bader, B.L.; Monkley, S.J.; Worbs, T.; Wedlich-Söldner, R.; Hirsch, K.; Keller, M.; Förster, R.; Critchley, D.R.; Fässler, R.; et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 2008, 453, 51–55.
- Goudarzi, M.; Banisch, T.U.; Mobin, M.B.; Maghelli, N.; Tarbashevich, K.; Strate, I.; van den Berg, J.; Blaser, H.; Bandemer, S.; Paluch, E.; et al. Identification and Regulation of a Molecular Module for Bleb-Based Cell Motility. Dev. Cell 2012, 23, 210–218.
- Wu, S.K.; Priya, R. Spatio-temporal regulation of RhoGTPases signaling by myosin II. Front. Cell Dev. Biol. 2019, 7, 90.
- Wolf, K.; te Lindert, M.; Krause, M.; Alexander, S.; te Riet, J.; Willis, A.L.; Hoffman, R.M.; Figdor, C.G.; Weiss, S.J.; Friedl, P. Physical limits of cell migration: Control by ECM space and nuclear deformation and tuning by proteolysis and traction force. J. Cell Biol. 2013, 201, 1069–1084.
- Lele, T.P.; Dickinson, R.B.; Gundersen, G.G. Mechanical principles of nuclear shaping and positioning. J. Cell Biol. 2018, 217, 3330–3342.
- Xia, Y.; Pfeifer, C.R.; Discher, D.E. Nuclear mechanics during and after constricted migration. Acta Mech. Sin. Xuebao 2019, 35, 299–308.
- Swift, J.; Ivanovska, I.L.; Buxboim, A.; Harada, T.; Dingal, P.C.D.P.; Pinter, J.; Pajerowski, J.D.; Spinler, K.R.; Shin, J.W.; Tewari, M.; et al. Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation. Science 2013, 341, 1240104.
- Harada, T.; Swift, J.; Irianto, J.; Shin, J.W.; Spinler, K.R.; Athirasala, A.; Diegmiller, R.; Dingal, P.C.D.P.; Ivanovska, I.L.; Discher, D.E. Nuclear lamin stiffness is a barrier to 3D migration, but softness can limit survival. J. Cell Biol. 2014, 204, 669–682.
- Stecklein, S.R.; Kumaraswamy, E.; Behbod, F.; Wang, W.; Chaguturu, V.; Harlan-Williams, L.M.; Jensen, R.A. BRCA1 and HSP90 cooperate in homologous and non-homologous DNA double-strand-break repair and G2/M checkpoint activation. Proc. Natl. Acad. Sci. USA 2012, 109, 13650–13655.
- Stephens, A.D.; Banigan, E.J.; Adam, S.A.; Goldman, R.D.; Marko, J.F. Chromatin and lamin A determine two different mechanical response regimes of the cell nucleus. Mol. Biol. Cell 2017, 28, 1984–1996.
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183.
- Lambert, L.H.; Goebrecht, G.K.E.; De Leo, S.E.; O’Connor, R.S.; Nunez-Cruz, S.; De Li, T.; Yuan, J.; Milone, M.C.; Kam, L.C. Improving T Cell Expansion with a Soft Touch. Nano Lett. 2017, 17, 821–826.
- Bashoura, K.T.; Gondarenko, A.; Chen, H.; Shen, K.; Liu, X.; Huse, M.; Hone, J.C.; Kam, L.C. CD28 and CD3 have complementary roles in T-cell traction forces. Proc. Natl. Acad. Sci. USA 2014, 111, 2241–2246.
- Kim, S.T.; Takeuchi, K.; Sun, Z.Y.J.; Touma, M.; Castro, C.E.; Fahmy, A.; Lang, M.J.; Wagner, G.; Reinherz, E.L. The αβ T cell receptor is an anisotropic mechanosensor. J. Biol. Chem. 2009, 284, 31028–31037.
- Basu, R.; Whitlock, B.M.; Husson, J.; Le Floc’h, A.; Jin, W.; Oyler-Yaniv, A.; Dotiwala, F.; Giannone, G.; Hivroz, C.; Biais, N.; et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell 2016, 165, 100–110.
- Buxboim, A.; Swift, J.; Irianto, J.; Spinler, K.R.; Dingal, P.C.D.P.; Athirasala, A.; Kao, Y.R.C.; Cho, S.; Harada, T.; Shin, J.W.; et al. Matrix elasticity regulates lamin-A,C phosphorylation and turnover with feedback to actomyosin. Curr. Biol. 2014, 24, 1909–1917.
- Judokusumo, E.; Tabdanov, E.; Kumari, S.; Dustin, M.L.; Kam, L.C. Mechanosensing in T lymphocyte activation. Biophys. J. 2012, 102, L5–L7.
- Jin, W.; Tamzalit, F.; Chaudhuri, P.K.; Black, C.T.; Huse, M.; Kam, L.C. T cell activation and immune synapse organization respond to the microscale mechanics of structured surfaces. Proc. Natl. Acad. Sci. USA 2019, 116, 19835–19840.
- Pruitt, H.C.; Lewis, D.; Ciccaglione, M.; Connor, S.; Smith, Q.; Hickey, J.W.; Schneck, J.P.; Gerecht, S. Collagen fiber structure guides 3D motility of cytotoxic T lymphocytes. Matrix Biol. 2020, 85–86, 147–159.
More
Information
Subjects:
Oncology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.3K
Entry Collection:
Biopharmaceuticals Technology
Revisions:
2 times
(View History)
Update Date:
05 Jul 2022
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No