 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tomer Avidor-Reiss | + 746 word(s) | 746 | 2020-09-28 05:03:45 | | | |
| 2 | Tomer Avidor-Reiss | Meta information modification | 746 | 2020-10-01 12:43:28 | | | | |
| 3 | Bruce Ren | Meta information modification | 746 | 2020-10-09 03:07:48 | | | | |
| 4 | Rui Liu | + 47 word(s) | 793 | 2020-10-15 03:34:07 | | |
Video Upload Options
The centrosome, which consists of two centrioles surrounded by pericentriolar material, is a unique structure that has retained its main features in organisms of various taxonomic groups from unicellular algae to mammals over one billion years of evolution. In addition to the most noticeable function of organizing the microtubule system in mitosis and interphase, the centrosome performs many other cell functions. In particular, centrioles are the basis for the formation of sensory primary cilia and motile cilia and flagella. Another principal function of centrosomes is the concentration in one place of regulatory proteins responsible for the cell's progression along the cell cycle. Despite the existing exceptions, the functioning of the centrosome is subject to general principles.
1. Introduction
Nearly 150 years ago, almost simultaneously, three researchers described in dividing cells two symmetrically located structures that looked like a “radiance” and were called the centrosphere [1][2][3] (Fig 1). At the centrosphere's focus, granules were sometimes visible, which were originally called “polar corpuscles” [3]. Van Beneden and Nate [4] and independently Boveri [5] found that the polar corpuscles do not entirely disappear after mitosis, but remain in interphase, often located near the geometric center of the cell.
Later, these granules were named centrioles [6]. Shortly after, Henneguy and von Lenhossék showed that centrioles and basal bodies are the same structure at distinct functional stages. They proposed a “hypothesis about the homology of centrioles and basal bodies of flagella” [7][8]. This idea was overlooked for many years but ultimately is correct. After the ultrastructure of the centrosome was studied using transmission electron microscopy, centrioles (and basal bodies) were shown to have a conserved structure in many types of cells—hollow cylinders containing triplets of microtubules (MTs) in their walls [9][10][11]. The unique centrally symmetric structure of centrioles has always generated amazing, and sometimes fantastic, hypotheses about the centrosome's origin and functions. Fundamental advances in molecular biology, immunocytochemistry, and high-resolution light microscopy have allowed us to advance the understanding of the fundamental functions of this organelle that is small in size, but most important for many functions in the cells.
Summing up the many years of centrosome research, we formulated 12 postulates of centrosomal biology in 2007 [12]. A few years later, these postulates were somewhat supplemented and developed in a book on the centrosome [13]. The discoveries of recent years make it necessary once again to return to these postulates and update them.
2. Postulates
Postulate 1: The Centriole and the Basal Body Are Two Forms of the Same Organelle [7][8].
Postulate 2: The Centriole Is a Polar Structure with Two Morphologically and Functionally Different Ends.
Postulate 3: The Outer Diameter of the Distal Part of the Centriolar Cylinder Is Smaller Than That of the Proximal End.
Postulate 4: The Centriolar Cylinder Length Is Highly Regulated.
Postulate 5: In the Centrosome of Proliferating Cells, Two Centrioles Differ Structurally and Functionally.
Postulate 6: The Proximal Ends of the Mother and Daughter Centrioles Are Connected via a Bundle of Thin Fibers.
Postulate 7: The Centriole Lumen Helps in Stabilizing the Centriole .
Postulate 8: Maintaining a Cylindrical Shape of Centrioles Can Be Independent of MT Triplets
Postulate 9: The Structure and Activity of Centrosomes Differ in Interphase and Mitotic Cells .
Postulate 10: New Centrioles Are Usually Formed in Association with Mother Centrioles but Can Be Formed without Preexisting Centriole De Novo .
Postulate 11: The Centrosomes Have Four Types of MT Nucleating Activity.
Postulate 12: The Complete Process of Centrioles Maturation from Procentriole to Mother Centriole Takes More Than One and a Half Cell Cycles in Duration .
Postulate 13: The Centrosome Is the Center of the Organization of Actin Microfilaments in the Cell .
Postulate 14: Centrioles Can Gain an Atypical Structure and Composition and Become Undetected Using Standard Expectations and Techniques .
Postulate 15: The centrosome is a multiprotein and multifunctional cell organelle that includes complexes of cell cycle regulation, cilia and flagella formation, cytokinesis regulation, centriole duplication and separation, as well as microtubule nucleation and anchoring. Some centrosomal proteins are simultaneously involved in the regulation of several intracellular processes [14].
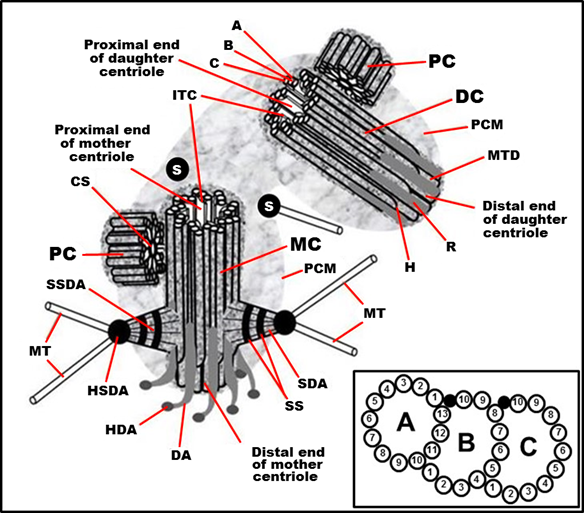
Figure 1. Typical centrosome structure in the S phase of the cell cycle in proliferating mammalian cells. MC: mother (mature) centriole; DC: daughter centriole; PC: procentriole; PCM: pericentriolar material (pericentriolar matrix); A: “A” MT of triplet; B: “B” MT of triplet; C: “C” MT of triplet; H: hook of “C” MT; MTD: A-B MT duplex (in the distal part of centriolar cylinder); ITC: internal triplets connections system (scaffold structure), which include A-C linkers; CS: cartwheel structure (an axis with spokes); SDA: sub-distal appendage; HSDA: head of sub-distal appendage; SSDA: the stem of sub-distal appendage (connected to three triplets in this case); S: satellites; SS: the striated structure of sub-distal appendage stem; MT: microtubule; DA: distal appendage; HAD: head of distal appendage; R: rib. From [12] with modifications. Insertion: The fine ultrastructure of the MT triplet, showing protofilaments of MTs (the data from [15][16] were used to make this drawing).
References
- Flemming, W. Studien in der entwicklungsgeschichte der najaden. Sitz. Akad. Wissensch Wien. 1875, 71, 81–147.
- Hertwig, O. Beitrage zur kenntniss der bildung, befruchtung und theilung des thierischen eies. Morphol. Jb. 1875, 1, 347–434.
- Van Beneden, E. Recherches sur les dicyémides, survivants actuels d’un embranchement des mésozoaires. Bull. Acad. Roy. Méd. Belg. 1876, 41, 1160–1205.
- Van Beneden, E., Neyt, A. Nouvelles recherches sur la fecondation et la division mitoshes l’ascaride megalocephale. Bull. Acad. Roy. Méd. Belg. 1887, 14.
- Boveri, T. Zellenstudien i. Jena Zeit. Nat. 1887, 21.
- Boveri, T. Uber das verhalten der centrosomen bei der befruchtung des seeigeleies, etc. Verhandl. Phys. Med. Ges. Wurzbg. XXIX. 1895.
- Henneguy, L.F. Sur les rapports des cils vibratiles avec les centrosomes. Arch. Microsc. Morph. Exp. 1898, 1, 481–496.
- Von Lenhossék, M. Uber flimmerzellen. Verh. Anat. Ges., Kiel. 1898, 12, 106–128.
- Afzelius, B. Electron microscopy of the sperm tail; results obtained with a new fixative. J. Biophys. Biochem. Cytol. 1959, 5, 269–278.
- Gibbons, I.R.; Grimstone, A.V. On flagellar structure in certain flagellates. J. Biophys. Biochem. Cytol. 1960, 7, 697–716.
- Brinkley, B.R., and Stubblefield, E. Ultrastructure and interaction of the kinetochore and centriole in mitosis and meiosis In Advances in cell biology, Prescott, D.M., G.L., Mc Conkey, E., Ed. Appleton-Century Crofts: New York, 1970; Vol. 1, pp 119-184.
- Uzbekov, R.; Prigent, C. Clockwise or anticlockwise? Turning the centriole triplets in the right direction! FEBS Lett. 2007, 581, 1251–1254.
- Uzbekov, R.E., Alieva, I.B. Centrosome: History of study and new discoveries. From cytoplasmic granule to the center of intracellular regulation. Moscow University Press: Moscow, 2013.
- R. Uzbekov; Tomer Avidor-Reiss; Principal Postulates of Centrosomal Biology. Version 2020. Cells 2020, 9, 2156, 10.3390/cells9102156.
- McNitt, R. Centriole ultrastructure and its possible role in microtubule formation in an aquatic fungus. Protoplasma 1974, 80, 91–108.
- Greenan, G.A.; Keszthelyi, B.; Vale, R.D.; Agard, D.A. Insights into centriole geometry revealed by cryotomography of doublet and triplet centrioles. ELife 2018, 7: e36851.

