Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | L. A. Stanciu | -- | 2785 | 2022-04-29 14:49:19 | | | |
| 2 | Jessie Wu | Meta information modification | 2785 | 2022-05-05 11:45:23 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Feketea, G.; Bocsan, I.C.; Popescu, C.; Gaman, M.; Stanciu, L.A.; Zdrenghea, M. MiRNAs Relevant for Asthma. Encyclopedia. Available online: https://encyclopedia.pub/entry/22508 (accessed on 09 August 2026).
Feketea G, Bocsan IC, Popescu C, Gaman M, Stanciu LA, Zdrenghea M. MiRNAs Relevant for Asthma. Encyclopedia. Available at: https://encyclopedia.pub/entry/22508. Accessed August 09, 2026.
Feketea, Gavriela, Ioana Corina Bocsan, Cristian Popescu, Mihaela Gaman, L. A. Stanciu, Mihnea Zdrenghea. "MiRNAs Relevant for Asthma" Encyclopedia, https://encyclopedia.pub/entry/22508 (accessed August 09, 2026).
Feketea, G., Bocsan, I.C., Popescu, C., Gaman, M., Stanciu, L.A., & Zdrenghea, M. (2022, April 29). MiRNAs Relevant for Asthma. In Encyclopedia. https://encyclopedia.pub/entry/22508
Feketea, Gavriela, et al. "MiRNAs Relevant for Asthma." Encyclopedia. Web. 29 April, 2022.
Copy Citation
There is an imbalance in asthma between classically activated macrophages (M1 cells) and alternatively activated macrophages (M2 cells) in favor of the latter. MicroRNAs (miRNAs) play a critical role in regulating macrophage proliferation and differentiation and control the balance of M1 and M2 macrophage polarization, thereby controlling immune responses.
asthma
microRNA
macrophages
1. MiRNAs Polarizing Macrophages Towards a Pro-Inflammatory M1 Phenotype
The miRNAs that polarize macrophages towards a pro-inflammatory M1 phenotype (Table 1, Figure 1) are important in balancing the anti-inflammatory/regulatory M2 macrophages which promote Th2 immune responses. In addition, pro-inflammatory M1 macrophages promote T helper/cytotoxic type 1 antiviral responses, and virus infections are the most common cause of asthma exacerbations. An excessive macrophage pro-inflammatory response could, however, play a role in severe asthma and, particularly, in neutrophilic asthma.
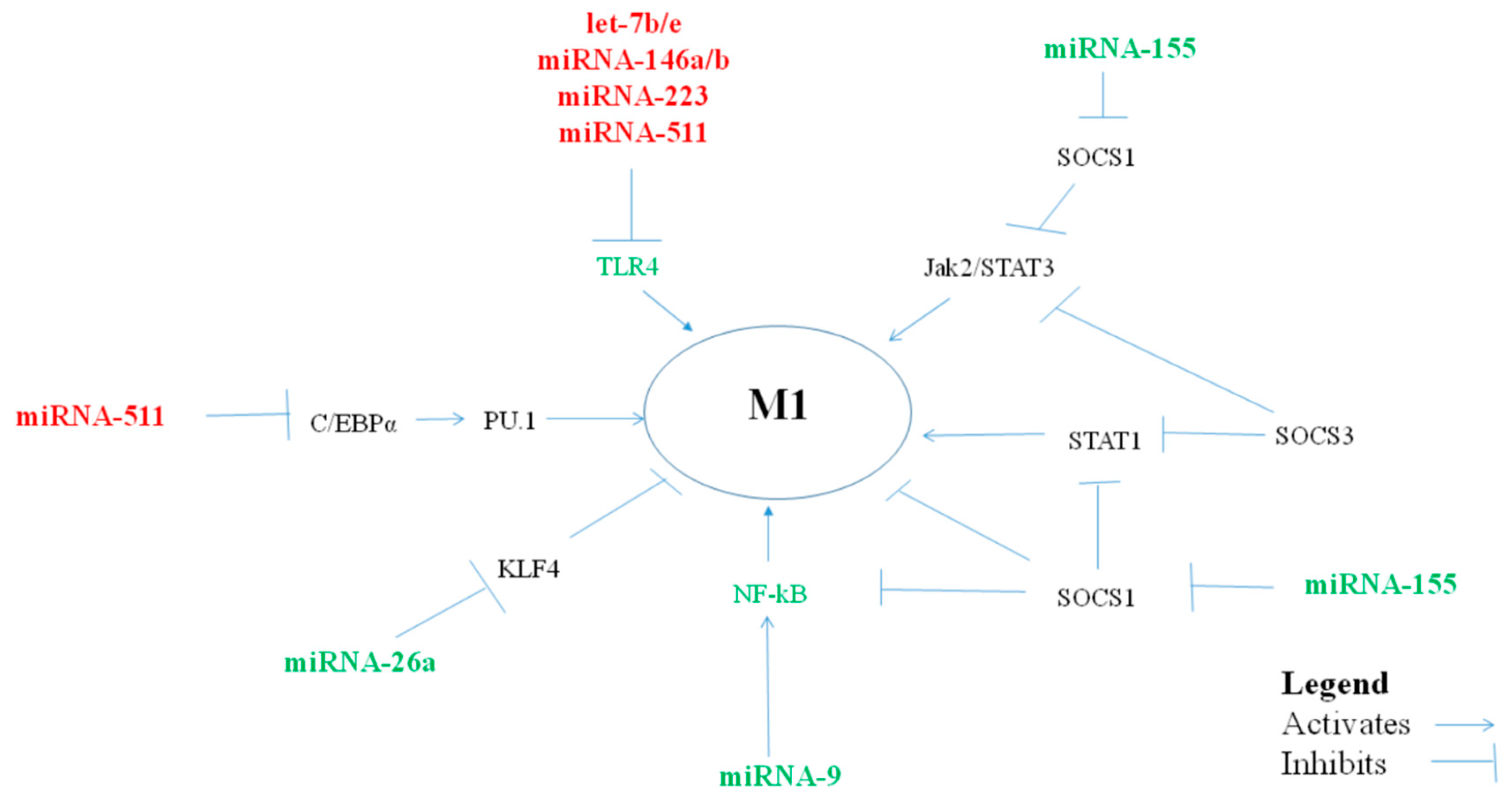
Figure 1. miRNAs, via their targets, influence pro-inflammatory M1-macrophage polarization.
Table 1. Pro-inflammatory miRNA levels in asthma.
| miRNA | Macrophage Type | miRNA Levels in Asthma | ||||||
|---|---|---|---|---|---|---|---|---|
| Polarization | Serum/Plasma | Brushing Airway Cells | Biopsies | Nasal Mucosa | Sputum | BAL Exosomes/Exhaled Breath | ||
|
let-7f |
M1/M2a |
M1/M2 |
↑[1] |
|||||
|
miRNA-9 |
M1 |
M1 |
↑[2] |
|||||
|
miRNA-18a |
M1 |
M1 |
↓[3] |
↓[4] |
||||
|
miRNA-19a |
M1 |
M1 |
↑[5] |
|||||
|
miRNA-19b-3p |
M1 |
M1 |
||||||
|
miRNA-26a/b |
M2c |
M1 |
↓[8] |
↓[6] |
↓[9] |
|||
|
miRNA-27a/b |
M2b |
M1 |
||||||
|
miRNA-125b |
M1/M2a/M2c |
M1 |
↑[8] |
|||||
|
miRNA-155 |
M1 |
M1 |
↓[8] |
↓[4] |
↓[10] |
↓[11] |
||
1.1. MiRNA let-7f
Let-7f miRNAs belong to a highly conserved let-7 (lethal-7) microRNA family consisting of 12 genes encoding for nine different miRNAs (let-7a to let-7i). Although the members share the same “seed region” (similarities in nucleotides 2–8 at their 5′ end), let-7 family members are each unique and encoded on different chromosomes [12]. Among the tumor suppressor miRNAs, reduced let-7 expression occurs most frequently in cancer and typically correlates with poor prognosis.
Let-7f appears to polarize macrophages towards an M1 phenotype.
In the THP1 cell line, pri-let-7f levels were increased in response to LPS while the corresponding mature miRNA let-7f levels were reduced. The overexpression of let-7f in THP-1 differentiated macrophages led to a reduction in SOCS4 protein levels [13].
When individual members of the let-7 family were measured, the expression of let-7f was reported to be higher in cultured epithelial brushings from asthmatic donors [1].
IL-17A is associated with severe asthma and requires IL-23R signaling, which is negatively regulated by let-7f miRNA in CD4+ lymphocytes [14]. Let-7f was negatively regulated by estrogen receptor signaling in the MCF-7 breast cancer cell line [15]. Women have an increased prevalence of severe asthma compared with men. In patients with severe asthma, IL-17A production was increased to a greater degree in TH17 cells from women compared with those from men [16]. Let-7f expression was lower and IL-23R expression was higher in TH17-differentiated cells from women compared with men with severe asthma [16].
1.2. MiRNA-9
Expression of miRNA-9 was found higher in pro-inflammatory M1 than M2 macrophages, and promoted human M1 polarization [17] and miRNA-9 levels correlated with levels of peroxisome proliferator-activated receptor-δ (PPAR-δ, also known as PPAR-β) in M1 macrophages [18].
MiRNA-9 levels were synergistically increased following IFNγ/LPS co-exposure in isolated macrophages and in vivo, in an IFN-γ/LPS–induced mouse model of steroid-resistant airway hyperresponsiveness (AHR) [2]. MiRNA-9 production decreased protein phosphatase 2A (PP2A) activity and also inhibited steroid-induced glucocorticoid receptor (GR) nuclear translocation. AntagomiRNA-mediated inhibition of miRNA-9 increased both PP2A activity and GR nuclear translocation in macrophages, and restored steroid sensitivity in isolated macrophages and in several mouse models of steroid-resistant AHR [2].
The pro-inflammatory miRNA-9 was found to be increased in sputum samples from patients with neutrophilic asthma, which is often resistant to corticosteroid therapy, compared with eosinophilic asthma [2]. Animal studies suggest that targeting miRNA-9 could be an effective treatment for steroid-resistant asthma.
1.3. MiRNA-17-18-19-20-92
The miRNA-17-92 cluster (miRNA-17-5p, miRNA-17-3p, miRNA-18a, miRNA-19a, miRNA-19b, miRNA-20a, and miRNA-92-1) or oncomiRNA-1 increases cell proliferation or inhibits apoptosis by suppressing pro-apoptotic Bcl2 protein Bim, transcription factor E2 factor 1 (E2F1), and tumor suppressor phosphate and tensin homolog (PTEN). Downregulation of miRNA-17-92 is critical for normal myeloid differentiation via induction of PU.1 [19]. PU.1 transcription factor has been reported to play an important role in the differentiation of macrophages towards the pro-inflammatory M1-like phenotype [20].
MiRNA-19 has a direct role in upregulating NF-κB signaling and pro-inflammatory cytokine production. miRNA-19a-3p is capable of inducing an M1 phenotype by targeting Fos-related antigen-1 (Fra-1) transcription factor which plays a key role in the polarization of the M1 to the M2 phenotype and, consequently, decreasing the expression of the Fra-1 downstream genes VEGF, STAT3, and pSTAT3 [21].
The available studies concerning the levels of individual miRNAs of the miRNA-17-92 cluster in different biological specimens in asthma report decreased levels of miRNA-17, miRNA-18, miRNA 19b, and miRNA-20. On the contrary, one study reported increased miRNA-19a levels. miRNA-17-5p was reported downregulated in brushing bronchial epithelial cells cultured in vitro from severe asthmatics compared to cells from healthy donors [7]. Downregulation of miRNA-18a was observed in nasal mucosa biopsies in patients with asthma in comparison with control subjects [4], and in bronchial epithelial cells, obtained by brushing and cultured in vitro, from patients with asthma with different degrees of severity compared to cells from healthy donors [3]. Upregulation of miRNA-19a was found in epithelial cells isolated from biopsies of subjects with severe asthma (only 6) compared with cells from subjects with mild asthma (9) and healthy controls (9), and miRNA-19a was shown to enhance proliferation of bronchial epithelial cells from patients with severe asthma by targeting the TGF-β receptor 2 gene [5]. The levels of miRNA-19b-3p were found to be decreased in bronchial cells obtained by bronchoscopic brushing from patients with steroid-naïve asthma [6] and in brushing bronchial epithelial cells from severe asthmatics cultured in vitro compared to cells from healthy donors [7]. miRNA-20a-5p levels were found to be downregulated in brushing bronchial epithelial cells cultured in vitro from severe asthmatics compared to cells from healthy donors [7]. In another study, miRNA-20 was expressed at the same level in primary bronchial epithelial cells of patients with severe or difficult-to-treat asthma, treated with inhaled corticosteroids and subjects without asthma [22].
1.4. MiRNA-26a/b
It was shown that miRNA-26a directly downregulated IFN-β in human macrophages [23]. miRNA-26a induced a pro-inflammatory M1-type of human macrophage activation by downregulating M2-polarizing macrophage colony-stimulating factor (M-CSF) and IL-10 expression [24]. Kruppel-like factor 4 (KLF4) was a target of miRNA-26a-5p in primary human macrophages [25].
All available studies report decreased levels of miRNA-26 in asthma. miRNA-26a has been found to be decreased in bronchial epithelial cells obtained by brushing from patients with steroid-naïve asthma [6]. Levanen and colleagues demonstrated for the first time the presence of miRNAs in exosomes from BAL fluid of both healthy and asthmatic subjects with mild intermittent stable disease [9]. They showed that exosomal miRNA profiles at baseline are different, and in asthma the expression profiles of the identified miRNAs were highly correlated with forced expiratory volume in 1 s (FEV1). Levels of miRNA-26a were downregulated in BAL fluid exosomes in patients with mild asthma [9]. Decreased expression of serum miRNA-26a was observed in a very small group of mild persistent or moderate persistent asthmatic patients (n = 10, 8/10 allergic, smokers 4/10), compared to control subjects (n = 10, 3 being allergic, smokers 5/10) [26]. In another study of the same group, the plasma levels of miRNA-26b, but not miRNA-26a, were decreased in patients with asthma [8].
1.5. MiRNA-27a/b
In human MDM, miRNA-27a expression was higher in M2b macrophages [27]. Treatment with LPS, IFN-β, or IFN-γ repressed accumulation of miRNA-27a during human monocyte-to-macrophage differentiation (IFN-β more than IFN-γ) [28]. The stimulation through Toll-like receptor TLR2/TLR4 (not TLR3) decreased miRNA-27a in human MDM (monocytes were cultured with M-CSF). Upregulation of miRNA-27a enhanced the expression of pro-inflammatory cytokines in TLR2/4-activated macrophages.
The expression levels of miRNA-27a and miRNA-27b-3p in freshly isolated bronchial epithelial brushings were lower in patients with steroid-naïve asthma and steroid-using asthma, compared to healthy control subjects [6]. miRNA-27a and miRNA-27b-3p expression was downregulated in bronchial epithelial cells obtained by brushing and cultured in vitro from patients with asthma with different degrees of severity, compared to cells obtained from healthy donors [3].
1.6. MiRNA-125b
In vitro, miRNA-125b-5p was significantly upregulated in M1 and M2a/M2c-polarized macrophages compared with unpolarized macrophages [29]. MiRNA-125b was significantly upregulated by TLR4 engagement in THP-1 cells and miRNA-125b overexpression induced M1 polarization in THP-1 cells, mimicking the IFN-γ/LPS stimulation effect [30].
Circulating plasma miRNA-125b levels were most predictive of asthmatic status, being increased in patients with asthma who had high eosinophil counts [8].
1.7. MiRNA-155
MiRNA-155 was increased in M1 and was downregulated in IL-10-polarized human M2c macrophages [17][29][31][32]. miRNA-155 promotes pro-inflammatory classical M1 activation by blocking anti-inflammatory signals and transcription factors, but it can also prevent excessive TLR signaling. Early during TLR activation, miRNA-155 expression is upregulated and inhibits the expression of the negative regulators, such as IL-10, allowing TLR signal transduction and type I IFN-mediated antiviral response. Later on, the increase in anti-inflammatory miRNA-21 induces IL-10 production, and increased IL-10 reduces miRNA-155 expression, limiting the TLR signaling pathways [33]. miRNA-155 also enhances M1-polarization by the repression of negative regulators of pro-inflammatory responses including SOCS1, SH2 domain-containing inositol 5-phosphatase 1 (SHIP1), and BCL6 in various diseases. The serine-threonine kinases Akt contribute to macrophage polarization, and miRNA-155 was found to be essential in serine-threonine kinase Akt–dependent M1/M2 polarization of macrophages (with Akt1 involved in M2- and Akt2 in M1-polarization) by targeting CCAAT/enhancer binding protein-β (C/EBPβ), a key regulator of M2 polarization [34]. SOCS2, a marker of M2, is an essential controller of macrophage activation and function, and also regulates SOCS1 and SOCS3 expression levels through proteasomal degradation [35].
In human macrophages, but not in BEAS-2B bronchial epithelial cell line, miRNA-155 downregulated the levels of IL-13Rα1, thus reducing the phosphorylation of STAT6 [36][37].
Data regarding the levels of miRNA-155 in asthma are divergent. In primary non-asthmatic and asthmatic human airway smooth muscle cells (hASMCs) isolated from non-transplantable donor lungs or resected lung tissue by enzymatic digestion, miRNA-155 expression was higher in IL-1β/TNF-α/IFN-γ-treated asthmatic cells as compared to normal cells [38].
Levels of miRNA-155 were lower in asthmatic bronchial epithelial cells obtained by brushing and cultured in vitro, than in cells from healthy donors [3]. miRNA-155 expression was also found downregulated in the nasal mucosa [4], in exhaled breath condensates [11], and in cell-free induced sputum in patients with asthma in comparison to control subjects [10]. Low levels of miRNA-155 were reported also in plasma of patients with asthma [8]. In a recent report, patients with severe asthma had higher plasma levels of miRNA-155 when compared with mild-to-moderate asthmatics and non-asthmatic control subjects, suggesting that miRNA-155 may contribute to the severity of inflammation [39].
Recent animal data showed increased expression of miRNA-155 in an ovalbumin (OVA)-induced mouse model of asthma, and lentiviral vector-delivered small interfering (si)RNA targeting miRNA-155 resulted in reduced AHR, airway inflammation, and Th2 cytokine production [40]. These data suggest that miRNA-155 could be involved in asthma severity and that targeting miRNA-155 could be a novel approach for the treatment of allergic asthma [40].
In conclusion, pro-inflammatory miRNA-9, miRNA-18/19, miRNA-26, miRNA-27, miRNA-125b, and miRNA-155 have been shown in vitro to polarize macrophages towards a pro-inflammatory M1 phenotype and their levels were generally found to be decreased in various tissues in asthma. Theoretically, M1-polarized macrophages balance M2 macrophages and play a positive role in both normal individuals and in patients with asthma. However, these pro-inflammatory miRNAs in excess may play a detrimental role in severe asthma.
MiRNA-9 was increased in patients with neutrophilic asthma and the targeting of miRNA-9 has been suggested for the treatment of steroid-resistant asthma.
MiRNA-125b and miRNA-155 were increased in severe asthma, which would suggest that in this situation targeting these miRNAs could be an additional tool in its treatment.
2. MiRNAs Polarizing Macrophages towards an anti-Inflammatory/Suppressor M2 Phenotype
Certain miRNAs may prove to play an important role in the pathogenesis of allergic inflammation in asthma by polarizing macrophages towards the M2 phenotype (Table 2, Figure 2).
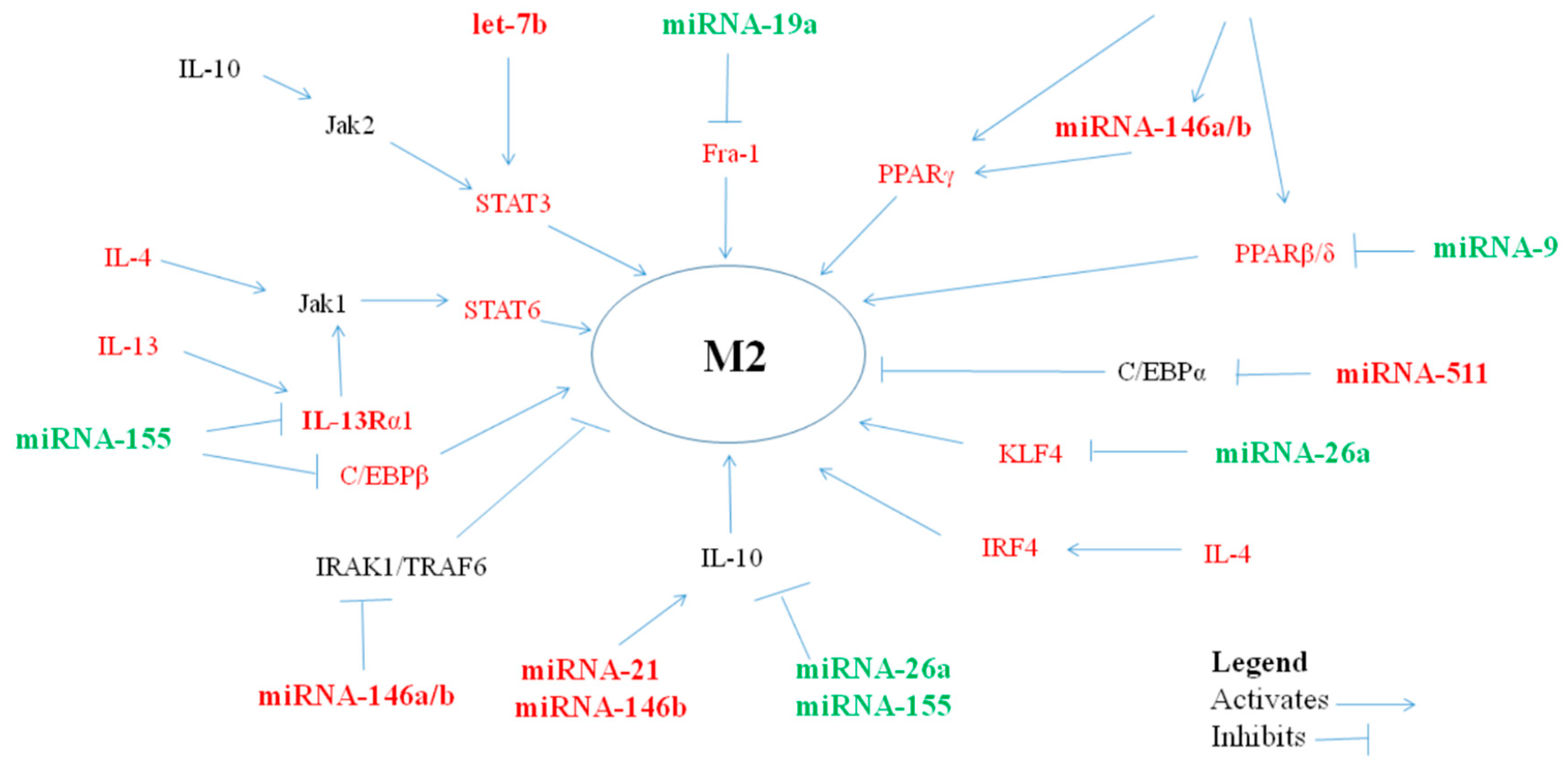
Figure 2. miRNAs, via their targets, influence anti-inflammatory M2-macrophage polarization.
Table 2. Anti-inflammatory miRNA levels in asthma.
| miRNA | Macrophage Type | miRNA Levels in Asthma | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Polarization | Serum/Plasma | Brushing Airway Cells | Biopsies | Nasal Mucosa | Sputum | BAL Exosomes/Exhaled Breath | Monocytes | ||
|
let-7a |
M2 |
M2 |
↓[26] |
||||||
|
let-7b |
M2 |
M2 |
↑[8] |
||||||
|
let-7c |
M2a |
M2 |
↑[8] |
↓[6] |
|||||
|
let-7e |
M2 |
M2 |
↑[8] |
↓[4] |
|||||
|
miRNA-21 |
M2c |
M2 |
↑[8] |
↑[41] |
|||||
|
miRNA-34 |
M2c |
M2 |
↓[42] |
↓[6] |
|||||
|
miRNA-124a |
M2b |
M2 |
↑[31] |
||||||
|
miRNA-146a |
M1 |
M2 |
↑[8] |
↓[43] |
|||||
|
miRNA-223 |
M2 |
↑[8] |
↑[44] |
||||||
|
miRNA-511 |
M2a |
M2 |
↓[45] |
||||||
2.1. MiRNA let-7a/b/c/d/e
The miRNA let-7 family members are generally implicated in regulating the TLR/NF-κB signal pathway and are known as tumor suppressor miRNAs, participating in cell differentiation, proliferation, and apoptosis [46]. The levels of let-7a/b/c/e were found upregulated in tumor-associated macrophages (TAMs), which displayed characteristics of anti-inflammatory M2 macrophages [47].
Levels of let-7a were significantly decreased in exosomes isolated from BAL fluid in patients with mild intermittent asthma [9] and in exhaled breath condensates from patients with asthma [11]. Serum levels of miRNA-7a were reported to be decreased in patients with asthma [26].
In human macrophages, IFN-β upregulated the expression of let-7b, and let-7b directly downregulated IFN-β, suggesting a negative feedback loop [23]. LPS-preconditioned mesenchymal stromal cells containing exosome (extracellular vesicles derived from cell endocytosis which act as transmitters between cells)-shuttled let-7b polarized THP-1 cells to a M2 phenotype via TLR4/NF-κB/STAT3/AKT regulatory signaling pathway [50]. The upregulation of let-7b is characteristic of prostatic TAMs. The downregulation of let-7b in TAMs leads to changes in the expression profiles of cytokines, such as those of IL-12, IL-23, IL-10, and TNF-α [47].
In contrast to reports of mouse studies, TLR7/TLR8 activation triggered monocytes to differentiate into a M2 macrophage surface phenotype with a mixed M1/M2 cytokine secretion profile [51].
Treatment with antagomiRNA against let-7b in a house dust mite-induced model of AHR did not suppress features of the disease [52].
Plasma let-7b expression was found increased in patients with asthma who had high eosinophil counts [8].
Levels of let-7c were found decreased in bronchial cells obtained by bronchoscopic brushing from patients with steroid-naïve asthma [6]. let-7c levels were reported increased in plasma in asthmatic patients with high eosinophil counts [8].
Serum miRNA let-7d levels were reported decreased in patients with asthma [26].
let-7e targeted STAT3 inhibitor SOCS1 in human suppressor antigen-presenting cells (obtained by monocyte stimulation with the TLR7/8- and NLR-activator R848 (Resiquimod)) [53]. In primary nasal epithelial cells from patients with allergic rhinitis, one pathway of the anti-inflammatory effect of miRNA-let-7e was via activation of the JAK1/STAT3 pathway, which caused a decrease in SOCS4 expression [54].
In human THP-1 cells, let-7e targeted TLR4 and pro-inflammatory cytokines, suggesting a feedback mechanism TLR4-let-7e-TLR4 [49].
In the nasal mucosa of patients with asthma, let-7e expression was lower than in control subjects [4], but plasma let-7e levels were reported increased in asthmatic patients with high eosinophil counts [8].
2.2. MiRNA-21
In in vitro IL-10-stimulated macrophages (i.e., M2c macrophages), miRNA-21 was shown to be increased [29]. The miRNA-21-containing exosomes, engulfed in vitro by CD14+ human monocytes, suppressed the expression of M1 and increased that of M2 markers [55].
By targeting transcripts of proteins that regulate cell division and apoptosis, such as PTEN and programmed cell death 4 (PDCD4) (negative regulators of cell death and cell cycle, respectively) miRNA-21 is considered an oncomiRNA. miRNA-21 inhibited NF-kB and the production of IL-6 and increased IL-10 indirectly, via downregulation of the pro-inflammatory molecule tumor suppressor PDCD4, a negative regulator of AP-1 and IL-10, in a macrophage cell line and bone marrow-derived macrophages [56][57].
Upregulation of miRNA-21 was observed in diphenylcyclopropenone-challenged skin biopsies in human allergic contact dermatitis [58].
The data available concerning miRNA-21 levels in human adult asthma are conflicting, with different degrees of modulation being observed in different cell types and tissues.
Levels of miRNA-21 were found decreased in exosomes from BAL and exhaled breath condensates from patients with asthma [9][11].
In contrast, miRNA-21 expression was increased in cultured epithelial cell specimens collected with a tracheal swab in patients with asthma regardless of treatment with inhaled corticosteroids (ICS) compared to the normal subjects, and was significantly higher in the ICS non-treated than the ICS-treated patients with asthma [41]. When these epithelial cells were cultured in vitro with IL-13, miRNA-21 expression increased with increasing IL-13 concentration.
Upregulation of plasma miRNA-21 was also reported in eosinophilic asthma, and it was suggested that miRNA-21 represents a profile of type 2 airway inflammation [8].
The miRNA-21 seems to reflect the type 2 environment characteristic to asthma, and targeting miRNA-21 could be beneficial in asthma [41].
References
- Jardim, M.J.; Dailey, L.; Silbajoris, R.; Diaz-Sanchez, D. Distinct microRNA expression in human airway cells of asthmatic donors identifies a novel asthma-associated gene. Am. J. Respir. Cell Mol. Biol. 2012, 47, 536–542.
- Li, J.J.; Tay, H.L.; Maltby, S.; Xiang, Y.; Eyers, F.; Hatchwell, L.; Zhou, H.; Toop, H.D.; Morris, J.C.; Nair, P.; et al. MicroRNA-9 regulates steroid-resistant airway hyperresponsiveness by reducing protein phosphatase 2A activity. J. Allergy Clin. Immunol. 2015, 136, 462–473.
- Martinez-Nunez, R.T.; Bondanese, V.P.; Louafi, F.; Francisco-Garcia, A.S.; Rupani, H.; Bedke, N.; Holgate, S.; Howarth, P.H.; Davies, D.E.; Sanchez-Elsner, T. A microRNA network dysregulated in asthma controls IL-6 production in bronchial epithelial cells. PLoS One 2014, 9, e111659.
- Suojalehto, H.; Lindstrom, I.; Majuri, M.L.; Mitts, C.; Karjalainen, J.; Wolff, H.; Alenius, H. Altered microRNA expression of nasal mucosa in long-term asthma and allergic rhinitis. Int. Arch. Allergy Immunol. 2014, 163, 168–178.
- Haj-Salem, I.; Fakhfakh, R.; Berube, J.C.; Jacques, E.; Plante, S.; Simard, M.J.; Bosse, Y.; Chakir, J. MicroRNA-19a enhances proliferation of bronchial epithelial cells by targeting TGFbetaR2 gene in severe asthma. Allergy 2015, 70, 212–219.
- Solberg, O.D.; Ostrin, E.J.; Love, M.I.; Peng, J.C.; Bhakta, N.R.; Hou, L.; Nguyen, C.; Solon, M.; Nguyen, C.; Barczak, A.J.; et al. Airway epithelial miRNA expression is altered in asthma. Am. J. Respir. Crit. Care Med. 2012, 186, 965–974.
- Martinez-Nunez, R.T.; Rupani, H.; Plate, M.; Niranjan, M.; Chambers, R.C.; Howarth, P.H.; Sanchez-Elsner, T. Genome-Wide Posttranscriptional Dysregulation by MicroRNAs in Human Asthma as Revealed by Frac-seq. J. Immunol. 2018, 201, 251–263.
- Panganiban, R.P.; Wang, Y.; Howrylak, J.; Chinchilli, V.M.; Craig, T.J.; August, A.; Ishmael, F.T. Circulating microRNAs as biomarkers in patients with allergic rhinitis and asthma. J. Allergy Clin. Immunol. 2016, 137, 1423–1432.
- Levanen, B.; Bhakta, N.R.; Torregrosa Paredes, P.; Barbeau, R.; Hiltbrunner, S.; Pollack, J.L.; Skold, C.M.; Svartengren, M.; Grunewald, J.; Gabrielsson, S.; et al. Altered microRNA profiles in bronchoalveolar lavage fluid exosomes in asthmatic patients. J. Allergy Clin. Immunol. 2013, 131, 894–903.
- Malmhall, C.; Johansson, K.; Winkler, C.; Alawieh, S.; Ekerljung, L.; Radinger, M. Altered miR-155 Expression in Allergic Asthmatic Airways. Scand. J.Immunol. 2017, 85, 300–307.
- Pinkerton, M.; Chinchilli, V.; Banta, E.; Craig, T.; August, A.; Bascom, R.; Cantorna, M.; Harvill, E.; Ishmael, F.T. Differential expression of microRNAs in exhaled breath condensates of patients with asthma, patients with chronic obstructive pulmonary disease, and healthy adults. J. Allergy Clin. Immunol. 2013, 132, 217–219.
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516.
- Naqvi, A.R.; Fordham, J.B.; Khan, A.; Nares, S. MicroRNAs responsive to Aggregatibacter actinomycetemcomitans and Porphyromonas gingivalis LPS modulate expression of genes regulating innate immunity in human macrophages. Innate Immune. 2014, 20, 540–551.
- Li, Z.; Wu, F.; Brant, S.R.; Kwon, J.H. IL-23 receptor regulation by Let-7f in human CD4+ memory T cells. J. Immunol. 2011, 186, 6182–6190.
- Bhat-Nakshatri, P.; Wang, G.; Collins, N.R.; Thomson, M.J.; Geistlinger, T.R.; Carroll, J.S.; Brown, M.; Hammond, S.; Srour, E.F.; Liu, Y.; et al. Estradiol-regulated microRNAs control estradiol response in breast cancer cells. Nucleic Acids Res. 2009, 37, 4850–4861.
- Newcomb, D.C.; Cephus, J.Y.; Boswell, M.G.; Fahrenholz, J.M.; Langley, E.W.; Feldman, A.S.; Zhou, W.; Dulek, D.E.; Goleniewska, K.; Woodward, K.B.; et al. Estrogen and progesterone decrease let-7f microRNA expression and increase IL-23/IL-23 receptor signaling and IL-17A production in patients with severe asthma. J. Allergy Clin. Immunol. 2015, 136, 1025–1034.
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.C. MiRNA-Mediated Macrophage Polarization and its Potential Role in the Regulation of Inflammatory Response. Shock 2016, 46, 122–131.
- Thulin, P.; Wei, T.; Werngren, O.; Cheung, L.; Fisher, R.M.; Grander, D.; Corcoran, M.; Ehrenborg, E. MicroRNA-9 regulates the expression of peroxisome proliferator-activated receptor delta in human monocytes during the inflammatory response. Int. J. Mol. Med. 2013, 31, 1003–1010.
- Yan, Y.; Hanse, E.A.; Stedman, K.; Benson, J.M.; Lowman, X.H.; Subramanian, S.; Kelekar, A. Transcription factor C/EBP-beta induces tumor-suppressor phosphatase PHLPP2 through repression of the miR-17-92 cluster in differentiating AML cells. Cell Death Differ. 2016, 23, 1232–1242.
- Juhas, U.; Ryba-Stanislawowska, M.; Szargiej, P.; Mysliwska, J. Different pathways of macrophage activation and polarization. Postepy. Hig. Med. Dosw. 2015, 69, 496–502.
- Yang, J.; Zhang, Z.; Chen, C.; Liu, Y.; Si, Q.; Chuang, T.H.; Li, N.; Gomez-Cabrero, A.; Reisfeld, R.A.; Xiang, R.; et al. MicroRNA-19a-3p inhibits breast cancer progression and metastasis by inducing macrophage polarization through downregulated expression of Fra-1 proto-oncogene. Oncogene 2014, 33, 3014–3023.
- Moheimani, F.; Koops, J.; Williams, T.; Reid, A.T.; Hansbro, P.M.; Wark, P.A.; Knight, D.A. Influenza A virus infection dysregulates the expression of microRNA-22 and its targets; CD147 and HDAC4, in epithelium of asthmatics. Respir. Res. 2018, 19, 145.
- Witwer, K.W.; Sisk, J.M.; Gama, L.; Clements, J.E. MicroRNA regulation of IFN-beta protein expression: rapid and sensitive modulation of the innate immune response. J. Immunol. 2010, 184, 2369–2376.
- Chai, Z.T.; Zhu, X.D.; Ao, J.Y.; Wang, W.Q.; Gao, D.M.; Kong, J.; Zhang, N.; Zhang, Y.Y.; Ye, B.G.; Ma, D.N.; et al. microRNA-26a suppresses recruitment of macrophages by down-regulating macrophage colony-stimulating factor expression through the PI3K/Akt pathway in hepatocellular carcinoma. J. Hematol. Oncol. 2015, 8, 56.
- Sahu, S.K.; Kumar, M.; Chakraborty, S.; Banerjee, S.K.; Kumar, R.; Gupta, P.; Jana, K.; Gupta, U.D.; Ghosh, Z.; Kundu, M.; et al. MicroRNA 26a (miR-26a)/KLF4 and CREB-C/EBPbeta regulate innate immune signaling, the polarization of macrophages and the trafficking of Mycobacterium tuberculosis to lysosomes during infection. PLoS Pathog. 2017, 13, e1006410.
- Panganiban, R.P.; Pinkerton, M.H.; Maru, S.Y.; Jefferson, S.J.; Roff, A.N.; Ishmael, F.T. Differential microRNA epression in asthma and the role of miR-1248 in regulation of IL-5. Am. J. Clin. Exp. Immunol. 2012, 1, 154–165.
- Graff, J.W.; Dickson, A.M.; Clay, G.; McCaffrey, A.P.; Wilson, M.E. Identifying functional microRNAs in macrophages with polarized phenotypes. J. Biol. Chem. 2012, 287, 21816–21825.
- Eigsti, R.L.; Sudan, B.; Wilson, M.E.; Graff, J.W. Regulation of activation-associated microRNA accumulation rates during monocyte-to-macrophage differentiation. J. Biol. Chem. 2014, 289, 28433–28447.
- Cobos Jimenez, V.; Bradley, E.J.; Willemsen, A.M.; van Kampen, A.H.; Baas, F.; Kootstra, N.A. Next-generation sequencing of microRNAs uncovers expression signatures in polarized macrophages. Physiol. Genom. 2014, 46, 91–103.
- Duroux-Richard, I.; Roubert, C.; Ammari, M.; Presumey, J.; Grun, J.R.; Haupl, T.; Grutzkau, A.; Lecellier, C.H.; Boitez, V.; Codogno, P.; et al. miR-125b controls monocyte adaptation to inflammation through mitochondrial metabolism and dynamics. Blood 2016, 128, 3125–3136.
- Veremeyko, T.; Siddiqui, S.; Sotnikov, I.; Yung, A.; Ponomarev, E.D. IL-4/IL-13-dependent and independent expression of miR-124 and its contribution to M2 phenotype of monocytic cells in normal conditions and during allergic inflammation. PLoS ONE 2013, 8, e81774.
- Jablonski, K.A.; Gaudet, A.D.; Amici, S.A.; Popovich, P.G.; Guerau-de-Arellano, M. Control of the Inflammatory Macrophage Transcriptional Signature by miR-155. PLoS ONE 2016, 11, e0159724.
- McCoy, C.E.; Sheedy, F.J.; Qualls, J.E.; Doyle, S.L.; Quinn, S.R.; Murray, P.J.; O’Neill, L.A. IL-10 inhibits miR-155 induction by toll-like receptors. J. Biol. Chem. 2010, 285, 20492–20498.
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014.
- McCormick, S.M.; Heller, N.M. Regulation of Macrophage, Dendritic Cell, and Microglial Phenotype and Function by the SOCS Proteins. Front. Immunol. 2015, 6, 549.
- Martinez-Nunez, R.T.; Louafi, F.; Sanchez-Elsner, T. The interleukin 13 (IL-13) pathway in human macrophages is modulated by microRNA-155 via direct targeting of interleukin 13 receptor alpha1 (IL13Ralpha1). J. Biol. Chem. 2011, 286, 1786–1794.
- Matsukura, S.; Osakabe, Y.; Sekiguchi, A.; Inoue, D.; Kakiuchi, Y.; Funaki, T.; Yamazaki, Y.; Takayasu, H.; Tateno, H.; Kato, E.; et al. Overexpression of microRNA-155 suppresses chemokine expression induced by Interleukin-13 in BEAS-2B human bronchial epithelial cells. Allergol. Int. 2016, 65, S17–23.
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. Cyclooxygenase-2 and microRNA-155 expression are elevated in asthmatic airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2015, 52, 438–447.
- Qiu, L.; Zhang, Y.; Do, D.C.; Ke, X.; Zhang, S.; Lambert, K.; Kumar, S.; Hu, C.; Zhou, Y.; Ishmael, F.T.; et al. miR-155 Modulates Cockroach Allergen- and Oxidative Stress-Induced Cyclooxygenase-2 in Asthma. J. Immunol. 2018, 201, 916–929.
- Chen, H.; Xu, X.; Cheng, S.; Xu, Y.; Qi, X.; Cao, Y.; Xie, J.; Wang, C.Y.; Xu, Y.; Xiong, W. Small interfering RNA directed against microRNA-155 delivered by a lentiviral vector attenuates asthmatic features in a mouse model of allergic asthma. Exp. Ther. Med. 2017, 14, 4391–4396.
- Wu, X.B.; Wang, M.Y.; Zhu, H.Y.; Tang, S.Q.; You, Y.D.; Xie, Y.Q. Overexpression of microRNA-21 and microRNA-126 in the patients of bronchial asthma. Int. J. Clin. Exp. Med. 2014, 7, 1307–1312.
- Yin, H.; Zhang, S.; Sun, Y.; Li, S.; Ning, Y.; Dong, Y.; Shang, Y.; Bai, C. MicroRNA-34/449 targets IGFBP-3 and attenuates airway remodeling by suppressing Nur77-mediated autophagy. Cell death Dis. 2017, 8, e2998.
- Faiz, A.; Weckmann, M.; Tasena, H.; Vermeulen, C.J.; Van den Berge, M.; Ten Hacken, N.H.T.; Halayko, A.J.; Ward, J.P.T.; Lee, T.H.; Tjin, G.; et al. Profiling of healthy and asthmatic airway smooth muscle cells following interleukin-1beta treatment: a novel role for CCL20 in chronic mucus hypersecretion. Eur. Respir. J. 2018, 52.
- Maes, T.; Cobos, F.A.; Schleich, F.; Sorbello, V.; Henket, M.; De Preter, K.; Bracke, K.R.; Conickx, G.; Mesnil, C.; Vandesompele, J.; et al. Asthma inflammatory phenotypes show differential microRNA expression in sputum. J. Allergy. Clin. Immunol. 2016, 137, 1433–1446.
- Zhou, Y.; Do, D.C.; Ishmael, F.T.; Squadrito, M.L.; Tang, H.M.; Tang, H.L.; Hsu, M.H.; Qiu, L.; Li, C.; Zhang, Y.; et al. Mannose receptor modulates macrophage polarization and allergic inflammation through miR-511-3p. J. Allergy Clin. Immunol. 2018, 141, 350–364.
- Choudhury, S.N.; Li, Y. miR-21 and let-7 in the Ras and NF-kappaB pathways. Microrna 2012, 1, 65–69.
- Wang, Z.; Xu, L.; Hu, Y.; Huang, Y.; Zhang, Y.; Zheng, X.; Wang, S.; Wang, Y.; Yu, Y.; Zhang, M.; et al. miRNA let-7b modulates macrophage polarization and enhances tumor-associated macrophages to promote angiogenesis and mobility in prostate cancer. Sci. Rep. 2016, 6, 25602.
- He, X.; Jing, Z.; Cheng, G. MicroRNAs: new regulators of Toll-like receptor signalling pathways. BioMed. Res. INT. 2014, 2014, 945169.
- Curtale, G.; Renzi, T.A.; Mirolo, M.; Drufuca, L.; Albanese, M.; De Luca, M.; Rossato, M.; Bazzoni, F.; Locati, M. Multi-Step Regulation of the TLR4 Pathway by the miR-125a~99b~let-7e Cluster. Front. Immunol. 2018, 9, 2037.
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 308.
- Saha, B.; Kodys, K.; Adejumo, A.; Szabo, G. Circulating and Exosome-Packaged Hepatitis C Single-Stranded RNA Induce Monocyte Differentiation via TLR7/8 to Polarized Macrophages and Fibrocytes. J. Immunol. 2017, 198, 1974–1984.
- Collison, A.; Mattes, J.; Plank, M.; Foster, P.S. Inhibition of house dust mite-induced allergic airways disease by antagonism of microRNA-145 is comparable to glucocorticoid treatment. J. Allergy Clin. Immunol. 2011, 128, 160–167.
- Hildebrand, D.; Eberle, M.E.; Wolfle, S.M.; Egler, F.; Sahin, D.; Sahr, A.; Bode, K.A.; Heeg, K. Hsa-miR-99b/let-7e/miR-125a Cluster Regulates Pathogen Recognition Receptor-Stimulated Suppressive Antigen-Presenting Cells. Front. Immunol. 2018, 9, 1224.
- Li, L.; Zhang, S.; Jiang, X.; Liu, Y.; Liu, K.; Yang, C. MicroRNA-let-7e regulates the progression and development of allergic rhinitis by targeting suppressor of cytokine signaling 4 and activating Janus kinase 1/signal transducer and activator of transcription 3 pathway. Exp. Ther. Med. 2018, 15, 3523–3529.
- Hsieh, C.H.; Tai, S.K.; Yang, M.H. Snail-overexpressing Cancer Cells Promote M2-Like Polarization of Tumor-Associated Macrophages by Delivering MiR-21-Abundant Exosomes. Neoplasia 2018, 20, 775–788.
- Sheedy, F.J.; Palsson-McDermott, E.; Hennessy, E.J.; Martin, C.; O’Leary, J.J.; Ruan, Q.; Johnson, D.S.; Chen, Y.; O’Neill, L.A. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat. Immunol. 2010, 11, 141–147.
- Sheedy, F.J. Turning 21: Induction of miR-21 as a Key Switch in the Inflammatory Response. Front. Immunol. 2015, 6, 19.
- Vennegaard, M.T.; Bonefeld, C.M.; Hagedorn, P.H.; Bangsgaard, N.; Lovendorf, M.B.; Odum, N.; Woetmann, A.; Geisler, C.; Skov, L. Allergic contact dermatitis induces upregulation of identical microRNAs in humans and mice. Contact Dermat. 2012, 67, 298–305.
More
Information
Subjects:
Allergy
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
05 May 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No