Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hernando P. Bacosa | -- | 10809 | 2022-04-02 13:48:48 | | | |
| 2 | Camila Xu | -45 word(s) | 10764 | 2022-04-06 03:26:53 | | | | |
| 3 | Camila Xu | Meta information modification | 10764 | 2022-04-06 04:23:21 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Bacosa, H.P.; Ancla, S.M.; Arcadio, C.G.L.; Dalogdog, J.R.; Ellos, D.M.; Hayag, H.D.; Jarabe, J.G.; , .; Navarro, C.K.; Palma, M.P.; et al. Spilled Oil in Marine Environment. Encyclopedia. Available online: https://encyclopedia.pub/entry/21317 (accessed on 08 August 2026).
Bacosa HP, Ancla SM, Arcadio CGL, Dalogdog JR, Ellos DM, Hayag HD, et al. Spilled Oil in Marine Environment. Encyclopedia. Available at: https://encyclopedia.pub/entry/21317. Accessed August 08, 2026.
Bacosa, Hernando P., Sheila Mae Ancla, Cris Gel Loui Arcadio, John Russel Dalogdog, Dioniela Mae Ellos, Heather Dale Hayag, Jiza Gay Jarabe, , Carl Kenneth Navarro, Mae Princess Palma, et al. "Spilled Oil in Marine Environment" Encyclopedia, https://encyclopedia.pub/entry/21317 (accessed August 08, 2026).
Bacosa, H.P., Ancla, S.M., Arcadio, C.G.L., Dalogdog, J.R., Ellos, D.M., Hayag, H.D., Jarabe, J.G., , ., Navarro, C.K., Palma, M.P., Romarate Ii, R., Similatan, K., Yurong, S.N., Mabuhay-Omar, J., Inoue, C., & Adhikari, P.L. (2022, April 02). Spilled Oil in Marine Environment. In Encyclopedia. https://encyclopedia.pub/entry/21317
Bacosa, Hernando P., et al. "Spilled Oil in Marine Environment." Encyclopedia. Web. 02 April, 2022.
Copy Citation
Oil spill is a widespread environmental problem with significant ecological impacts. When oil is spilled in the sea, it undergoes different degradation processes such as dispersion, photooxidation, evaporation, dissolution, emulsion, biodegradation and other weathering processes. The most important among these processes is biodegradation, which is facilitated by naturally occurring oil-degrading bacteria. The interaction of bacteria with hydrocarbons in oil is affected by different environmental factors. Several studies have evaluated the effects of each factor on the biodegradation of the oil by bacterial, but none has taken all these factors together.
oil spills
oil weathering
bioremediation
polycyclic aromatic hydrocarbons (PAHs)
1. Introduction
Marine environmental pollution has been closely associated with rapid industrialization and related human activities, including numerous incidences of oil spills. While the demand for petroleum has significantly increased in the past few decades, the usage and transportation of petroleum hydrocarbons began around the nineteenth century [1][2]. Recently, there has been significant investment towards electric vehicles, however, the demand for crude oil is still growing in part due to its utility in other industries. Thus, accidental and sometimes intentional oil spills can be a great danger to marine ecosystems and the health of people in the surrounding community [1]. Oil spills can occur during exploration, drilling, transportation, refining and production of petroleum, as well as during times of conflict. For example, the Deepwater Horizon (DWH) oil spill in 2010 occurred following an accidental explosion at a deep-sea drill site, which released over 200 million gallons (or 4.9 million barrels) of crude oil into the northern Gulf of Mexico [3][4]. The fate of the spilled oil in marine environments depends on the composition of the oil, the weathering processes, and the prevailing environmental conditions [5]. To assess the risk of oil spills in coastal-marine environments, it is important to understand the fate of spilled oil, as well as the persistence and degradation of potentially toxic constituent compounds in the spilled oil.
Marine oil spills are considered to have significant negative environmental, social, economic and health impacts. The potential health outcome caused by oil spills can be divided into different categories: the effect of the toxic/carcinogenic compounds to the workers involved in cleaning spilled oils; effects to visitors and the community as a whole; mental health problem caused by its social and economic disruption; and harm to the natural ecosystems [6]. Short-term human health problems of oil exposure may include irritations, dizziness, and nausea; long-term effects may include damage to the respiratory, renal, endocrine, neurologic and hematologic systems [6]. As recently seen during the DWH oil spill, fires, explosions and sinking of ships/oil rigs are also associated with oil spills, which can pose great harm to the workers and economic losses to the company [7]. The spilled oil affects the environment by one or more mechanisms, such as physical smothering limiting locomotion or impacting physiological functions; chemical toxicity giving rise to death or impairment of cellular functions; ecological changes, including loss of key organisms from the community and the takeover of habitats by opportunistic species; lastly, indirect effects, such as the loss of habitat [8][9]. Marine wildlife (e.g., seabirds, dolphins, whales, turtles, manatees) can be greatly affected by marine oil spills. For example, the death of seabirds due to oil slicks is of increasing concern, particularly as at-sea mortality is difficult to routinely monitor leading to potential underestimations of seabird mortality following an oil spill [10].
When an oil spill occurs in offshore locations, a primary goal is to prevent the oil from reaching ecologically and financially important coastal habitats (including the coral reefs and the marshes) and to prevent it from affecting the onshore ecosystems [11]. The primary prevention mechanism can include physical blocking of oil sheen, using floats, barrier walls or booms. Dispersants are also used to chemically disrupt the oil sheens by creating smaller droplets of oil which are more easily dispersed in water. For example, the environmental impact of the DWH oil spill was partially mitigated by applying Corexit 9500 dispersant on the surface waters (with the aid of planes and small vessels) and by injecting dispersant at the site of the spill, a deep-sea wellhead [4][12]. Another way to reduce the impact of an oil spill is the controlled burning of oil on the site of the spill [11].
Crude oils are typically composed of a large number of hydrocarbons, and their relative compositions vary widely depending upon the source of the crude oil. Hydrocarbons are a group of chemical compounds consisting of carbon and hydrogen atoms linked with covalent bonds that make them highly stable. Due to a lack of functional groups, these compounds show weak solubility in water [13]. Crude oil is the most reduced form of natural hydrocarbons [14], and these hydrocarbons are one of the great sources of energy for daily living and many industries [15]. In general, crude oils contain alkanes and aromatic hydrocarbons, and resins and asphaltene hydrocarbons. Typically, the elemental composition of crude oil is approximately 83–87% carbon, 11–16% hydrogen, 0–4% oxygen plus nitrogen, and 0–4% sulfur [11].
Some of the constituents of crude oil belong to the family of neurotoxic and xenobiotic organic pollutants [16][17], which are known to be toxic and carcinogenic. For example, select polycyclic aromatic hydrocarbons (PAHs) in crude oils, consisting of two or more fused benzene rings, are known to be carcinogens and neurotoxic [15][18], thus are the major contributors to toxicity during and after any spill. Because of their different metabolic pathways, PAHs produce metabolites with oxidative and carcinogenic properties and have the ability to attack and bind to DNA and proteins [1][19][20]. As such, it is necessary to assess the capacity for compounds, such as PAHs, to degrade over time and space in order to predict toxicity.
Once oil spills into a coastal-marine environment, various natural processes act on spilled oil and oil residues over time to remove or degrade the oil compounds in the aquatic environment. These natural processes are called oil weathering, which include evaporation, dispersion, photooxidation, oxidation, emulsification and biodegradation (Figure 1). Over longer periods, oil weathering processes contribute substantially to the reduction of spilled oil in the natural environment. Immediately following an oil spill, the volatile fractions (such as low molecular weight alkanes and monoaromatic hydrocarbons) evaporate, leaving the dense oil on the water surface as a slick which will undergo a series of physical and chemical changes known as weathering process. When waves and turbulent events happen, the oil slick will generate droplets by the virtue of their buoyancy onto the water column. Thus, the large droplets will eventually resurface while the smaller droplet will spread and diffuse into the water column. Although, this small droplet will remain in the subsurface for an extended period of time and can be resurfaced when there is a decrease in wave turbulence [21]. The effects of toxicity of these oil residues on marine organisms and humans depend on a multitude of factors; e.g., composition (physical and chemical) of the residues, degree of weathering, exposure route and bioavailability [1][22][23][24]. A dominant process responsible for the removal of petroleum compounds in the oil residues from the environment is believed to be biodegradation [15].
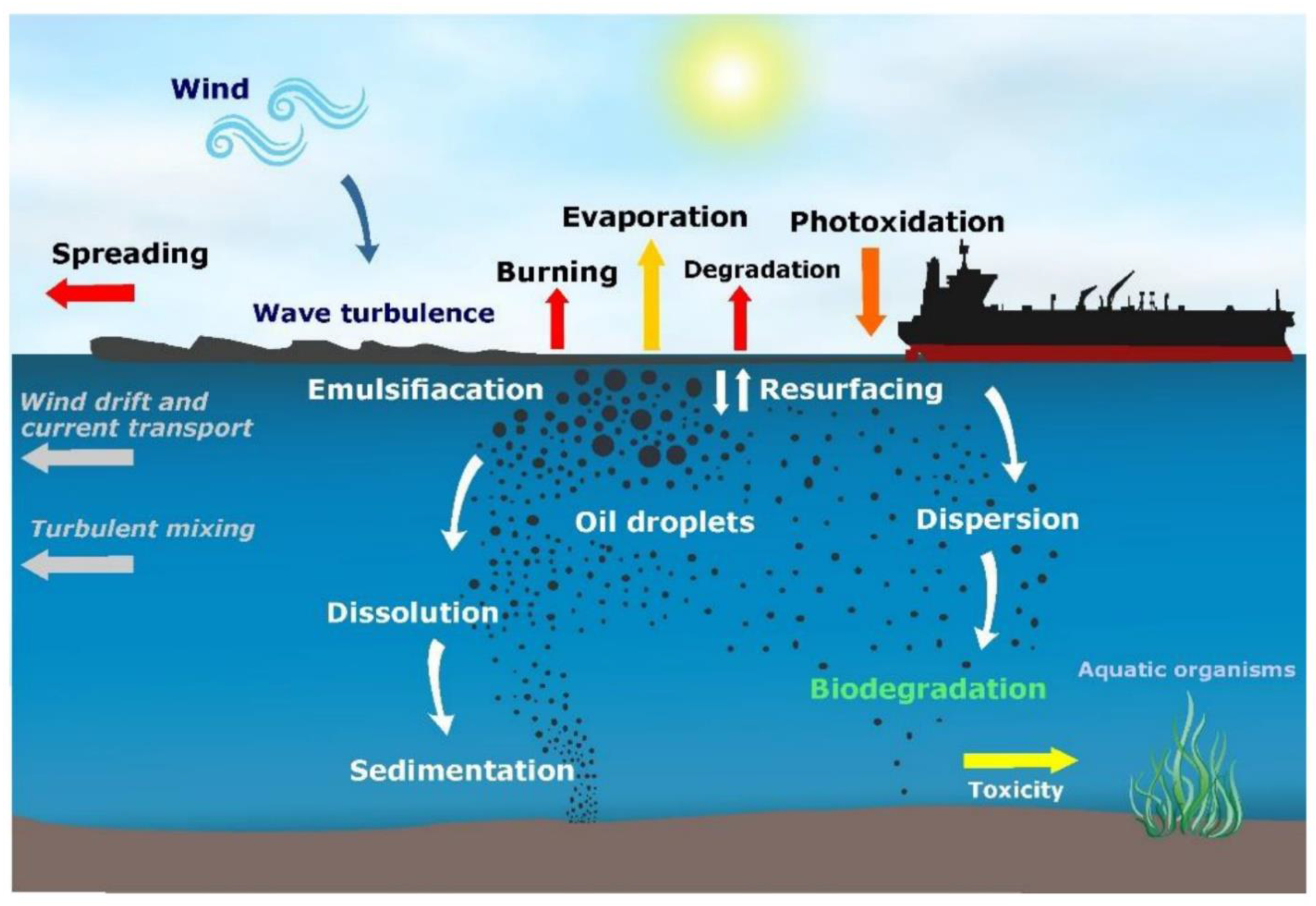 Figure 1. Transportation, weathering and fate of spilled oil in the marine environment. Adapted from Keramea et al. [21].
Figure 1. Transportation, weathering and fate of spilled oil in the marine environment. Adapted from Keramea et al. [21].Oil and oil residues in coastal-marine environments can be acted upon by naturally-occurring bacterial communities, ultimately leading to the disintegration of the oil components into carbon dioxide and water [25][26]. A variety of microorganisms are known to have hydrocarbon biodegradation capacity, e.g., hydrocarbonoclastic bacteria. These microorganisms are naturally present in coastal-marine environments, even before the occurrence of oil spills, but they are mostly in low abundance in natural, unpolluted environments [27][28][29]. They are widely distributed within surface water, sediments, and soil habitats, and the population of these bacteria rapidly increases after oil spills [30][31][32]. Hydrocarbons are readily degraded under favorable conditions, nevertheless, there is a clear preference for the catabolism of some molecules before others; hence the composition of fuel or crude oil changes as biodegradation proceeds [16][26][33]. Oil biodegradation can occur, albeit slower, even in deep and in areas with extremely low temperatures [27][34]. As such, bioremediation of hydrocarbon pollutants is advantageous following an oil spill. [35].
2. Factors Affecting Biodegradation of Oil Residues in Marine Environments
Despite the natural abundance of hydrocarbon-degrading bacteria, oil biodegradation and the interaction of bacteria and hydrocarbons are affected by various environmental factors. These factors include temperature, nutrients, sunlight, oxygen, type of oil, oil concentration, pressure, shoreline energy, pH, bacterial community, mineral particles, and salinity. This entry provides a comprehensive review of how these factors affect the bacterial activity and biodegradation of spilled oil and hydrocarbons in marine environments.
2.1. Temperature
The physical nature and chemical composition of oil residues, the rate of oil degradation, and the composition of microbial communities are influenced by ambient temperature [15]. The sea surface temperature affects the microbial colonization of spilled oil and the degradation of hydrocarbons in marine environments. The effects of temperature on the biodegradation of petroleum hydrocarbons in the marine environment are two folds: changing the physical properties of oil and affecting microbial physiology [15]. The composition and bioavailability of the spilled oil in seawater changes with temperature-dependent processes such as evaporation and dissolution, which are responsible for the disappearance of spilled oil [36]. Low temperatures can crystalize some components of oil, consequently protecting them from microbial attacks [37]. Temperature also affects microbial physiology, as the metabolic activities within microbial cells are temperature dependent. Enzymatic activities correlated with biodegradation of oil are reduced at low temperatures [38].
Temperature is one of the most important factors causing differential biodegradation of alkanes and PAHs. In microcosm experiments with South Louisiana crude oil and the dispersant Corexit 9500, PAHs were undetectable by day 11 in the absence of dispersant and by day 4 in its presence when incubated at 25 °C as shown in Figure 2 [37]. Alkanes are generally more amenable to biodegradation than PAHs [16][17][33]. At 5 °C, however, the rates of PAHs biodegradation were unusually higher than alkanes, which was explained to be due to crystallization of all above tridecane at temperature ≤5 °C, reducing biodegradation [37]. Additionally, bacterial communities that develop at low temperatures may prefer to degrade PAH over alkanes [16][39][40]. At low temperatures, alkanes may be less accessible to microbial populations than the more soluble PAHs [39][40].
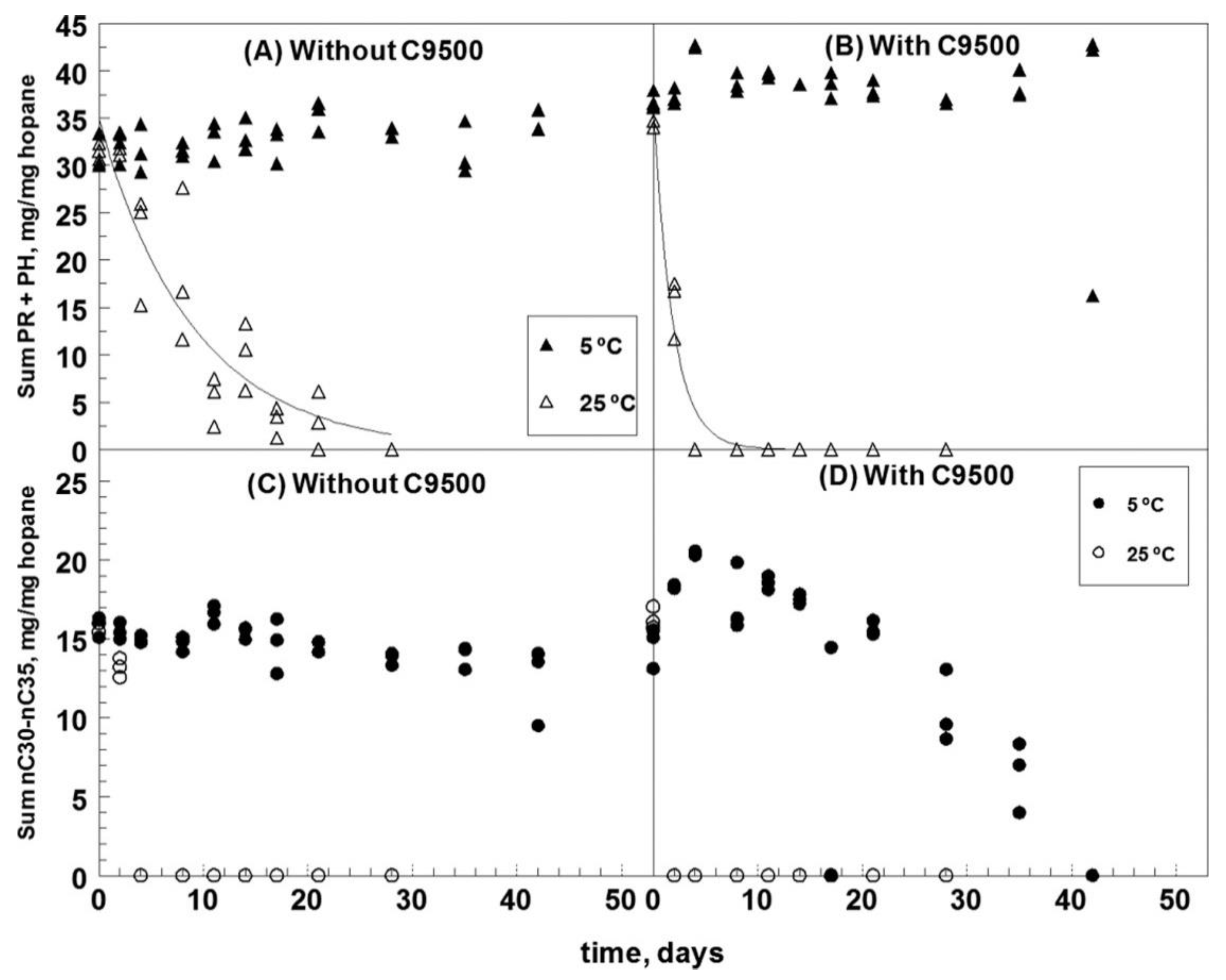 Figure 2. Biodegradation of the branched alkanes (A,B) and n-alkanes (C,D) > nC29 in the absence and presence of C9500 dispersant. Adapted from Campo et al. [37]. (A) Without C9500, (B) With C9500, (C) Without C9500, (D) With C9500.
Figure 2. Biodegradation of the branched alkanes (A,B) and n-alkanes (C,D) > nC29 in the absence and presence of C9500 dispersant. Adapted from Campo et al. [37]. (A) Without C9500, (B) With C9500, (C) Without C9500, (D) With C9500.Temperature also influences the mass transfer of substrate and/or electron acceptors in frozen ground, crucial to the cold-adapted microbes and consequent bioremediation [41][42]. In a study conducted on hydrocarbon bioremediation in simulated petroleum-polluted seawater collected from Tokyo Bay, it was found that the highest bioremediation efficiency was achieved in summer, followed by spring, autumn and winter respectively [43]. Moreover, heavy fuel (Bunker C) biodegradation by indigenous organisms in the North Sea was four times greater in summer (18 °C) than in winter (4 °C). For example, the temperature threshold for remarkable oil biodegradation is around 0 °C in the Arctic/sub-Arctic environments [44]. Although the microbial biodegradation activity does not cease at sub-zero temperatures, the process has not been thoroughly studied in the Arctic.
Among all environmental parameters, the most profound effects on bacterial communities are caused by temperature. Temperature affects the metabolic rates, including growth and survival, of microbes and determines the types of bacteria that can adapt to certain temperature ranges. The composition of the microbial community is affected by temperature as different types of bacteria are adapted to certain temperature ranges [39]. During the DWH oil spill, the fluctuation in temperature was the main leading factor in the bacterial community shifts from the deep-sea plume to surface slick [45][46]. For example, temperatures of 4 °C and 24 °C particularly select psychrophile and mesophile, respectively [47][48]. Temperature influences the activities of environmental communities, with some heterotrophic bacteria responding positively with increasing growth [49] and other smaller bacteria decreasing in growth [50].
Liu et al. [39] studied the deep and surface waters of the northern Gulf of Mexico for the roles of temperature, initial bacterial communities and nutrients in selecting oil degraders. The authors found that the development of microbial communities in high and low temperatures are different. The initial microbial community in the initial bottom water was primarily composed of α- and γ-Proteobacteria, Flavobacteria, Sphingobacteriia, Clostridia, and Opitutae. The microbial community shifted to mainly α-Proteobacteria (38 ± 15%) and γ-Proteobacteria (60 ± 16%) at 4 °C incubation to a less diverse community, which is dominated by Sulfitobacter. At 4 °C incubations, higher percentages of α-Proteobacteria (47 ± 17%) and lower percentages of γ-Proteobacteria (34 ± 19%) were found. Cycloclasticus, a known PAH degrader, dominated on day 50 at 4 °C but was nearly undetectable at 24 °C. The surface water had Cyanobacteria (60.4%), as the dominant bacterial community. At 4 °C incubation, Cyanobacteria was reduced to 10–30%. However, these Cyanobacteria were nearly undetectable at 24 °C. At 4 °C incubation, Sulfitobacter was the most dominant genus while at 24 °C, Oleibacter and Alteromonas predominated [39]. Total n-alkanes generally degrade faster at 24 °C, while PAHs decrease faster at 4 °C. Alkanes are generally more amenable to biodegradation than PAHs, however, n-alkanes above tridecane crystallize at ≤5 °C, thus, at a lower temperature, more soluble PAHs degrade faster. Moreover, the bacterial communities that develop at low temperatures may preferably degrade PAHs over alkanes.
2.2. Nutrients
Nutrients play a crucial role in the biodegradation of spilled oil [51]. The microbial activity is limited by the presence of inorganic nutrients such as nitrogen (N), phosphorus (P), and potassium (K), which are essential for microbial growth [52]. The availability of nutrients in seawater varies in different seasons, such as when nutrients are scarce in the fall and winter but abundant in the spring and summer [53]. Moreover, nitrate and phosphate are typically enriched in deep water, but depleted in surface water, and may regulate the development of oil-degrading bacterial communities [54][55][56].
Recent studies show that the addition of nutrients increases the degradation efficiency of oil. For example, Chen et al. [7] reported that a 10.0 g/L of crude oil-polluted water amended with 0.02 g/L and 2 g/L of nutrients showed a significant increase in degradation (76.9% and 155.4% increase. respectively) (Figure 3). Similarly, 0.01 g/L of oil amended with the same nutrients showed an increase of 20.4% and 24.0% [7]. Another study that used a combination of N and P observed a significant removal of 70% of lighter and 65% of heavier alkanes for just 1 h of incubation [57]. The addition of enough fertilizers that contain bioavailable nitrogen and phosphorus has been recorded to enhance oil biodegradation [27], increasing the hydrocarbon respiration rate [7][58]. The addition of N and P to marine sediments in Malaysia showed to stimulate alkane-degrading microbes, resulting in a decrease in n-alkane [59]. Laboratory results from the Gulf of Mexico show that the respiration rate in slick samples is five times higher than outside the slick, indicating that microorganisms in these oligotrophic waters quickly adjusted to breaking down hydrocarbon substrates [54]. This is due to inorganic nutrient amendments, which boost bacterial biomass and respiration.
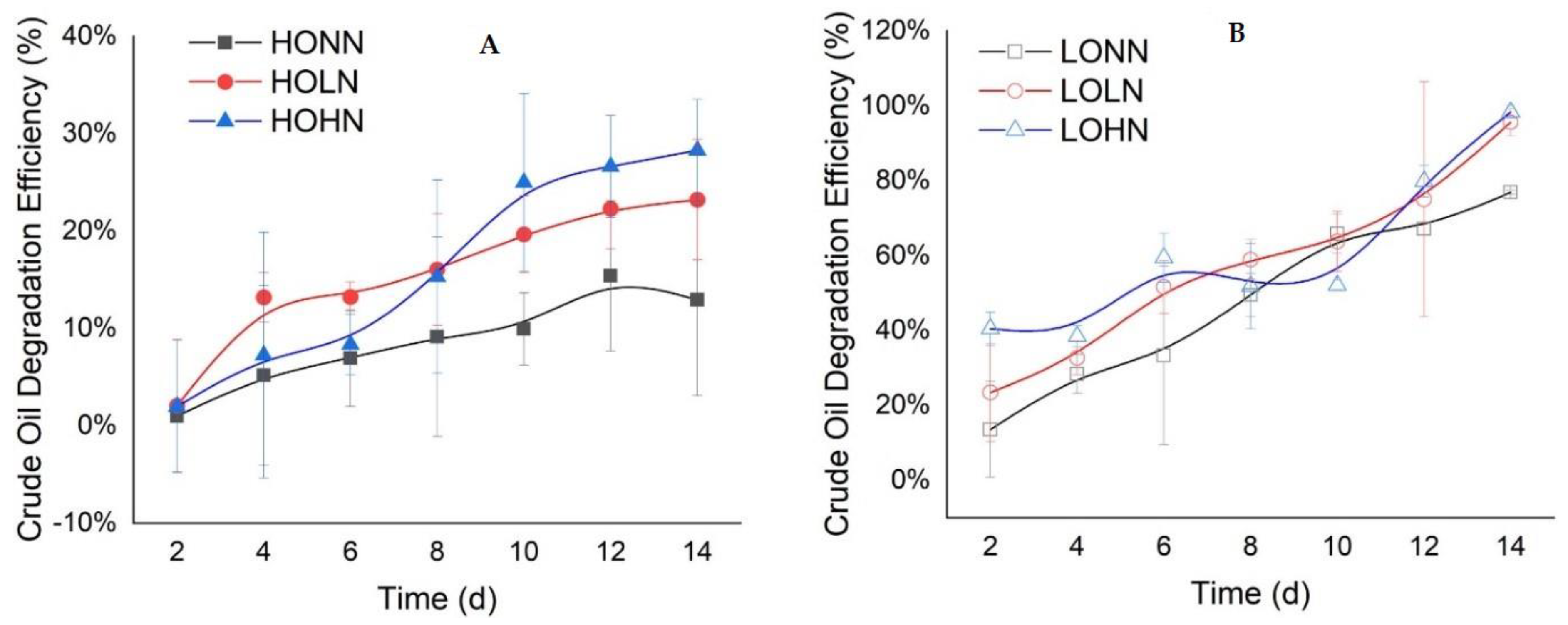 Figure 3. Biodegradation efficiency of crude oil with nutrient addition over 14 days of incubation. Three experiments with high oil concentration of oil 10 g/L are illustrated in (A), first with high oil concentration with no added nutrient (HONN), second with high oil concentration with low nutrients added (0.02 g/L of nutrient) (HOLN), and lastly, high oil concentration with high nutrients added (2 g/L of nutrients) (HOHN). Similarly, three experiments with a low oil concentration of 0.01 g/L of crude oil are illustrated in (B) low oil concentration with no nutrient added (LONN), low oil concentration with low nutrients added (0.02 g/L nutrients) (LOLN), and lastly, low oil concentration with high nutrients added (2 g/L nutrients) (LOHN). Adapted from Chen et al. [7].
Figure 3. Biodegradation efficiency of crude oil with nutrient addition over 14 days of incubation. Three experiments with high oil concentration of oil 10 g/L are illustrated in (A), first with high oil concentration with no added nutrient (HONN), second with high oil concentration with low nutrients added (0.02 g/L of nutrient) (HOLN), and lastly, high oil concentration with high nutrients added (2 g/L of nutrients) (HOHN). Similarly, three experiments with a low oil concentration of 0.01 g/L of crude oil are illustrated in (B) low oil concentration with no nutrient added (LONN), low oil concentration with low nutrients added (0.02 g/L nutrients) (LOLN), and lastly, low oil concentration with high nutrients added (2 g/L nutrients) (LOHN). Adapted from Chen et al. [7].However, other studies reported that the addition of more nutrients can inhibit biodegradation activity [60][61][62]. Having too many nutrients is considered to be toxic to cell growth, while the optimum concentration of fertilizers at 0.1% concentrations is best for the degradation of oil [63][64]. High concentrations of N, P. and K levels can contribute to the inhibition of biodegradation activity [65][66]. It is important to note that different microbial communities have different ratios of N/P optimum for the degradation of different hydrocarbons [67]. Excess nutrients can cause changes in lakes, rivers, and coastal ecosystems [68], resulting in unfavorable consequences like eutrophication [67]. Eutrophication can cause the reduction of biodiversity in both autotrophic and heterotrophic organisms [68], including biodegrading microorganisms.
Nearshore habitats are rich in microbial communities that provide services such as nutrient regeneration and the removal of pollutants [69][70]. These specific oil-degrading bacteria developed differently in rich nutrient and low nutrient environments from bottom water (nutrient-rich) to the surface water (nutrient-poor) environment. Alteromonas, Pseudoalteromonas, Oleibacter, and Winowgradskyella have been observed to grow better at bottom water-rich nutrient environments while Reinekea and Thalassobius develop better in surface water. [39]. Dave & Vya [52] found that the addition of nutrients beyond their maximum concentrations will have no significant effects, that adding phosphorus to three isolates M. albus, M. mesophicum, and N. otitdiscavairum at concentrations of 0.1% and 0.5% boosts growth and biodegradation rates, with a maximum of 0.5 concentration. Following the addition of nitrogen (concentrations from 0.1% to 1%), there was an increase in both biodegradation and growth rate on the three isolates. Lastly, potassium fertilizer addition at a maximum of 0.01% on the three isolates results in the decrease of degradation and growth rate [52].
During oil spills, it has been recorded that at least an estimated 150 g of N and 30 g of P are needed to be able to convert 1 kg of hydrocarbon to cell material [71]. An incubation of Galveston Bay seawater shows that the presence of oil tends to increase the abundance of heterotrophic bacteria regardless of nutrient concentration, but autotrophic microbial plankton decreased in abundance with the presence of oil (Figure 4). However, the addition of nutrients ameliorated the effects of oil on the autotrophic microbial community, such that it increased the abundance of all microbial groups in the nutrient treatments leading to a significant decrease in oil [31]. The addition of N in oil-polluted sites does not just increase the cell growth; it can also contribute to the decrease of lag phase and can result in maintaining a high microbial population [66][72].
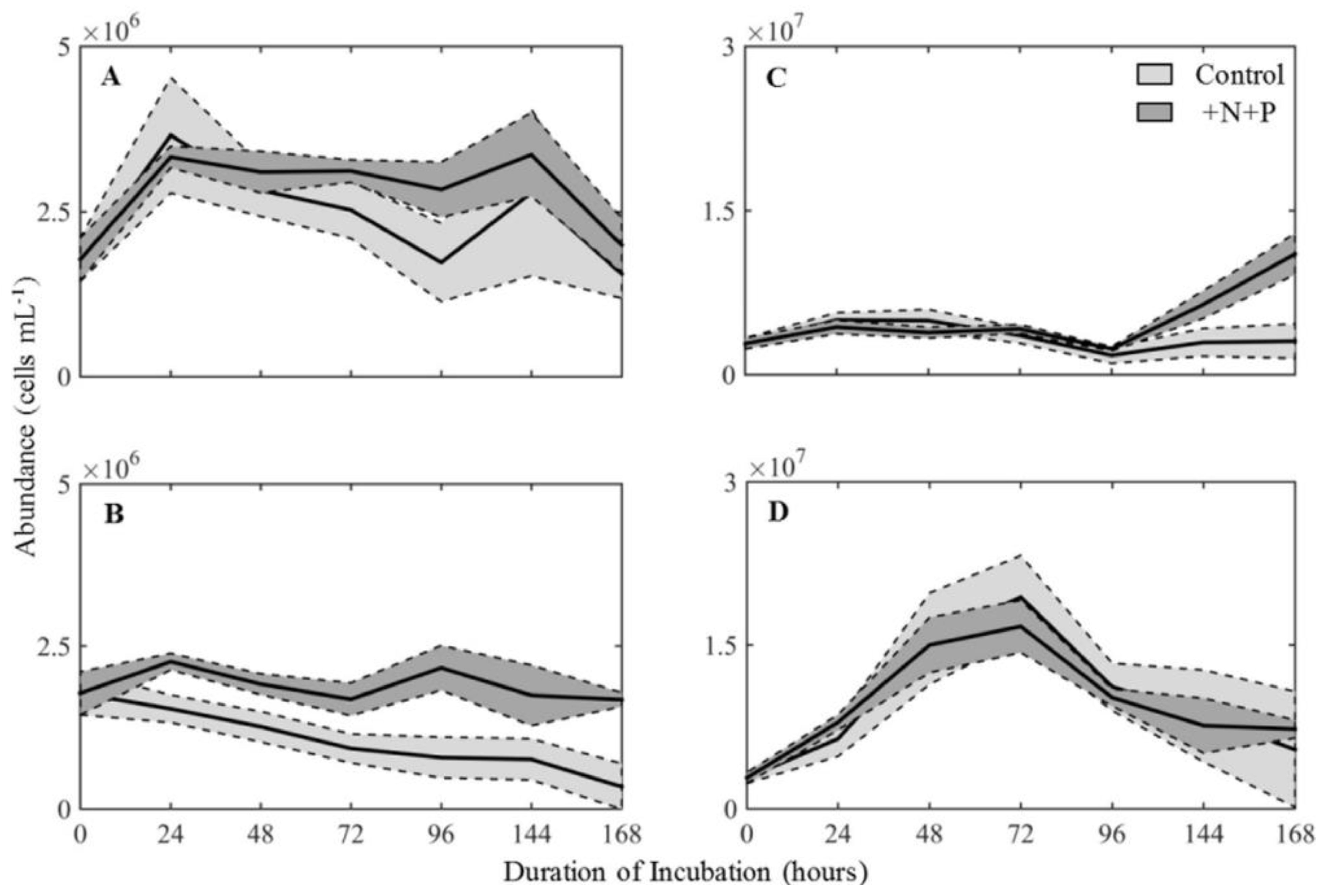 Figure 4. Comparing the microbial plankton abundance in incubated treatments over a total of 168 h, without nutrient enrichment (control) and with nitrogen and phosphorus (+N, +P). The black line is the mean value while the shaded represents standard deviation among the three replicates. The concentrations of autotrophic cells are illustrated in (A) for Station 2 and (B) for Station 3. As well, the concentrations of heterotrophic cells at Station 2 and Station 3 are illustrated in (C,D), respectively. Adapted from Williams et al. [31].
Figure 4. Comparing the microbial plankton abundance in incubated treatments over a total of 168 h, without nutrient enrichment (control) and with nitrogen and phosphorus (+N, +P). The black line is the mean value while the shaded represents standard deviation among the three replicates. The concentrations of autotrophic cells are illustrated in (A) for Station 2 and (B) for Station 3. As well, the concentrations of heterotrophic cells at Station 2 and Station 3 are illustrated in (C,D), respectively. Adapted from Williams et al. [31].2.3. Sunlight
Sunlight is an important factor affecting the degradation of crude oil and oil residues following an oil spill in marine environments. Specifically, photooxidation through the action of ultraviolet (UV) light from sunlight is one of the most important processes in the weathering of the spilled oil in surface water and is very effective at oxidizing 3 to 5 ring aromatics hydrocarbons in floating slicks and stranded oil [73][74]. Aromatics components of crude oils (e.g., PAHs) are the most photosensitive, followed by n-alkanes [75], although cyclic alkanes may decrease even faster than aromatic compounds [76].
There are two ways oil photooxidation can occur, direct and indirect photooxidation [77]. In direct photooxidation, compounds in crude oil such as PAHs are oxidized through direct absorption of sunlight. Indirect photooxidation happens when a wide range of reactive products are produced from crude oil during the absorption of sunlight such as singlet oxygen, peroxy radicals and hydroxyl radicals which oxidize other compounds in oil but not those that directly absorbs sunlight [77]. Thus, when the direct pathway is dominant, oxidation would be limited to a tiny fraction: only crude oil components that absorb light directly from the sun. Whereas, if the indirect photooxidation pathway is dominant, a much larger fraction of spilled oil would be vulnerable to oxidation [77].
Crude oil is typically exposed to photooxidation as a film or dissolved in surface waters known as the water-soluble fraction (WSF). The PAHs in WSF have exhibited a high degree of photooxidation, while PAHs in oil film have been shown to be less affected [78][79][80]. The greater the size and alkyl substitution, the higher is the sensitivity of aromatic compounds present in the crude oil to photooxidation. In the case of the DWH oil spill, it was suggested that the photooxidation pathway was indirect oxidation [81], such that hydroxyl radicals and singlet oxygen were formed [80]. It was reported that photooxidation was the main driver for degrading aromatic hydrocarbons, and biodegradation was the key process controlling the degradation of saturated hydrocarbons (Figure 5). Bacosa et al. [33] observed that photooxidation could degrade up to 40% of PAHs in 10 days and 90% of PAHs in 20 days. On both light and dark incubations, n-alkanes degrade faster in dispersed than in non-dispersed oil (Figure 5A). On the other hand, sunlight drove the degradation pattern of PAHs and their alkylated homologues (Figure 5B). Individual PAHs with lower molecular weight were more sensitive to photooxidation than PAHs with heavier molecular weight. After a long period of exposure to sunlight, about 60% of alkanes were degraded after 10 to 20 days with bacteria present [33]. Alkane degradation decreases as chain length increases. Also, specific types of alkanes (e.g., pristane and phytane) were almost completely depleted in the dark, an unexposed treatment, but has shown no significant hint of degradation in the light, exposed treatment even with the aid of a dispersant [33]. In general, unalkylated PAHs degrade slower than their alkylated homologues, and with the alkylation of each family, the removal of alkylated PAHs is increased [82].
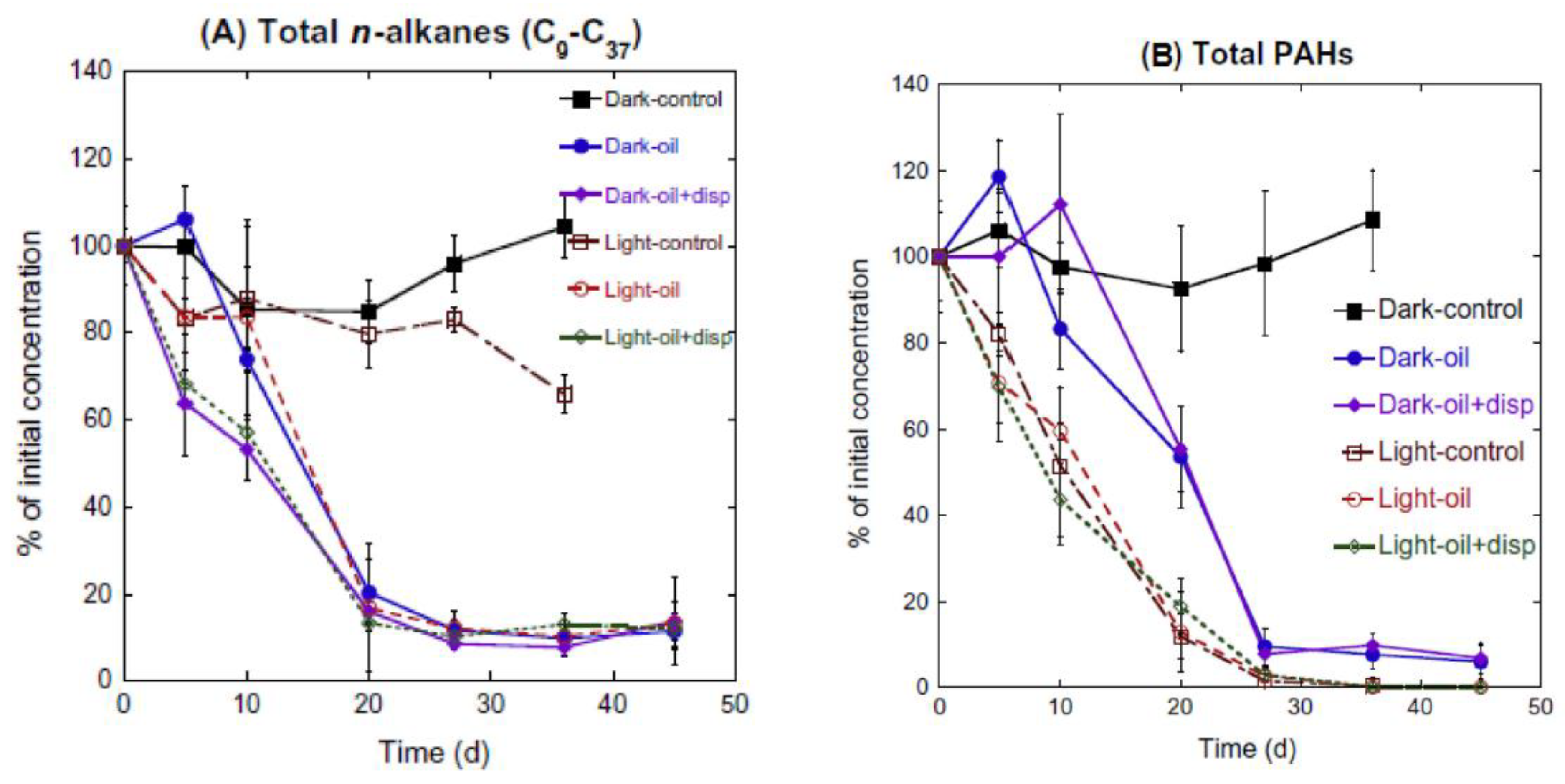 Figure 5. Degradation of total (A) n-alkanes and (B) PAHs under dark and light conditions. Adapted from Bacosa, et al. [33].
Figure 5. Degradation of total (A) n-alkanes and (B) PAHs under dark and light conditions. Adapted from Bacosa, et al. [33].Photooxidation can also increase the number of oil components susceptible to biodegradation, which is essential in the growth of oil-degrading bacterial communities [33][83]. In general, sunlight reduces the bacterial diversity and evenness in waters amended with crude oil alone, as well as with dispersed crude oil. Bacteria exposed to sunlight have been shown to be more dispersed with the change in oil chemistry compared to those that are unexposed to sunlight [33][84]. Sunlight has been shown to inhibit the growth of some bacterial species, but favor the growth of others (Figure 6) including Alteromonas, Marinobacter, Labrenzia, Sandarakinotalea, Bartonella, and Halomonas genera. On the other hand, crude oil-polluted water that has not been exposed to sunlight favored the growth and abundance of Thalassobius, Winogradskyella, Alcanivorax, Formosa, Pseudomonas, Eubacterium, Erythrobacter, Natronocella, and Coxiella [84]. Sunlight-favored bacteria were predominant after examining the mousse that is exposed to sunlight. This suggests that high surface temperature and/or strong sunlight played an important role in the growth and development of the bacterial community in oil-polluted waters.
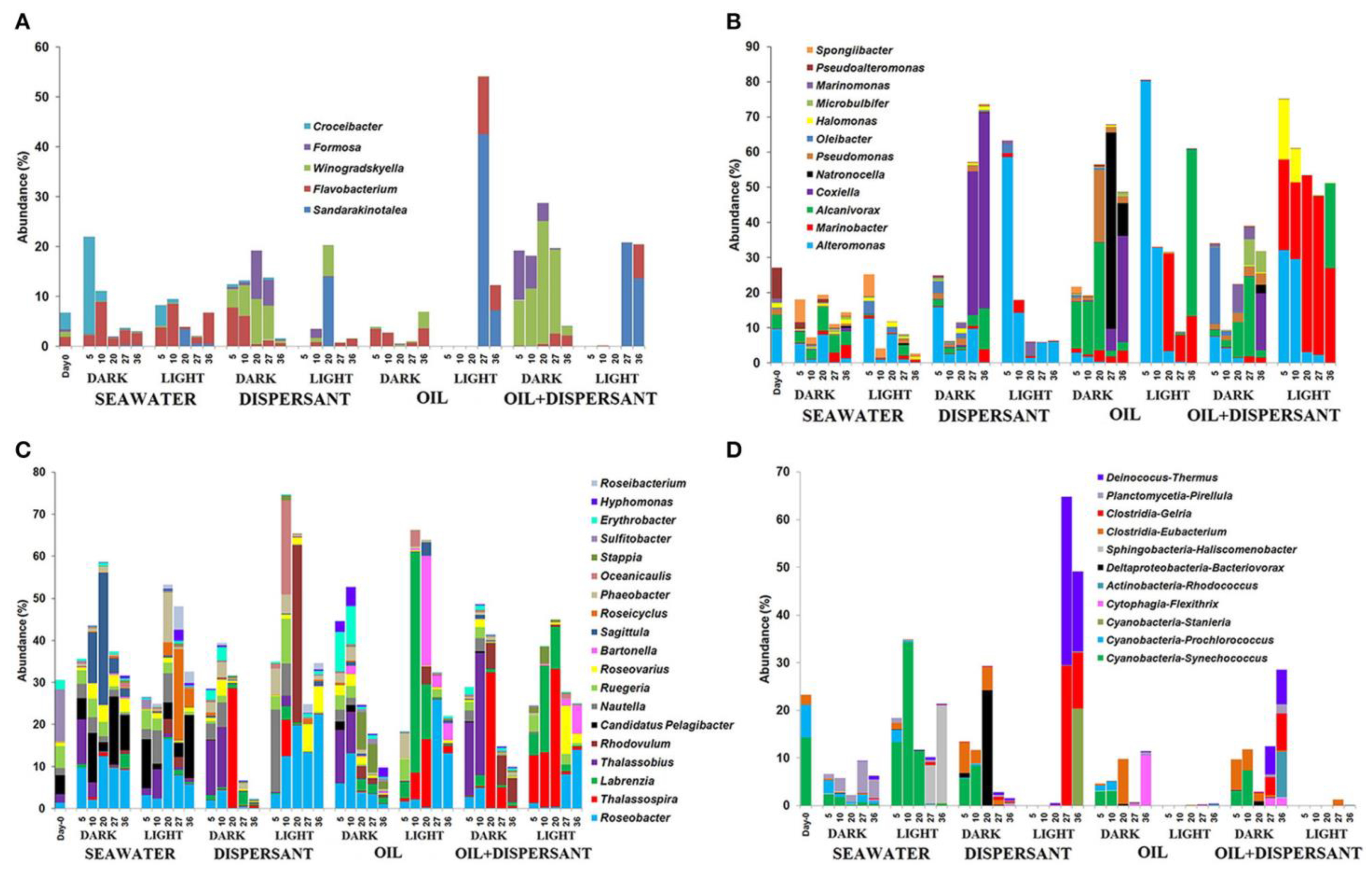 Figure 6. Abundances of bacteria at genus level in seawater alone, seawater with dispersant Corexit, crude oil, and both oil and dispersant incubated under the dark and light conditions for 36 days. Only abundances greater than 5% in at least one of the samples are shown. (A) Flavobacteria, (B) Gammaproteobacteria, (C) Alphaproteobacteria, (D) Other bacteria. The numbers indicate days of incubation. (Adapted from Bacosa et al. [84].)
Figure 6. Abundances of bacteria at genus level in seawater alone, seawater with dispersant Corexit, crude oil, and both oil and dispersant incubated under the dark and light conditions for 36 days. Only abundances greater than 5% in at least one of the samples are shown. (A) Flavobacteria, (B) Gammaproteobacteria, (C) Alphaproteobacteria, (D) Other bacteria. The numbers indicate days of incubation. (Adapted from Bacosa et al. [84].)Oil is an important source of hydroxyl radicals. A series of experiments have revealed that the total amount of singlet oxygen and hydroxyl radical produced in the irradiation of the thin oil films (100 mg) taken over the Gulf of Mexico were 2.0 × 10−5 and 3.7 × 10−7 moles, respectively [85][86]. Photo-produced singlet oxygen and hydroxyl radicals in steady-state concentrations were near 1 × 10−12 and 1.2 × 10−12 M in seawater under thin films of oil [85]. In the case of Macondo oil (DWH oil), it was observed that 90% of 350 nm radiation attenuated in the first 9 µm of the 60 µm film and that lower steady-state concentrations were produced by thicker oil than that of the thinner oil. This suggests why singlet oxygen formed at the very surface of the oil film were unlikely to diffuse through viscous oil to reach the aqueous layer [85]. Also, the Macondo oil produces higher triplet state energies than the natural dissolved oxygen matter [86], which can be explained by the relatively high concentrations of PAHs in oil [87].
When exposed to oil, bacteria release extracellular polymeric substances or EPS forming a microbially colonized matrix to which cells and other particles can attach [88][89]. EPS is a precursor of marine oil snow observed such as those during the Deepwater Horizon oil spill [90]. Marine snow can scavenge oil particularly heavy molecular PAHs [91]. EPS is produced as one of the microbial defense strategies against harmful or stressful conditions such as oil spills. EPS is mainly composed of protein and carbohydrates and different bacteria produce varying ratios of protein to carbohydrates [88][92]. Recently, strong evidence suggests that the stickiness of the EPS is due to its high protein content [93]. Sunlight plays an important role in EPS formation during an oil spill. In the presence of oil, natural sunlight stimulates polysaccharide secretion, coinciding with increased reactive oxygen production [89]. Moreover, oiled water irradiated with natural sunlight produces larger size aggregates suggesting that sunlight induce aggregation of bacterial EPS in marine environments [89][94].
2.4. Oxygen
Aerobic conditions are generally considered necessary for extensive degradation of oil hydrocarbons in the environment since major degradation pathways for petroleum hydrocarbons involve substrate oxidation by oxygenases in the catabolism of alkanes, cyclic, and aromatic hydrocarbons [95][96]. The initial intracellular attack of organic hydrocarbons is an oxidative process, whereby activation and incorporation of oxygen are catalyzed by oxygenases and peroxidases [13]. The aerobic bacterial catabolism of aromatic compounds involves a wide variety of peripheral pathways that activate structurally diverse substrates into a limited number of common intermediates, which are further cleaved and processed by a few central pathways to the central metabolism of the cell. Metabolic pathways and encoding genes responsible for the degradation of specific aromatic compounds have been characterized for many isolated bacterial strains, predominantly from Proteobacteria and Actinobacteria [97].
According to McNally et al. [98] and Grishchenkov et al. [99], the biodegradation of PAHs was slower in anaerobic conditions compared to aerobic conditions. Under aerobic conditions, the bulk of organic pollutants degrade at the fastest and most complete rate [100]. This has been demonstrated in research by Mille et al. [101] (Table 1), for example, who found that when the dissolved oxygen concentration in the overlying seawater was 8 ppm and the redox potential was around +150 mV, hydrocarbon degradation was noticeable. After reducing the dissolved oxygen concentration (2–3 ppm) and the redox potential (+30 mV), the degradation slowed down 2–3 times. Furthermore, there was no degradation under sub-oxic conditions (dissolved oxygen 0.2–0.3 ppm and redox potential −180 to −200 mV). Kostka et al. [102] suggested that the activation of hydrocarbon molecules during degradation is energetically demanding for microorganisms in the absence of oxygen, and thus anaerobic hydrocarbon degradation is much slower in comparison to aerobic metabolism. The supply of oxygen and major nutrients controls oil biodegradation.
Table 1. Percentage of biodegradation of spilled oil in sediments after a 60-day incubation. Adapted from Mille, et al. [101].
| Reactor | Redox Potential (mV) |
Biodegradation (%) | ||
|---|---|---|---|---|
| Extractable Organic Matter (EOM) | Saturated Fraction (FS) | Aromatic Fraction (FA) |
||
| 3 (8 ppm O2, stirring) | 150 | 64 | 70 | 59 |
| 4 (8 ppm O2, without stirring) | 90 | 55 | 61 | 52 |
| 5 (2–3 ppm O2) | 30 | 11 | 20 | 17 |
| 6 (0.2–0.3 ppm O2) | −190 | NS | NS | NS |
Beaches are dynamic systems where water and sediments mix easily are suitable for hydrocarbon degradation as they mostly have aerobic conditions [69][70]. However, in areas such as salt marshes where oxygen is easily depleted, a slower biodegradation rate is expected because anaerobic hydrocarbon degradation is slower than aerobic degradation [70]. According to Nicholson & Fathepure [103], the ability of halophilic and halotolerant bacteria to rapidly degrade benzene, toluene, ethylbenzene, and xylene (BTEX) compounds under aerobic conditions is high, and such activity could be enhanced markedly by the addition of growth-promoting nutrients such as yeast extract (YE).
A number of petroleum hydrocarbons (e.g., aliphatics and polyaromatics) have been demonstrated to utilize oxidizing processes in their breakdown pathways; however, these processes range substantially due to the varied oxygenases found in different bacterial species. Moreover, Alcanivorax borkumensis, a surfactant-producing marine bacterium [104], Cycloclasticus pugetii which uses aromatic compounds, including biphenyl, naphthalene, phenanthrene, anthracene, and toluene as principal carbon sources for growth [105] and Oleispira antarctica, a novel hydrocarbonoclastic marine bacterium isolated from Antarctic coastal seawater are specific bacteria known to degrade hydrocarbons under aerobic conditions [106].
2.5. Type of Spilled-Oil
Crude oils are generally categorized into light and heavy oils. Light oils, which yield gasoline, jet fuels, diesel and kerosene, typically consist of hydrocarbon chains ranging from C8–C20 which are less complex hydrocarbons and more easily degradable [107]. Heavy oils, which yield transportation fuels and act as a source of raw material for plastics, petrochemicals, lubricants, and road surfacing, are generally comprised of complex branched and ringed carbon chains > C20 so fewer microorganisms can degrade them and biodegradation rates are slower [107]. Previous research has proven that microbial degradation is a key step in deciding the destiny of spilled oil, which implies that microbial biodegradation is one of the primary mechanisms for removing non-volatile crude oil components from an oil-contaminated environment where bacteria that degrade hydrocarbons are prevalent [108]. Microbial biodegradation also has been found to occur naturally in the remediation of oil pollution linked with a number of oil spills [109][110]. In addition to that, the duration and essence of hydrocarbons in oil-derived compounds depart from C1–C10, as in gasoline [111]; C9–C16, as in jet fuel [112]; C10–C18, as in kerosene [113]; >C12, as in diesel fuel/oil [113]; and C20–C50, as in heavy Crude oil and waste oil/lubricating oil [113][114]. More complex structures (those with branches, higher hydrocarbon number, and/or condensed ring structures) are more difficult to biodegrade [107]. Moreover, in the case of PAHs, it has been observed that several genera of bacteria use lower-molecular-weight PAHs with two or three fused aromatic rings as their only source of carbon and energy [115].
Using 16S ribosomal DNA, a study characterized an isolate capable of degrading gasoline. It shared 100% sequence identity with a reference strain of Mycobacterium austroafricanum and degraded 86% of the substrate during growth on gasoline [116] (Table 2). It has been reported that the strain Mycobacterium austroafricanum, IFP 2173, was capable of degrading 3-methyl groups via a carboxylation and deacetylation process. Kerosene amended water was dominated by microbial communities of the genera Achromobacter, Alcaligenes, and Cupriavidus [16]. Degradation tests in sealed 120 mL vials of 1% kerosene showed that aromatic fractions deteriorated faster than aliphatic fractions [16]. Aromatic fractions, which are more toxic than aliphatic fractions, were preferred by the microbial consortium. Furthermore, in diesel incubations, Pseudomonas aeruginosa, Acinetobacter lwoffii, Corynebacterium sp. were found during incubation with a 1% concentration of the oil [117]. The identified bacterial consortium utilized all observable hydrocarbon peaks inherent in diesel within 14 days [117].
Table 2. Major classifications of spilled oil and corresponding valuable biodegrading bacteria.
| Oil | Carbon Composition | Oil Concentration | % of Oil Biodegraded | Dominant Bacteria |
Isolate or Consortium | References |
|---|---|---|---|---|---|---|
| Light oil | C4–C13 | 7% | 75% | Pseudomonas sp. | Isolate | [118] |
| Gasoline | C1–C10 | 10% | 86% | Mycobacterium austroafricanum | Isolate | [116] |
| Kerosene | C10–C18 | 1% | 20% | Achromobacter, Alcaligenes, Cupriavidus, Burkholderia, Rhodanobacter | Consortium | [16] |
| Diesel | >C12 | 1% | 85% | Pseudomonas aeruginosa, Acinetobacter sp., Corynebacterium sp. | Consortium | [119] |
| Engine oil/Lubricating oil | C9–C16 | 10% | 80–90% | Flavobacterium sp., Acinetobacterium calcoaceticum, and Pseudomonas aeruginosa | Consortium | [120] |
| Heavy oil | C20–C50 | 1% | 21–30% | Pseudomonas spp. Burkholderia spp. |
Consortium | [17] |
| Enterobacteriaceae | [121] |
In engine oils, based on biochemical tests and 16S rRNA sequencing, three predominant biodegrading isolates were identified as Flavobacterium sp., Acinetobacterium calcoaceticum, and Pseudomonas aeruginosa. The gravimetric study showed that A. calcoaceticum and a consortium of isolates were capable of using 80% and 90% of used engine oil, respectively, in a 4-week cycle under laboratory conditions at 30 °C and 160 rpm with Bushnell-Haas media [120]. An increase in oil degradation was associated with an increase in microbial cell number, which implies that the oil degradation was caused by the bacterial isolates. Over a two-week stretch, the three isolates Flavobacterium sp., Acinetobacterium calcoaceticum, and Pseudomonas aeruginosa, were capable of degrading up to 80% of the n-paraffin [120]. Heavy and extra-heavy crude oils, on the other hand, contain higher amounts of hazardous components [121]. A study by Darvishi, et al. [121] demonstrated that the isolated strain ERCPPI-1, Enterobactercloacea, which belonged to the genera Enterobactercloacea and Enterobacter sakazakii, can successfully degrade heavy crude oil components; maximal degradation was attained at a temperature of 40 °C and pH of 7.0. Similarly, Bacosa et al. [17] used surface sediment samples collected from Okinawa mangrove sites and supplemented with heavy oil as the sole carbon and energy source and reported that Pseudomonas or Burkholderia dominated the oil-degrading consortia. When incubated with hydrocarbon compounds, the active bacterial population moved to support Pseudomonas superiority. The K1 consortium was a dominant degrader, with the best potential to degrade aliphatic and aromatic hydrocarbon compounds; it could also degrade heavy oil at a concentration of 15% [17] (Table 2).
In addition to all of these, Zhou et al. [118] used Pseudomonas sp., a microbial strain, in an experiment to test the degradation rate of Diesel oil (LD) and heavy oil (HO). Figure 7A shows a trend in the effectiveness of LD degradation with different bacterial inoculums. When the bacteria dosage was 7 mL, the degrading efficiency of LD reached 75%, after which it could no longer be increased by increasing the bacteria dosage. Figure 7B shows the effectiveness of HO degradation by various bacterial inoculums. Heavy oil removal productivity achieved a limit of 94% when the bacteria dosage was 10 mL. In contrast, if too many inoculums were added, the growth and reproduction of bacteria might result in a competing flora, which would be detrimental to the use of LD and HO [122]. Moreover, in light and heavy oils (Figure 8), LD and HO had initial concentrations of 2209 mg/L and 1549 mg/L, respectively. After 24 h of cultivation, the degrading efficiencies of LD and HO were 39% and 28%, respectively. The high degradation efficiency of LD was due to two factors: (1) the component of LD was simpler than that of HO; and (2) the microorganism inoculation time in the LD mixture was shorter. Importantly, no single bacterial strain has the metabolic potential to degrade all of the compounds present in crude oil. Biodegradation of crude oil in nature usually requires a succession of organisms within the consortia of microbes present.
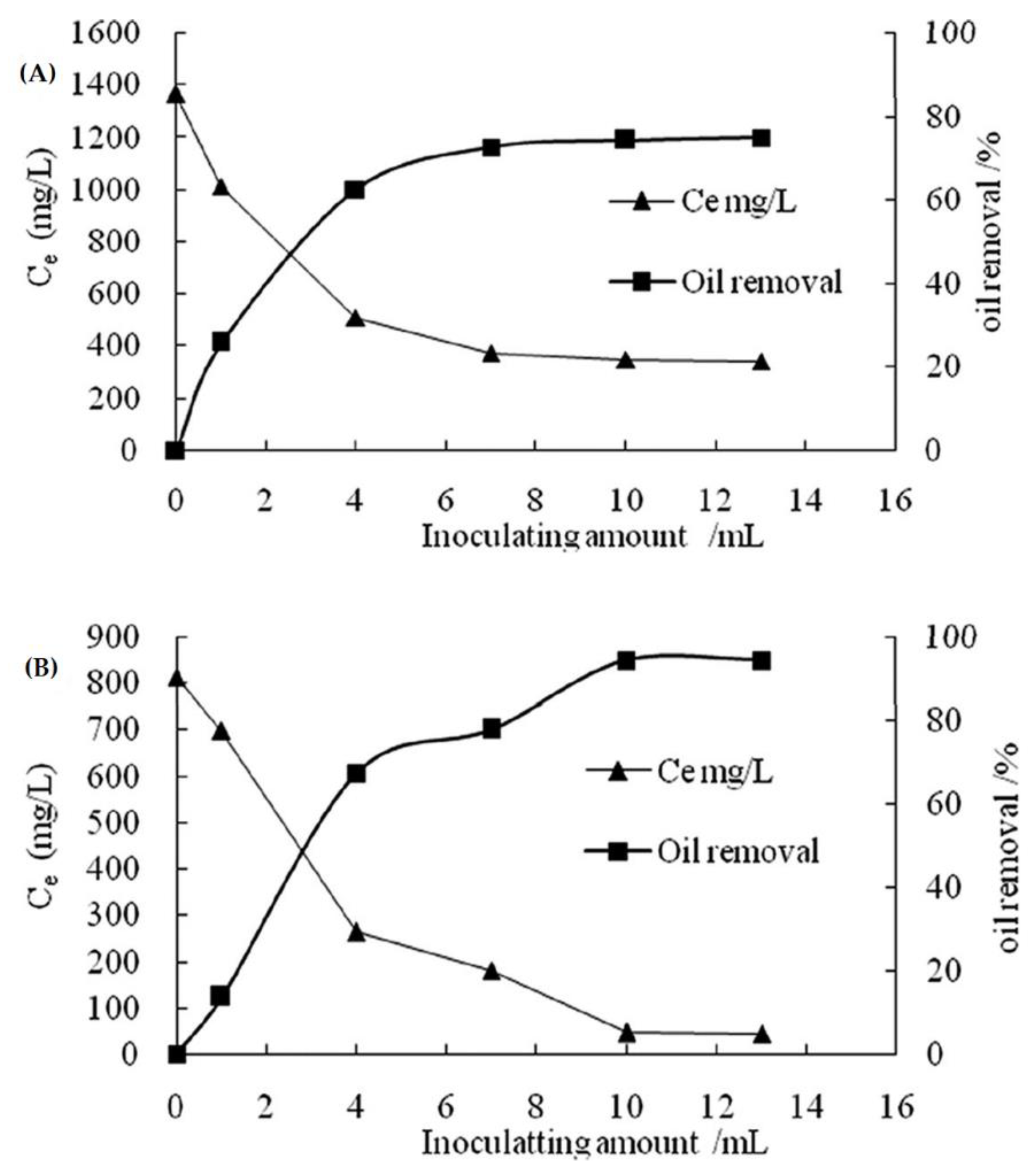 Figure 7. The impact of the number of bacteria inoculated on the biodegradation of Diesel Oil (LD) (A) and Heavy Oil (HO) (B). The x-axis (Ce) is the concentration of the residual oil after microbial degradation. Y-axis is the amount of inoculate per mL. Adapted from Zhou et al. [118].
Figure 7. The impact of the number of bacteria inoculated on the biodegradation of Diesel Oil (LD) (A) and Heavy Oil (HO) (B). The x-axis (Ce) is the concentration of the residual oil after microbial degradation. Y-axis is the amount of inoculate per mL. Adapted from Zhou et al. [118].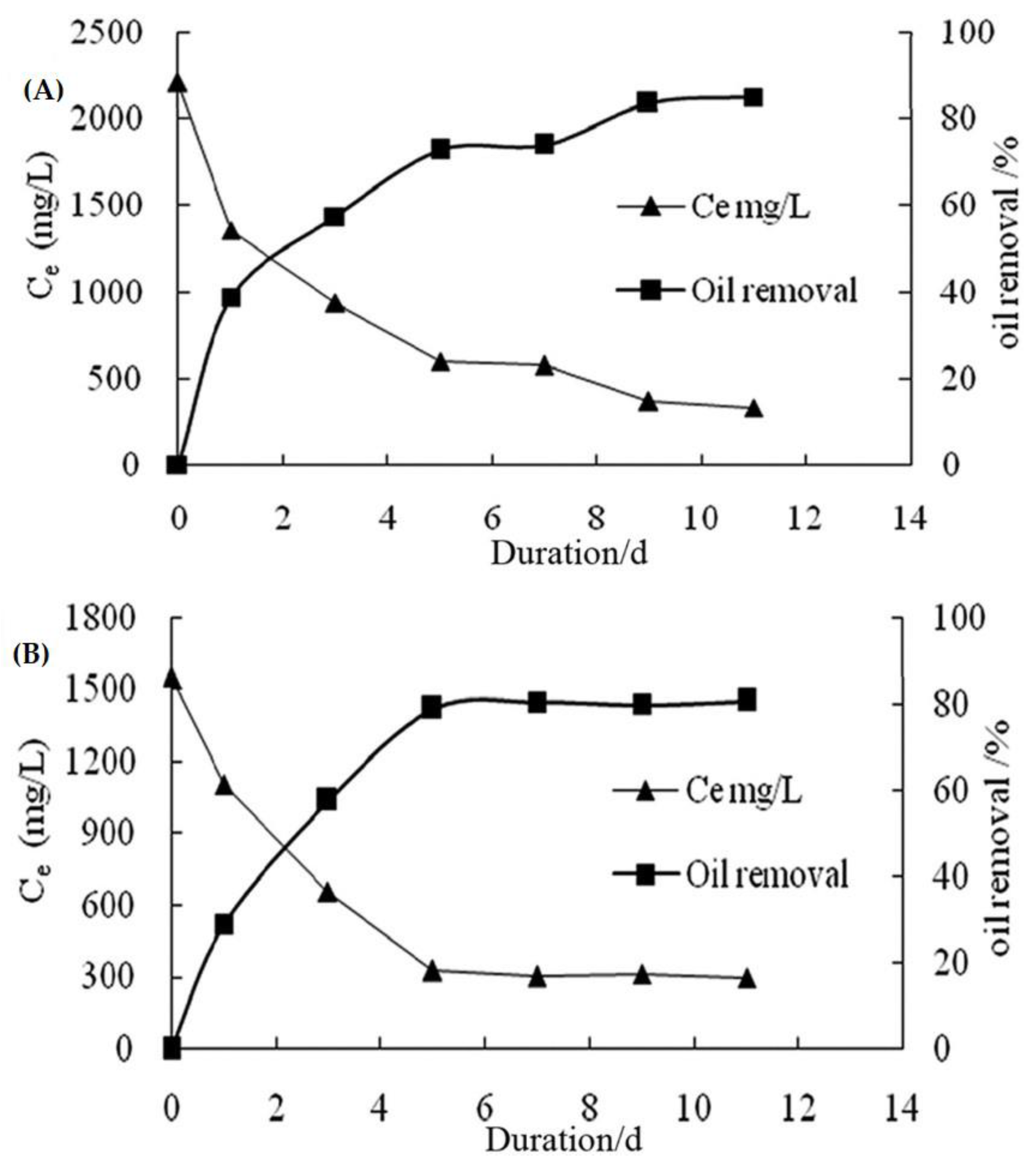
Figure 8. The biodegradation rate of the Diesel oil (LD) (A) and heavy oil (HO) (B) varies over time. x-axis (Ce) is the concentration of the residual oil after microbial degradation. Y-axis is the duration of incubation in days. Adapted from Zhou et al. [118].
2.6. Oil Concentration
Initial oil concentration affects the rate of oil biodegradation in the coastal-marine ecosystems. For example, in a study by Bacosa et al. [40], the alkane degradation rate constant was highest at 50 ppm (0.25 ppm/day) initial oil concentration, followed by 100 ppm and 200 ppm (0.13 ppm–0.15 ppm/day). The lowest degradation rate constants (0.05 ppm/day and 0.08 ppm/day) were observed in the high concentration 400 and 800 ppm treatments. The bacterial community structures at high oil concentrations differ from those at low concentrations [40] (Figure 9). As shown, Alcanivorax is abundant from 50 ppm to 200 ppm. However, it was barely detected at high concentrations (400 and 100 ppm). On the other hand, unclassified Rhodobacterales seem to tolerate high oil concentrations. It implies that some bacterial can adapt to stress caused by oil concentration and can metabolize hydrocarbons at such concentrated conditions better than the others.
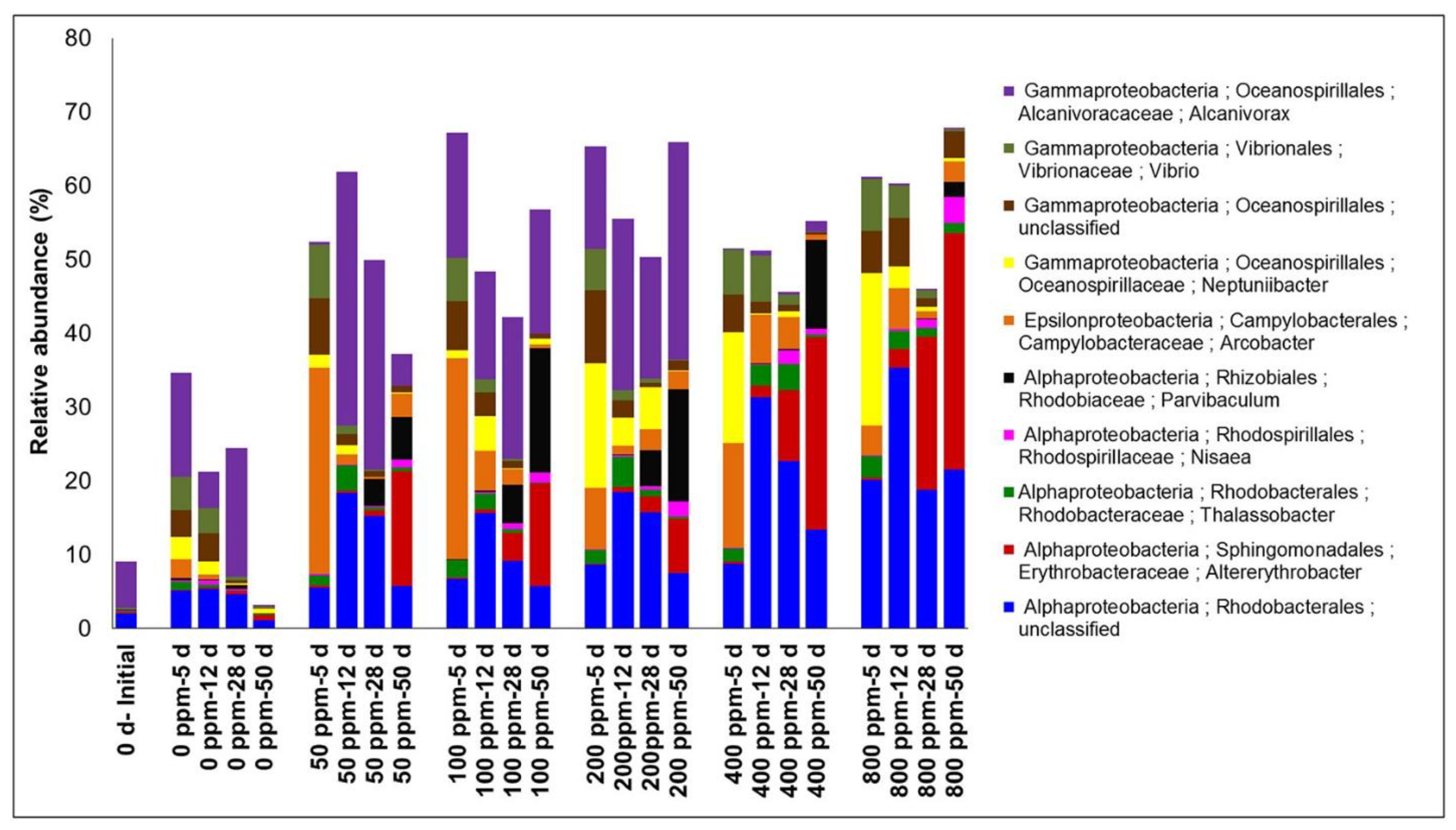 Figure 9. Relative abundances of potential biodegrading bacteria (at genus level) during the incubation of seawater with crude oil. Performed under controlled conditions of 0, 50, 100, 200, 4000 and 800 ppm oil in water and analyzed for initial community and 5, 12, 28 and 50 d of incubation. Adapted from Bacosa, et al. [40].
Figure 9. Relative abundances of potential biodegrading bacteria (at genus level) during the incubation of seawater with crude oil. Performed under controlled conditions of 0, 50, 100, 200, 4000 and 800 ppm oil in water and analyzed for initial community and 5, 12, 28 and 50 d of incubation. Adapted from Bacosa, et al. [40].In general, it has been shown that oil degradation decreases as oil concentration increases. However, various hydrocarbons such as alkanes and PAHs degrade at different rates. Oil concentration plays an important role in the determination of oil degraders in incubated surface saltwater. Based on the provided evidence about oil content, biodegradation studies (particularly when employing a closed system) must be considered. When exposed to distinct oil concentration gradients in a broader context, diverse bacteria may emerge in oil-polluted marine waters.
The abundance of certain phylotypes of bacteria may be associated with changes in oil chemistry. The alkane-degrading bacterium Alcanivorax and Arcobacter, a potential hydrocarbon-degrading bacterium, were prevalent at low concentrations [26][46]. Moreover, on higher concentrations, up to 800 ppm, alkane and aromatic hydrocarbon degraders have increased abundances such as unclassified Rhodobacterales, Altererytrobacter, and Neptuniibacter was observed). Thermogemmatisporaceae, Gemmataceae, and Thermodesulfovibrionaceae were shown to be connected with some medium- or long-chain alkanes in treatments with high alkane concentration [39]). Alkanes can be harmful at high doses and impede the growth of some microorganisms or beneficial to other bacteria. The structure of the microbial community can change as the dominant degraders of hydrocarbons or their metabolites outcompete others, which cannot break down at greater oil concentrations [123]. High oil concentrations inhibited Alcanivorax, due to the high concentration of oil, mainly alkanes and PAHs. Bacosa et al. [123] also reported that the biodegradation rates of alkanes, as determined by first-order kinetics, decreased with increasing oil concentration. The percent residual remained constant at 50% regardless of the initial engagement. Overall, Bacosa et al. [123] revealed that oil concentration significantly impacted hydrocarbon degradation and the bacterial community. Another study conducted by Rahman et al. [124] on the biodegradation of Bombay high crude oil using a mixed bacterial consortium showed that as the concentration of oil increases, the oil degradation rate decreases. Saturates, aromatics and polar chemicals found in various oil samples were all degraded to varying degrees by the same organisms [125]. A similar study conducted by Sathishkumar et al. [126] showed that the increase in crude oil concentration decreased the percent degradation; however, an increase in the quantity of natural oil degradation was still noticed.
2.7. Pressure
The bacterial community that biodegrades oil in the marine ecosystem can be influenced by pressure as bacterial communities can either be tolerant or sensitive to the changes in pressure [127]. In general, the deep ocean is considered to be an extreme environment for numerous forms of marine life (partially due to the high pressure), therefore their ability to perform certain metabolic tasks can be reduced [102]. An increase in pressure is commonly correlated to a decline in biodegradation [8]. This claim has been further proven by field and laboratory works, specifically on certain species of bacteria [128][129][130][131].
Piezophiles and piezo-tolerant organisms can tolerate an environment with extreme hydrostatic pressure [132]. Piezophilic bacteria can obtain their optimum growth at elevated hydrostatic pressures [133][134]. Some of the investigated oil-degrading bacterial communities do not commonly belong under this group because they lack certain flexible traits. Schedler et al. [128], found out that Rhodococcus qingshengii TUHH-12, a strain that can degrade alkane, grew best at a pressure of 15 MPa and its growth is slightly decreased at ambient pressure. In concert with the results of Schedler et al. [128]. Hackbusch et al. [131], who studied other Rhodococcus species, also observed that Rhodococcus sp. PC20 can tolerate pressures ranging from 0.1 MPa to 15 MPa. The growth of Rhodococcus sp. on n-hexadecane and crude oil at 0.1 and 15 MPa did not differ significantly (Figure 10). This strain clearly degrades a more saturated and aromatic fraction of oil at the deep-sea level (74.2% to 83.3%) than at an atmospheric pressure, where it can only degrade 60.5% to 71.2% of the oil components. Rhodococcus sp. PC20 can be classified as a piezo-tolerant organism because a pressure representing a deep-sea environment did not drastically hinder the ability of the bacteria to biodegrade aromatic hydrocarbons and saturates [131].
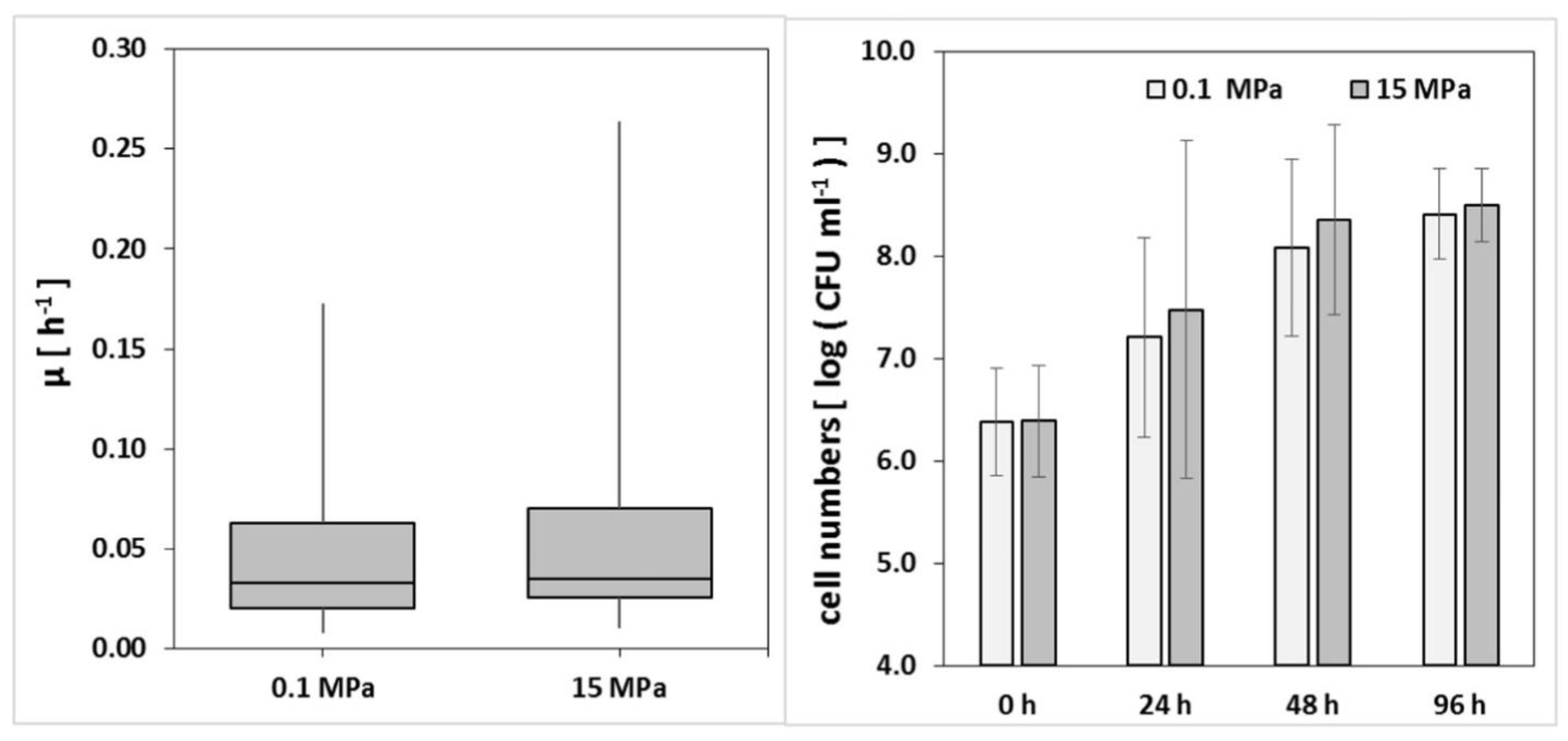 Figure 10. (Left): Boxplot of growth rates of all incubations of Rhodococcus sp. PC20 at 0.1 MPa and 15 MPa (22–192 h; n = 60 from 21 experiments). x-axis: amount of pressure in which the Rhodococcus sp. PC20 is exposed to; y-axis: average cell density of Rhodococcus sp. in atmospheric and elevated pressure (Right): Example of growth of Rhodococcus sp. PC20 at 0.1 and 15 MPa; 20 mL MM2 medium; 200 μL crude oil; 200 rpm; RT; in triplicates. 2 columns b/w. x-axis: incubation periods in hours; y-axis: Rhodococcus sp. PC20 cell densities on specific pressures. Adapted from Hackbusch et al. [131].
Figure 10. (Left): Boxplot of growth rates of all incubations of Rhodococcus sp. PC20 at 0.1 MPa and 15 MPa (22–192 h; n = 60 from 21 experiments). x-axis: amount of pressure in which the Rhodococcus sp. PC20 is exposed to; y-axis: average cell density of Rhodococcus sp. in atmospheric and elevated pressure (Right): Example of growth of Rhodococcus sp. PC20 at 0.1 and 15 MPa; 20 mL MM2 medium; 200 μL crude oil; 200 rpm; RT; in triplicates. 2 columns b/w. x-axis: incubation periods in hours; y-axis: Rhodococcus sp. PC20 cell densities on specific pressures. Adapted from Hackbusch et al. [131].In another study, Mason et al. [135] collected and analyzed samples of cells from the Gulf of Mexico four months after the DWH oil spill. The DWH oil spill occurred at ~1500 m below sea level, much deeper than previous oil spills. It exhibited an obvious shift to Photobacterium within the bacterial community when it was subjected to a pressure of 30 MPa. This clearly suggests that these bacteria can withstand notable increases in pressure [136]. Grossi et al. [137] also observed that Marinobacter hydrocarbonoclasticus grown in the presence of hexadecane has equivalent growth curves at both 0.1 and 35 MPa, but they are relatively slow. The γ-proteobacteria associated with oil biodegradation in coastal and deep-water ecosystems Colwellia and Pseudomonas, and the bacteroidetes Aequorivita, Flavobacterium, and Tenacibaculum were also found to be positively responsive to a high pressure of 30 MPa [138]. It was also found that the species under the genus Colwellia were one of the major oil-degrading bacteria following the DWH oil spill [135]. In general, piezo-tolerant organisms and piezophiles, especially the latter, can continuously undergo growth and have the ability to biodegrade oil due to their useful adaptation traits. Three common adaptation traits by these organisms include: (1) tuning the RNA transcripts that are common to piezophilic organisms; (2) expression of genes that are specified for piezophiles, and (3) molecular adaptation to high hydrostatic pressures [134].
On the other hand, piezosensitive organisms cannot tolerate high pressure [139]. Many, if not most, of the studied oil-degrading bacteria [129][138][140] belong to this group. Scoma et al. [141] discovered that a pressure of 5 and 10 MPa could significantly lower the growth yield of the strains Alcanivorax jadensis KS_339 and Alcanivorax dieselolei KS_293, especially the latter (Figure 11). The majority of the A. dieselolei KS_293 genome was already adversely impacted at mild pressure. In another laboratory experiment conducted by Scoma & Boon [129] where they studied other Alcanivorax species, they found out that A. borkumensis SK2 does not have enough of a resistance mechanism to help them improve their metabolism whenever they are exposed to increased hydrostatic pressure. This entry also showed that the growth of Alcanivorax species is mostly highly impacted by mild hydrostatic pressures, which further confirms their piezosensitive nature. Accounting for their piezosensitivity, this may be the major reason why they were not present among the hydrocarbon-degrading microbial communities found in the deep waters of the Gulf of Mexico following the DWH oil spill [138][142]. Sphingobium yanoikuyae B1, an aromatic hydrocarbon-degrading strain, was also highly inhibited by elevated pressures [128]. Thus, it can be deduced that species belonging to Alcanivorax have a limited role in the biodegradation of oil in deep-sea environments and that they are mostly useful for biodegradation in surface waters [129].
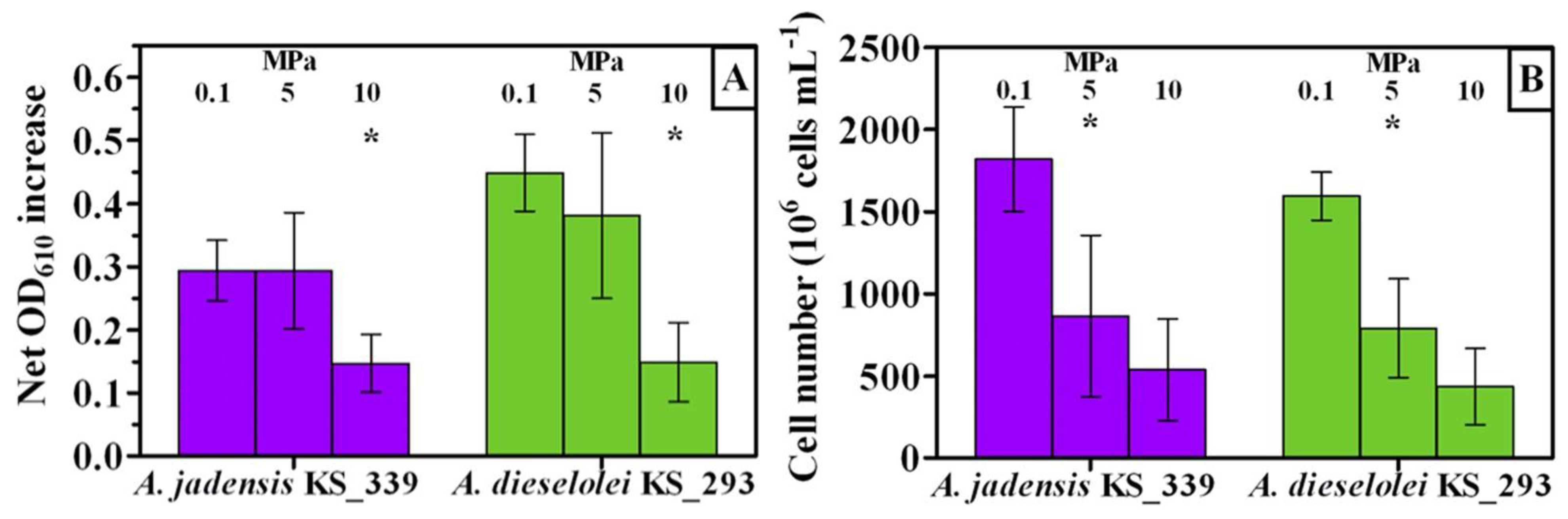 Figure 11. Growth of A. jadensis KS_339 (purple) and A. dieselolei KS_293 (green) under atmospheric (0.1 MPa) and mild pressure (5 and 10 MPa). Adapted from Scoma et al. [141] (Asterisks denote p-values: * < 0.05). (A): Net optical density increase. (B): Final cell number.
Figure 11. Growth of A. jadensis KS_339 (purple) and A. dieselolei KS_293 (green) under atmospheric (0.1 MPa) and mild pressure (5 and 10 MPa). Adapted from Scoma et al. [141] (Asterisks denote p-values: * < 0.05). (A): Net optical density increase. (B): Final cell number.According to the study of Prince et al. [143], the crude oil added to the seawater sample collected from Log Bay at 8 m deep was degraded faster by the indigenous microbes at atmospheric pressure than at 15 MPa (i.e., both at 5 °C), although minimal, a difference in the rate was observed. Moreover, the half-life of the total detectable hydrocarbons was obtained after 13 days at atmospheric pressure but it took 16 days at a pressure of 15 MPa [143]. Calderon et al. [138] also observed a much higher relative abundance of the genera Halomonas, Marinobacter, Amphritea Pseudophaeobacter and Aestuariicella at a pressure of 0.1 MPa from the samples collected from the Hatton–Rockall Basin at 2900 m deep. Even though Halomonas can survive in both deep and shallow waters, these authors deduced that this genus can biodegrade hydrocarbons better in shallow waters [138]. With these available studies, it can be concluded that certain bacteria are adapted to their in situ environment [102]. Furthermore, the degradation of hydrocarbons is only moderately affected at a pressure of 10 to 15 MPa, but high pressures approaching 50 MPa have a more drastic effect [102]. In contrast, Aulenta et al. [144] claimed that a hydrostatic pressure of 10 MPa can already severely impact oil biodegradation.
Further, low temperature and high pressure can act synergistically to slacken the biodegradation of hydrocarbons [102][143][145]. Louvado et al. [146] conveyed that high hydrostatic pressure, side by side with low temperature, can reduce PAHs biodegradation because of the reduction in its permeability. This inhibits the PAHs uptake, which is essential for the intracellular initial dioxygenation reaction and the alteration of PAH-dioxygenase supramolecular conformation. Catalytic sites may also inactivate essential enzymes [146].
2.8. Shoreline Energy
Hydrography of the spill site (e.g., shoreline energy) can affect petroleum hydrocarbon weathering, especially for spilled oil reaching the coastal zone where shoreline energy (i.e., wave activity/intensity) can plan an important role in oil weathering [5][147]. Historically, the effects of shoreline phenomena on transport and fates of spilled oil have been studied following major oil spills. Following the Exxon Valdez spill in Alaska and Gulf War spill along the Saudi Arabian Coast, many researchers documented the entrainment of oil in protected shorelines [148][149][150]. Researchers have also summarized how variable combinations of low shoreline energy and protected coastal geography have led to multi-decadal persistence of petroleum following various oil spills [151], such as the Amoco Cadiz spill [152], the Gulf War spill [149], the Exxon Valdez spill [148]. In addition, an exposure index for predicting oil spill damage on shorelines as a function of shoreline geography and wave energy has been pioneered [153].
The shoreline energy increases the surface area of the oil as well as exchange between oil, water, microbes, oxygen, and nutrients [154]. Oil degrades most rapidly where these are greatest [154]. The most obvious driver for advanced degradation with higher shoreline energy is physical advection, which breaks apart oil droplets; this creates a greater surface area for dissolution of hydrocarbons into the surrounding water, enhances evaporation of volatile hydrocarbons into the air, and increases surface area and dissolved oxygen content for bacteria to break down petroleum hydrocarbons [151][155]. For instance, Evans et al. [147] showed that fewer hydrocarbons persisted in high energy environments than low energy environments following DWH, a pattern consistently observed in contaminated coastlines after Amoco Cadiz, Exxon Valdez, and Gulf War oil spills [148][149][152]. This is likely due to advanced dispersion from wave action, which enhances dissolution and increases surface area for biodegradation [151][155].
On the other hand, predicting the dispersion of oil droplets in the water column is one of the major factors that needs further assessment in order to determine the impacts of spilled oil on the ecosystem and communities. For oil released on the ocean surface, breaking waves play an important role in the dispersion and fate of oil spills [156]. As studied by Tkalich and Chan [157] higher energy waves trigger more intensive breaking of the slick and dispersion. Evans et al. (2017) expanded the research on the effect of shoreline energy on spilled oil degradation by analyzing the compound-specific weathering patterns of the samples collected by Pendergraft and Rosenheim [154]. After source confirmation using biomarker ratios, they quantified the n-alkane, PAH and alkylated PAH contents of these samples. According to Evans et al. [147], this is an important expansion from the previous work of Pendergraft and Rosenheim [154] as toxicity and bioavailability are determined on a compound-specific basis, whereas Pendergraft and Rosenheim’s technique is useful to confirm the presence of labile versus recalcitrant hydrocarbons in general. Across the three locations analyzed, samples collected at the same time showed the highest specific hydrocarbon content in an enclosed marsh, followed by a protected beach and finally, a high energy, oceanfront beach. Figure 12 provides an example of this pattern observed between samples collected at different locations between 678 and 694 days following the spill for the sum of n-alkanes C10 with 37 and the 16 priority PAHs by the United States Environmental Protection Agency (EPA). Weathering patterns across the time also confirmed that early samples were least weathered and older samples were most weathered, as evidenced by the highest hydrocarbon concentrations in oil sheens and early oiled sediment. Importantly, at any one time, the most extensive weathering took place in high energy environments.
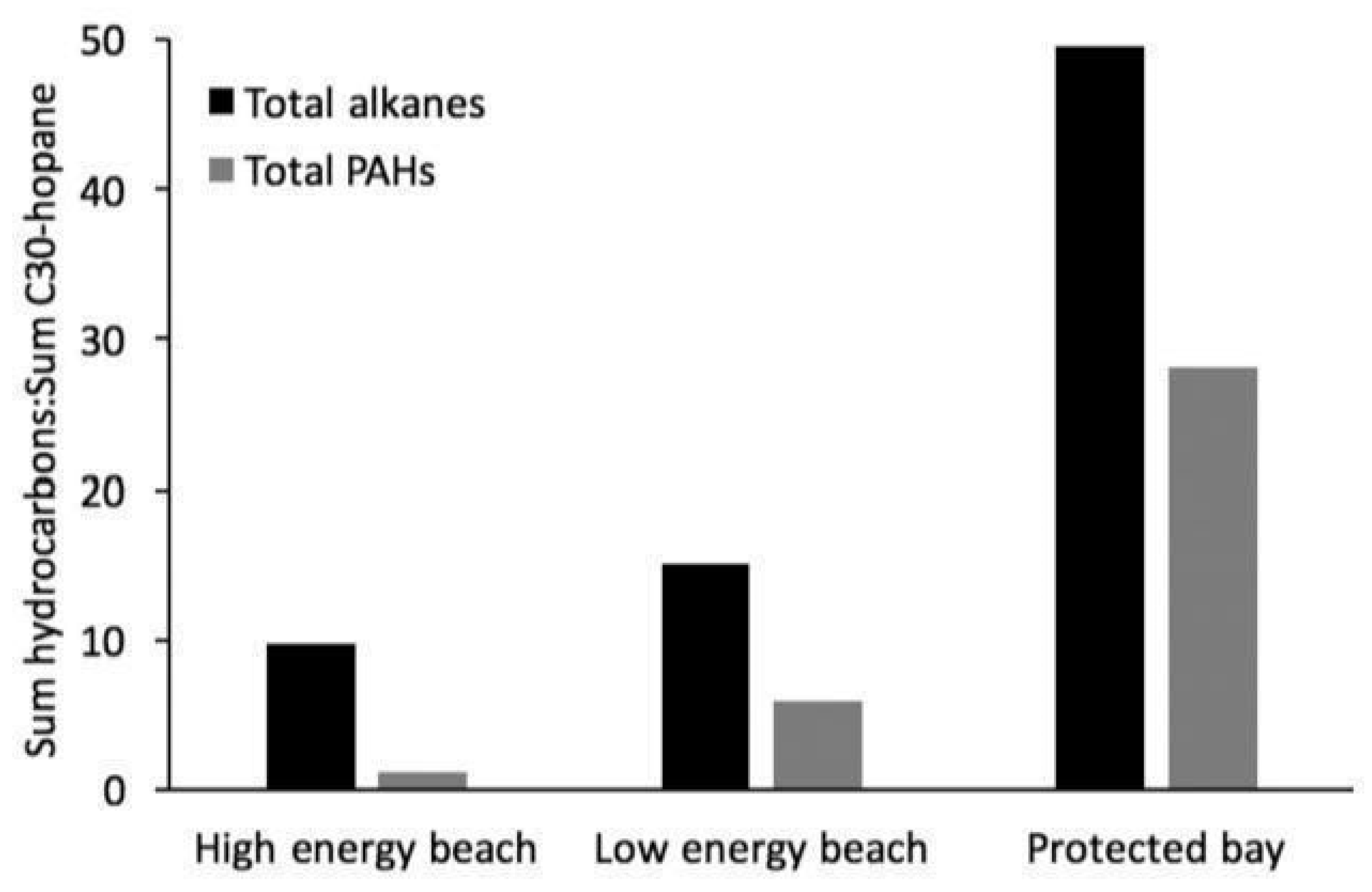 Figure 12. The sum of n-alkanes C10-37 and the 16 EPA listed priority PAHs, normalized to internal recalcitrant biomarker C30-hopane, are presented for three samples collected between 678 and 694 days following the Deepwater Horizon oil spill. For both hydrocarbon categories, content is highest in a protected bay sediment sample and lowest in a high energy beach sediment sample, illustrating the effect of shoreline energy on hydrocarbon weathering. Adapted from Bacosa et al. [5].
Figure 12. The sum of n-alkanes C10-37 and the 16 EPA listed priority PAHs, normalized to internal recalcitrant biomarker C30-hopane, are presented for three samples collected between 678 and 694 days following the Deepwater Horizon oil spill. For both hydrocarbon categories, content is highest in a protected bay sediment sample and lowest in a high energy beach sediment sample, illustrating the effect of shoreline energy on hydrocarbon weathering. Adapted from Bacosa et al. [5].2.9. Bacterial Community
The initial bacterial community composition at the spill site is one of the most important factors affecting the extent of biodegradation. In the natural environment, it has been observed that the initial microbial community can contribute to up to 19% of the oil-degrading bacterial community [39]. Initial bacterial communities prior to an oil spill, in particular the naturally present oil-degrading microorganisms, may be important in controlling the development of oil-degrading communities [158]. Recent studies have identified bacteria from more than 79 genera that have the capabilities to degrade petroleum hydrocarbons [159]. Several of these bacteria are Oceanospirillales, Rhodobacterales, Altererythrobacter, Neptuniibacter, Colwellia, Cycloclasticus, Pseudoalteromonas, Actinobacteria, Pseudomonas, Bacillus, Burkholderia, Enterobacter, Kocuria, Pandoraea, Corynebacterium [29][40][45][55][126]. Some bacteria including Alcanivorax, Marinobacter, Thallassolituus, Cycloclasticus, Oleispira, and a few other OHCB (obligate hydrocarbonoclastic bacteria), showed a low abundance or undetectable status before pollution but were dominant after petroleum oil contamination [106]. These occurrences suggest that these microorganisms significantly influence the transformation and fate of petroleum hydrocarbons in the environment.
Although researchers may generally refer to bacterial taxa as oil-degrading, most bacteria can only successfully degrade or utilize specific petroleum hydrocarbons in oil residues [160][161]. Different bacteria have different petroleum hydrocarbon degradation abilities because these bacteria have other catalytic enzymes, thus, their role in degrading petroleum hydrocarbons varies widely [162]. Bacterial cultures can metabolize only a limited range of hydrocarbons [163], which denotes that different bacteria exhibit different oil degradation capabilities. Moreover, even bacteria belonging to the same species have different hydrocarbon-degrading capabilities [164]. No single bacterial species can potentially degrade oil completely. These results suggest that establishing an extensive hydrocarbon-degrading microorganism consortium is a crucial factor for the bioremediation of spilled oils [165].
Generally, the microbial consortium has a higher degradation ability than bacterial isolates because a microbial consortium comprises different microbes with different characteristics and skills, thus it can degrade various components of oils [164]. Different bacteria can complement each other in a consortium, for example, through co-metabolic turnover events or interactions with substrates (e.g., via biofilm formation), resulting in a more effective degradation process [163]. Pelz et al. [166] demonstrated that primary hydrocarbon degraders’ metabolic and physiological limitations could be efficiently compensated in a consortium by the recruitment of other species from the mixed culture with similar physiology. Another benefit of a bacterial consortium is its ability to degrade the parent molecule into carbon dioxide and water, which is impossible to do using chemical or physical remediation procedures [16][164]. While a single strain can be incubated with a substrate, the interplay between microbial consortium members is far more complicated. Some consortium members make the oxygenases and dioxygenases needed to oxidize the hydrocarbon chain, while others make the micronutrients required by the primary degraders [20][28].
A number of published studies illustrate the importance of bacterial consortia in degradation. Rahman et al. [124] reported that individual bacterial cultures show less growth and degradation than the mixed bacterial consortium. The study showed that at 1% crude oil concentration, the mixed bacterial consortium degraded 78% of the Bombay High (BH) crude oil (Figure 13). Individual bacteria also showed degradation abilities, wherein Pseudomonas sp., specifically Pseudomonas sp. DS10-129 (66%) obtained the highest degradation rate, followed by Bacillus sp. DS6-86 (59%), Micrococcus sp. GS2-22 (49%), Corynebacterium sp. GS5-66 (43%), and Flavobacterium sp. DS5-73 (41%). The figure below shows the results of a study wherein different bacterial isolates and a mixed bacterial consortium were tested in different oil concentrations.
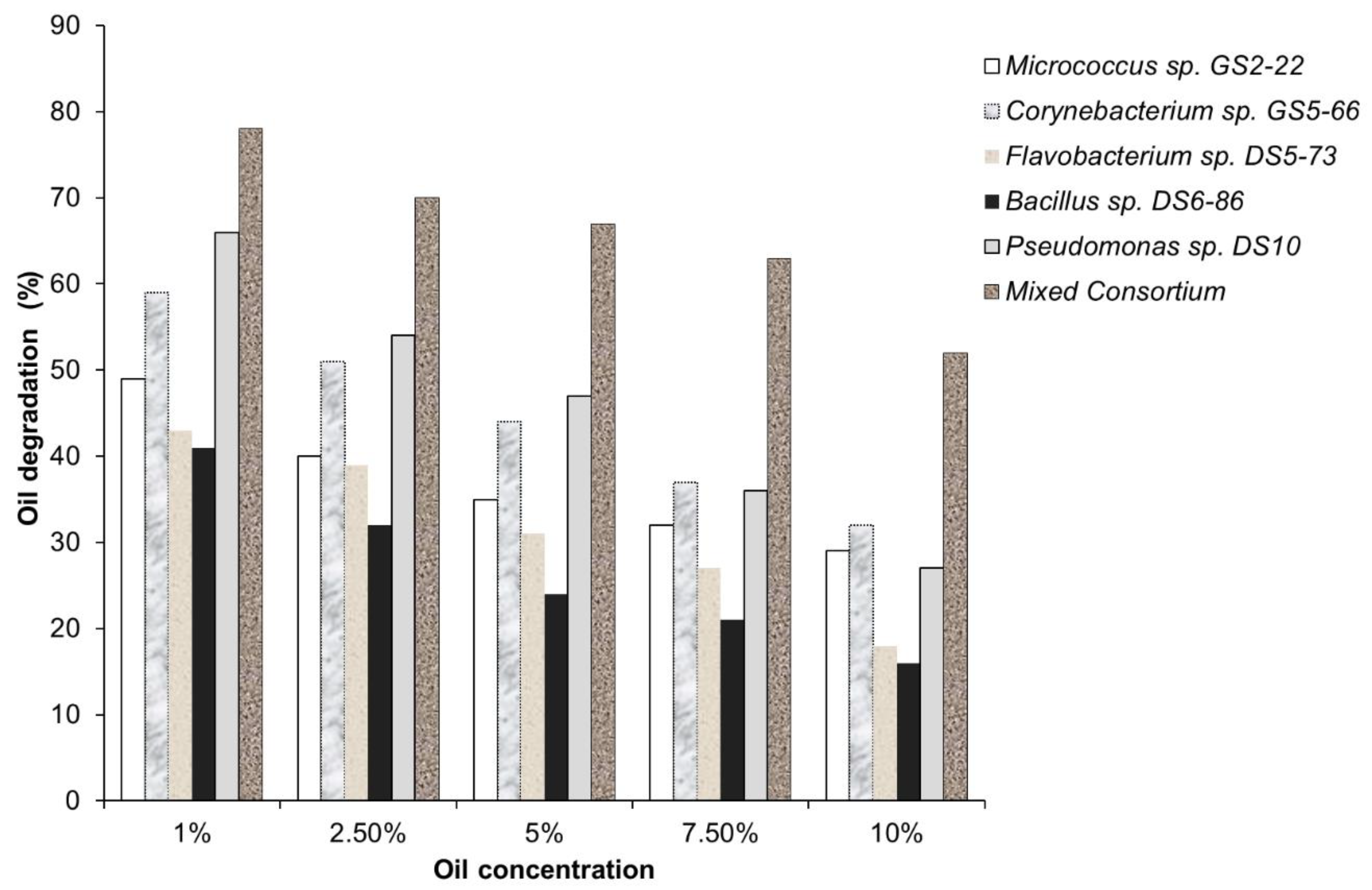 Figure 13. Biodegradation of BH crude oil by pure bacterial cultures and mixed bacterial consortium in different oil concentrations (%) after 20 days of incubation. Modified from Rahman, et al. [124].
Figure 13. Biodegradation of BH crude oil by pure bacterial cultures and mixed bacterial consortium in different oil concentrations (%) after 20 days of incubation. Modified from Rahman, et al. [124].The results showed that as the concentration of oil increases, the oil degradation rate of both the individual bacterial isolates and the bacterial consortium decreases. Moreover, it can be observed that the mixed bacterial consortium obtained the highest oil degradation rate in all the different oil concentrations, which implies that the consortium was able to degrade hydrocarbons more than the individual bacterial isolates. In addition, Pseudomonas sp. DS10 showed the highest degradation rate next to the mixed bacterial consortium. Similar results were observed in another study as reported by Sathiskumar et al. [126], where the mixed bacterial consortium showed more growth and degradation than the individual strains. In the study, at 1% crude oil, the mixed bacterial consortium degraded a maximum of 77% of the crude oil. The bacteria that showed the highest degradation ability in this entry is the Pseudomonas sp., specifically Pseudomonas sp. BPS1-8 then followed by Bacillus sp. IOS1-7 with 64% degradation rate, Pseudomonas sp. HPS2-5 with 45%, and Corynebacterium sp. BPS2-6 with 41%. Similar results were reported by Bacosa & Inoue [164] wherein the bacterial consortium namely T2, T1, T3, O, K1, and K2 has higher degradation rates compared to the bacterial isolates T2B (Pseudomonas sp.) and T2C (Burkholderia sp.). The isolates T2B and T2C were able to utilize various aliphatic and aromatic hydrocarbons but were unable to outperform the source consortium in terms of degradation. All bacterial consortia examined in the study were able to degrade heavy oil more efficiently than both isolates (Pseudomonas sp. and Burkholderia sp.) which indicates that the consortium is more superior in degrading heavy oil than any of the isolates.
Bioremediation is considered to be a superior approach since it effectively destroys the hydrocarbons and prevents the contaminants from accumulating. Studies revealed that different bacteria indeed exhibit different oil degradation capabilities, and no single bacterial species can potentially degrade oil completely. However, it can be suggested that under optimized conditions, a mixed bacterial consortium can be an effective and eco-friendly solution for oil degradation as it showed a higher degradation rate than bacterial isolates [124][126][164]. Moreover, individual isolates such as Pseudomonas sp. and Burkholderia sp., despite their lower degradation capabilities compared to the source consortia, can be useful to create a powerful and stable bacterial consortium for more effective cleanup of contaminated locations.
2.10. Mineral Particles
Suspended mineral particles affect the course of an oil spill in aquatic environments. It can mix naturally with oil in turbulent waters, and when this happens, oil-particle aggregates (OPA) are formed [167][168] (Figure 14). This can be viewed as a natural cleansing mechanism for removing oil residues from the marine water column by flocculation [169][170]. Flocculation happens when micron-sized mineral particles interact with oil droplets in the presence of water-containing ions [171]. The interaction reduces the adhesion of oil to solid surfaces because once flocculated, the oil droplets do not coalesce [172]. In addition, the adhesion of oil to the shoreline sediments is greatly reduced. Hence, the interaction of oil and mineral particles makes the dispersion and removal of oil droplets by the waves and tidal current easier [171].
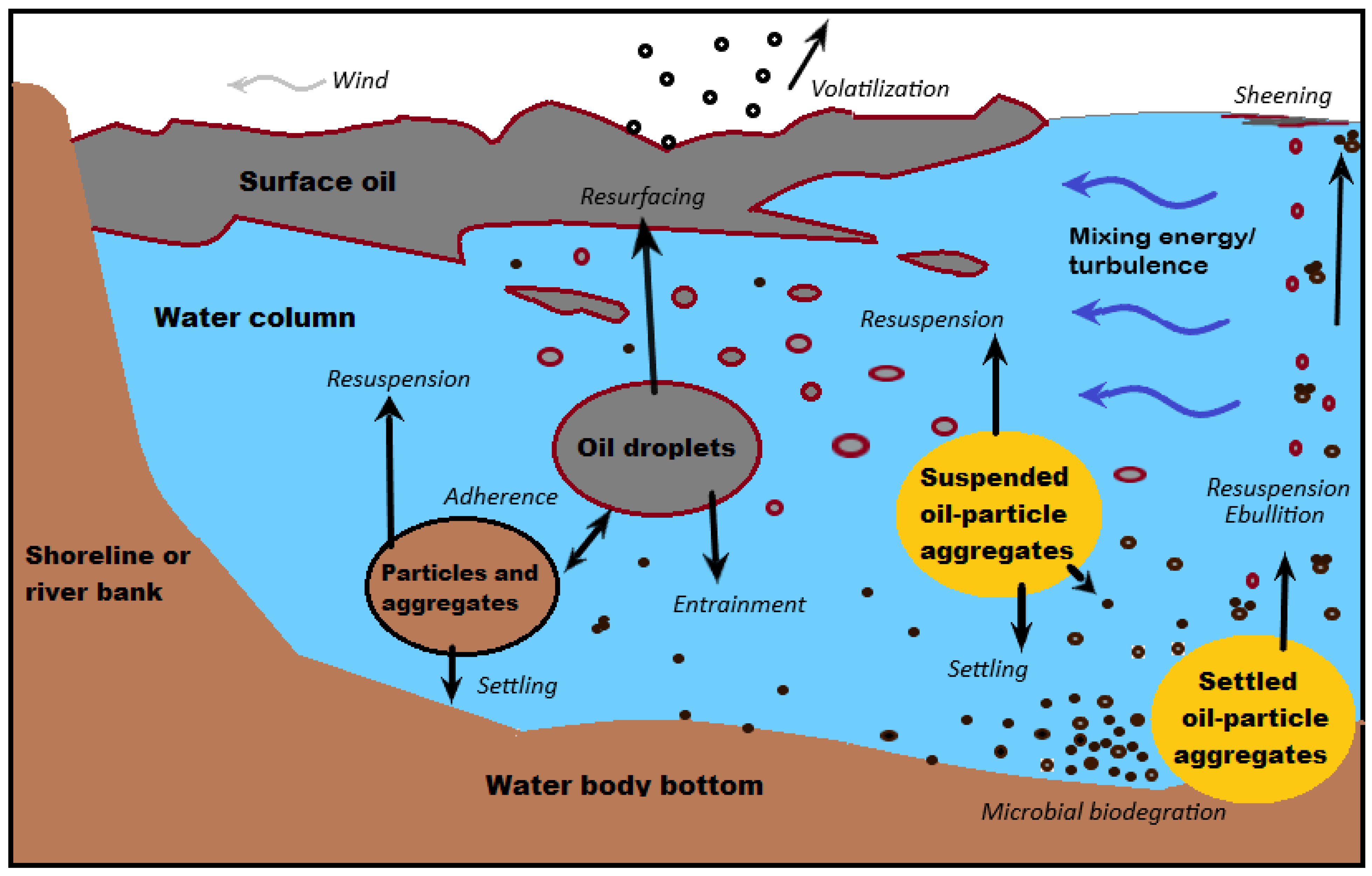 Figure 14. Simplified diagram of the processes and mechanisms leading to oil-particle aggregate formation and breakup in marine and freshwater environments. Modified from Fitzpatrick et al. [167].
Figure 14. Simplified diagram of the processes and mechanisms leading to oil-particle aggregate formation and breakup in marine and freshwater environments. Modified from Fitzpatrick et al. [167].The oil and mineral particle interactions also cause natural dissipation of spilled oil residues from shorelines over time. The interaction can occur in areas where there is no wave action or where mechanical wave energy is not present, such as a sheltered coastal environment. This cleanup procedure moves sediments around to make oil droplets more accessible, allowing for a faster physical engagement [173]. The more this contact occurs, the more nutrients, oxygen, and bacterial communities become accessible to the oil. This contact activates microbial activity by improving the rate and magnitude of oil degradation by increasing the surface area of the oil [172].
In addition, the mineral particles in marine and coastal environments act as supporting materials for the growth of bacteria. This can be seen in the results of the bacterial growth experiment by Chaerun et al. [160]. The formation of oil-particle aggregates extends and enhances the weathering processes of dissolution and evaporation by increasing the surface to volume ratio of oil. Water turbulence or breaking waves increase the transport and the formation of oil-particle aggregates. The flocculation rates of the oil and mineral are, in most cases, highest in low to intermediate salinity waters [174]. Chaerun et al. [160] show that minerals montmorillonite and kaolinite were able to promote the stimulation of microbial growth and the formation of biofilms, thereby resulting in the acceleration of bioremediation rates significantly. The authors also suggest that the clays used may be able to protect the microbes against the toxicity of high hydrocarbons concentrations. In the laboratory experiments done by Warr et al. [175] bioremediation of marine oil spills using clay mineral flakes was observed, showing the highest degree of alkane reduction by fertilizer clay treatment, with a reduction of 98%.
Interaction between the oil and the mineral particles will always result in the formation of oil-particle aggregates. Distinct mineral particles, on the other hand, might result in varying forms of oil-particle aggregates, as well as different hydrophobicity and hydrophilicity, stability, and buoyancy [176]. South American crude was found to be more resistive to hydrophilic mineral particles than Alaska North Slope crude, indicating that hydrophobic minerals interact more and have stronger oil and mineral particle attraction than hydrophilic minerals [177]. This, along with other variables in the interaction, will influence the fate of the oil-particle aggregates, whether they will float and be washed ashore by the waves, or whether they will stay and settle to the ocean’s bottom [176].
There are also organic materials that are the biotic counterpart of mineral particles such as fulvic acid, fecal pellets, phytoplankton, zooplankton, and bacteria that can all combine with oil to produce OPA-like aggregates. The production of oil-organic particle aggregates results from interactions between oil and organic materials, often known as marine snow [176]. Marine snow is small, stringy, veil-like mucous threads that form a vast web-like formation. Individual particles could be buoyant in one of three ways: positively, neutrally, or negatively [90]. They are formed by the aggregation of organic materials with sticky exopolymers, such as TEPs (transparent exopolymeric particles) and exopolymeric compounds (EPS) produced by microbial activity [178]. Oil droplets in the water column can become integrated during their aggregation. These web-like TEPs combine with oil and generate a web of strings that dangle in the water column, coalescing into huge marine snow particles. They eventually collide, generating a highly sticky mucus-rich marine snow that scavenges all particle kinds, including oil droplets, resulting in the formation of an even larger particle that can be over 500 µm in size and are responsible for the majority of organic material vertical export [176][179]. Following the Deepwater Horizon (DWH) and Ixtoc 1 oil spills, marine snow formation became the primary route for oil deposition [180][181].
Overall, it can be seen that the interaction of oil and mineral particles can work as a natural physical dispersant, and that, like man-made dispersants, it has the potential to harm marine and coastal environments. As previously stated, some oil-particle aggregates, as well as oil-organic particle aggregates, will settle to the ocean’s bottom, burying and suffocating smaller benthic species and causing habitat loss. Depending on the location of the affected areas, which may be sensitive and/or threatened, the cleanup of oil-particle aggregates can take a long time and cost a lot of money. Furthermore, in comparison to oil droplets in the water column, the toxicity of these oil-particle aggregates varies and is unclear [167].
2.11. Salinity
The solubility of toxic hydrocarbons from crude oil, the efficacy of chemical dispersants, and the binding properties of residual oil fractions into suspended particles can all be affected by aqueous salinity [182]. Moreover, microbial behavior is associated with salinity [183]. Mass transfer across cell membranes is the only way for biodegradation to start [184]. In the saline environment, microbial cells regulate cell surface hydrophobicity for water transportation to maintain a balance between extracellular and intracellular osmotic levels (Figure 15). At low salinities, the high cell surface hydrophobicity will stimulate cells to bind to the hydrophobic oil phase, accelerating the development of micelles and increasing hydrocarbon bioavailability. High salinities, on the other hand, causes water to permeate intracellularly to balance the osmotic level of bacterial cells, resulting in increased aqueous phase growth with decreased bio-emulsifier productivity [185].
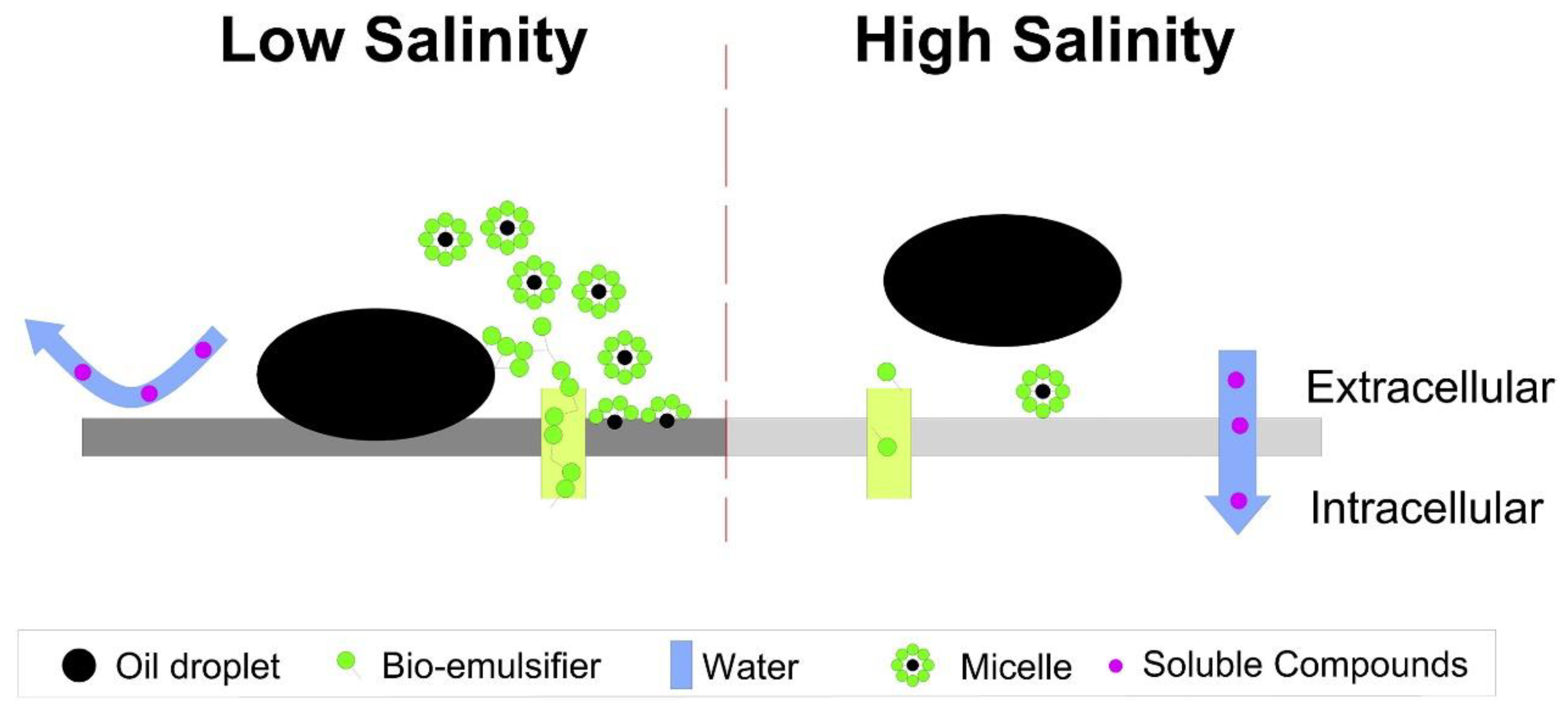 Figure 15. A schematic diagram describing the microbial eco-physiological mechanism when facing crude oil under low and high salinities. Adapted from Cao, et al. [185].
Figure 15. A schematic diagram describing the microbial eco-physiological mechanism when facing crude oil under low and high salinities. Adapted from Cao, et al. [185].Jamal and Pugazhendi [186], reported that the halophilic bacterial consortium could potentially degrade different PAHs (fluorene) up to 120 g/L. A phylogenetic analysis was also used in the study, revealing that the four degrading bacterial species belonged to the genera Dietzia, Arthrobacter, and Halomonas [186][187]. The bacteria Halomonas and Dietzia are important in the biodegradation of PAHs [187][188]. Arthrobacter, a member of the consortium, can degrade both aliphatic and aromatic hydrocarbons [189][190]. Mnif et al. [191] isolated halophilic bacterial strains of Halomonas sp. C2SS100 from a petroleum reservoir located near Kerkannah Island, Tunisia after enrichment of crude oil in the presence of 100 g/L NaCl at 37 °C. When refined petroleum products including diesel fuel and lubricating oil were used as the sole carbon source, biodegradation of heterogenic compounds increased significantly [191]. Moreover, Zhao et al. [192] reported that a halophilic bacterial consortium from the soil sample in Shengli Oilfield in China could completely degrade phenanthrene with 5–15% NaCl presence in eight days of enrichment. In the presence of 1–15% NaCl, Halomonas sp. and Marinobacter sp. degraded several PAHs, including naphthalene, phenanthrene, anthracene, fluoranthene, fluorine, benz(a)anthracene, and benzo(a)pyrene as carbon sources [193]. Several studies have also reported cases of halophilic archaea on their potential for degrading oil in saline environments including two extreme halophilic Haloferax strains and one strain each of Halobacterium and Halococcus [194] and a strain from the Natrialba genus [195]. Additionally, salinity influences the formation and dispersion of oil droplets [196]. Salinity is necessary for the flocculation of oil and suspended particulate matter (SPM). It influences the flocculation of fine materials and changes the surface morphology of oil droplets, thereby influencing the aggregation rate of suspended particulate matter and oil droplets [197]. Early research discovered that when oil and suspended particulate matter were mixed in seawater, they flocculated spontaneously [198], this is due to the chemistry of high-ionic-strength seawater that dominates the surface properties of the minerals [197].
It has also been reported that several species have been discovered to be capable of degrading a mixture of hydrocarbons in a wide range of salinities [199]. The effects of salinity on the bioremediation process include a direct inhibitory effect of metabolism due to the microbe’s environment’s unfavorable high osmotic potential, as well as altered solubility or sorption of toxic or crucial ions. It is important to note that the interactions between hydrocarbon-degrading bacteria and archaea can lead to more effective hydrocarbon biodegradation in hypersaline environments. Thus, more research into diversity would provide useful information for the development of reliable bioremediation processes for hypersaline environments.
2.12. pH
pH has an important impact on biodegradation processes in marine waters [200]. It affects biochemical processes, such as cell membrane transport, catalytic reaction balance and even poses alterations in enzyme activities. Moreover, the microbial transformation of contaminants (bioremediation) is directly correlated to the favorable environment [201], which includes the status of pH levels that is within the range of tolerance for the oil-degrading bacteria. Biodegradation creates interrelated reactions to the environment. As secondary metabolites are being produced during the biodegradation process, it could block the O2/CO2 exchange directly and lead to oxygen depletion and pH change in seawater [201]. However, while Kuppusamy et al. [202] claimed that direct oil-degradation remains independent of pH level, pH has an enormous influence on the oil-degrading bacteria. Pointek et al. [203] stated that pH perturbation to measure pH tolerance of oil-degrading bacteria is easily done and monitored in the laboratory but does not apply in the marine environment. Moreover, pH does not fluctuate much in the ocean (usually remains between 7.6–8.1) and does not appear to have an important effect on the biodegradation rate in most marine environments [204][205]. Only Okoh [206] reported that a slightly higher pH of seawater is favorable for petroleum hydrocarbon degradation. However, this entry also considered oil-degrading bacteria present in sediments and agreed that the environmental response of the bacteria is still the main driver of oil degradation.
To assess the pH tolerance of valuable oil-degrading bacteria, several studies reported the ability of bacteria to grow and degrade crude oil at various salinities ranging from moderate to high salinity environments [207]. Thavasi et al. [183] found that the maximum biodegradation of crude oil by Pseudomonas aeruginosa in water was at pH 8.0. The degradation of phenanthrene in liquid media was favorable within the range of pH values 6.5–7.0 by Burkholderia cocovenenans, isolated from petroleum-polluted soil [208]. Similarly, it was reported that oil-degrading bacteria has the highest percentage in the total microbial count (TMC) at pH 6 [209]. Das and Chadran [15] supported this claim stating that if microorganisms are present, then hydrocarbon biodegradation should be sustained between pH 6–9. Moreover, Pawar [208] and Rahman et al. [124] observed that pH 7.5 was favorable for most of the bacterial isolates and mixed bacterial consortium associated with the study but the strain of Flavobacterium sp. DS5-73 showed maximum percentage degradation (43%) when the pH is at 8.5. Moreover, Sathishkumar et al. [126], after studying a pH range of 4.0–7.0 found out that 7.0 pH is the optimum for maximum degradation. Given the above-mentioned literature, it could be interpreted that oil-degrading bacteria thrive in pH 6–9 and could not withstand the extreme pH environment. It is also taken into consideration how these bacteria respond in a pH perturbation set-up compared to a natural environment.
References
- Saadoun, I.M. Impact of oil spills on marine life. In Emerging Pollutants in the Environment-Current and Further Implications; BoD–Books on Demand: Norderstedt, Germany, 2015; pp. 75–104.
- Murphy, D.; Gemmell, B.; Vaccari, L.; Li, C.; Bacosa, H.; Evans, M.; Gemmell, C.; Harvey, T.; Jalali, M.; Niepa, T.H. An in-depth survey of the oil spill literature since 1968: Long term trends and changes since Deepwater Horizon. Mar. Pollut. Bull. 2016, 113, 371–379.
- Reddy, C.M.; Arey, J.S.; Seewald, J.S.; Sylva, S.P.; Lemkau, K.L.; Nelson, R.K.; Carmichael, C.A.; McIntyre, C.P.; Fenwick, J.; Ventura, G.T. Composition and fate of gas and oil released to the water column during the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20229–20234.
- Crone, T.J.; Tolstoy, M. Magnitude of the 2010 Gulf of Mexico oil leak. Science 2010, 330, 634.
- Bacosa, H.P.; Evans, M.M.; Wang, Q.; Liu, Z. Assessing the role of environmental conditions on the degradation of oil following the deepwater horizon oil spill. In Oil Spill Environmental Forensics Case Studies; Elsevier: Amsterdam, The Netherlands, 2018; pp. 617–637.
- Goldstein, B.D.; Osofsky, H.J.; Lichtveld, M.Y. The Gulf oil spill. N. Engl. J. Med. 2011, 364, 1334–1348.
- Chen, Q.; Bao, B.; Li, Y.; Liu, M.; Zhu, B.; Mu, J.; Chen, Z. Effects of marine oil pollution on microbial diversity in coastal waters and stimulating indigenous microorganism bioremediation with nutrients. Reg. Stud. Mar. Sci. 2020, 39, 101395.
- Hassanshahian, M.; Cappello, S. Crude oil biodegradation in the marine environments. Biodegrad.-Eng. Technol. 2013, 5, 102–135.
- Park, B.S.; Erdner, D.L.; Bacosa, H.P.; Liu, Z.; Buskey, E.J. Potential effects of bacterial communities on the formation of blooms of the harmful dinoflagellate Prorocentrum after the 2014 Texas City “Y” oil spill (USA). Harmful Algae 2020, 95, 101802.
- Kingston, P.F. Long-term environmental impact of oil spills. Spill Sci. Technol. Bull. 2002, 7, 53–61.
- Fingas, M. Oil Spill Science and Technology; Gulf Professional Publishing: Oxford, UK, 2016.
- Kujawinski, E.B.; Kido Soule, M.C.; Valentine, D.L.; Boysen, A.K.; Longnecker, K.; Redmond, M.C. Fate of dispersants associated with the Deepwater Horizon oil spill. Environ. Sci. Technol. 2011, 45, 1298–1306.
- Abbasian, F.; Lockington, R.; Mallavarapu, M.; Naidu, R. A comprehensive review of aliphatic hydrocarbon biodegradation by bacteria. Appl. Biochem. Biotechnol. 2015, 176, 670–699.
- Overton, E.B.; Wetzel, D.L.; Wickliffe, J.K.; Adhikari, P.L. Spilled Oil Composition and the Natural Carbon Cycle: The True Drivers of Environmental Fate and Effects of Oil Spills. In Scenarios and Responses to Future Deep Oil Spills; Springer: Berlin/Heidelberg, Germany, 2020; pp. 33–56.
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810.
- Bacosa, H.; Suto, K.; Inoue, C. Preferential degradation of aromatic hydrocarbons in kerosene by a microbial consortium. Int. Biodeterior. Biodegrad. 2010, 64, 702–710.
- Bacosa, H.P.; Suto, K.; Inoue, C. Degradation potential and microbial community structure of heavy oil-enriched microbial consortia from mangrove sediments in Okinawa, Japan. J. Environ. Sci. Health Part A 2013, 48, 835–846.
- Christova, N.; Kabaivanova, L.; Nacheva, L.; Petrov, P.; Stoineva, I. Biodegradation of crude oil hydrocarbons by a newly isolated biosurfactant producing strain. Biotechnol. Biotechnol. Equip. 2019, 33, 863–872.
- Bacosa, H.P.; Inoue, C. Polycyclic aromatic hydrocarbons (PAHs) biodegradation potential and diversity of microbial consortia enriched from tsunami sediments in Miyagi, Japan. J. Hazard. Mater. 2015, 283, 689–697.
- Dominguez, J.J.A.; Bacosa, H.; Chien, M.-F.; Inoue, C. Enhanced degradation of polycyclic aromatic hydrocarbons (PAHs) in the rhizosphere of sudangrass (Sorghum × drummondii). Chemosphere 2019, 234, 789–795.
- Keramea, P.; Spanoudaki, K.; Zodiatis, G.; Gikas, G.; Sylaios, G. Oil spill modeling: A critical review on current trends, perspectives, and challenges. J. Mar. Sci. Eng. 2021, 9, 181.
- Bretherton, L.; Hillhouse, J.; Bacosa, H.; Setta, S.; Genzer, J.; Kamalanathan, M.; Finkel, Z.V.; Quigg, A. Growth dynamics and domoic acid production of Pseudo-nitzschia sp. in response to oil and dispersant exposure. Harmful Algae 2019, 86, 55–63.
- Kamalanathan, M.; Chiu, M.-H.; Bacosa, H.; Schwehr, K.; Tsai, S.-M.; Doyle, S.; Yard, A.; Mapes, S.; Vasequez, C.; Bretherton, L.; et al. Role of polysaccharides in diatom Thalassiosira pseudonana and its associated bacteria in hydrocarbon presence. Plant Physiol. 2019, 180, 1898–1911.
- Severin, T.; Bacosa, H.; Sato, A.; Erdner, D. Dynamics of Heterocapsa sp. and the associated attached and free-living bacteria under the influence of dispersed and undispersed crude oil. Lett. Appl. Microbiol. 2016, 63, 419–425.
- McGenity, T.J.; Folwell, B.D.; McKew, B.A.; Sanni, G.O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Biosyst. 2012, 8, 10.
- Bacosa, H.P.; Thyng, K.M.; Plunkett, S.; Erdner, D.L.; Liu, Z. The tarballs on Texas beaches following the 2014 Texas City “Y” Spill: Modeling, chemical, and microbiological studies. Mar. Pollut. Bull. 2016, 109, 236–244.
- Hazen, T.C.; Prince, R.C.; Mahmoudi, N. Marine Oil Biodegradation; American Chemical Society (ACS) Publications: Washington, DC, USA, 2016.
- Steichen, J.L.; Labonté, J.M.; Windham, R.; Hala, D.; Kaiser, K.; Setta, S.; Faulkner, P.C.; Bacosa, H.; Yan, G.; Kamalanathan, M. Microbial, physical, and chemical changes in Galveston Bay following an extreme flooding event, Hurricane Harvey. Front. Mar. Sci. 2020, 7, 186.
- Bacosa, H.P.; Steichen, J.; Kamalanathan, M.; Windham, R.; Lubguban, A.; Labonté, J.M.; Kaiser, K.; Hala, D.; Santschi, P.H.; Quigg, A. Polycyclic aromatic hydrocarbons (PAHs) and putative PAH-degrading bacteria in Galveston Bay, TX (USA), following Hurricane Harvey (2017). Environ. Sci. Pollut. Res. 2020, 27, 34987–34999.
- Abed, R.M.; Köster, J. The direct role of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegrad. 2005, 55, 29–37.
- Williams, A.K.; Bacosa, H.P.; Quigg, A. The impact of dissolved inorganic nitrogen and phosphorous on responses of microbial plankton to the Texas City “Y” oil spill in Galveston Bay, Texas (USA). Mar. Pollut. Bull. 2017, 121, 32–44.
- Gemmell, B.J.; Bacosa, H.P.; Dickey, B.O.; Gemmell, C.G.; Alqasemi, L.R.; Buskey, E.J. Rapid alterations to marine microbiota communities following an oil spill. Ecotoxicology 2018, 27, 505–516.
- Bacosa, H.P.; Erdner, D.L.; Liu, Z. Differentiating the roles of photooxidation and biodegradation in the weathering of Light Louisiana Sweet crude oil in surface water from the Deepwater Horizon site. Mar. Pollut. Bull. 2015, 95, 265–272.
- Bacosa, H.P.; Erdner, D.L.; Rosenheim, B.E.; Shetty, P.; Seitz, K.W.; Baker, B.J.; Liu, Z. Hydrocarbon degradation and response of seafloor sediment bacterial community in the northern Gulf of Mexico to light Louisiana sweet crude oil. ISME J. 2018, 12, 2532–2543.
- Chikere, C.B.; Okpokwasili, G.C.; Chikere, B.O. Monitoring of microbial hydrocarbon remediation in the soil. 3 Biotech 2011, 1, 117–138.
- Transportation Research Board and National Research Council. Oil in the Sea III: Inputs, Fates, and Effects; The National Academies Press: Washington, DC, USA, 2003.
- Campo, P.; Venosa, A.D.; Suidan, M.T. Biodegradability of Corexit 9500 and dispersed South Louisiana crude oil at 5 and 25 °C. Environ. Sci. Technol. 2013, 47, 1960–1967.
- Meng, L.; Liu, H.; Bao, M.; Sun, P. Microbial community structure shifts are associated with temperature, dispersants and nutrients in crude oil-contaminated seawaters. Mar. Pollut. Bull. 2016, 111, 203–212.
- Liu, J.; Bacosa, H.P.; Liu, Z. Potential environmental factors affecting oil-degrading bacterial populations in deep and surface waters of the northern Gulf of Mexico. Front. Microbiol. 2017, 7, 2131.
- Bacosa, H.P.; Kang, A.; Lu, K.; Liu, Z. Initial oil concentration affects hydrocarbon biodegradation rates and bacterial community composition in seawater. Mar. Pollut. Bull. 2021, 162, 111867.
- Ostroumov, V.; Siegert, C. Exobiological aspects of mass transfer in microzones of permafrost deposits. Adv. Space Res. 1996, 18, 79–86.
- Aislabie, J.; Saul, D.J.; Foght, J.M. Bioremediation of hydrocarbon-contaminated polar soils. Extremophiles 2006, 10, 171–179.
- AUNG, M.T.M.; LI, Q.; TAKAHASHI, S.; UTSUMI, M. Effect of Temperature on Hydrocarbon Bioremediation in Simulated Petroleum-Polluted Seawater Collected from Tokyo Bay. Jpn. J. Water Treat. Biol. 2018, 54, 95–104.
- Siron, R.; Pelletier, E.; Brochu, C. Environmental factors influencing the biodegradation of petroleum hydrocarbons in cold seawater. Arch. Environ. Contam. Toxicol. 1995, 28, 406–416.
- Redmond, M.C.; Valentine, D.L. Natural gas and temperature structured a microbial community response to the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20292–20297.
- Liu, Z.; Liu, J. Evaluating bacterial community structures in oil collected from the sea surface and sediment in the northern Gulf of Mexico after the Deepwater Horizon oil spill. Microbiol. Open 2013, 2, 492–504.
- Coulon, F.; Pelletier, E.; Gourhant, L.; Delille, D. Effects of nutrient and temperature on degradation of petroleum hydrocarbons in contaminated sub-Antarctic soil. Chemosphere 2005, 58, 1439–1448.
- Kirchman, D.L. Processes in Microbial Ecology; Oxford University Press: Oxford, UK, 2018.
- Vázquez-Domínguez, E.; Vaqué, D.; Gasol, J.M. Temperature effects on the heterotrophic bacteria, heterotrophic nanoflagellates, and microbial top predators of the NW Mediterranean. Aquat. Microb. Ecol. 2012, 67, 107–121.
- Morán, X.A.G.; Alonso-Sáez, L.; Nogueira, E.; Ducklow, H.W.; González, N.; López-Urrutia, Á.; Díaz-Pérez, L.; Calvo-Díaz, A.; Arandia-Gorostidi, N.; Huete-Stauffer, T.M. More, smaller bacteria in response to ocean’s warming? Proc. R. Soc. B 2015, 282, 20150371.
- Semboung Lang, F.; Tarayre, C.; Destain, J.; Delvigne, F.; Druart, P.; Ongena, M.; Thonart, P. The effect of nutrients on the degradation of hydrocarbons in mangrove ecosystems by microorganisms. Int. J. Environ. Res. 2016, 10, 583–592.
- Vyas, T.K.; Dave, B. Effect of addition of nitrogen, phosphorus and potassium fertilizers on biodegradation of crude oil by marine bacteria. IJMS 2010, 39, 143–150.
- Harrison, S.J. Lessons from the Taylor Energy Oil Spill: History, Seasonality, and Nutrient Limitation; University of Georgia: Athens, GA, USA, 2017.
- Edwards, B.R.; Reddy, C.M.; Camilli, R.; Carmichael, C.A.; Longnecker, K.; Van Mooy, B.A. Rapid microbial respiration of oil from the Deepwater Horizon spill in offshore surface waters of the Gulf of Mexico. Environ. Res. Lett. 2011, 6, 035301.
- Dubinsky, E.A.; Conrad, M.E.; Chakraborty, R.; Bill, M.; Borglin, S.E.; Hollibaugh, J.T.; Mason, O.U.; Piceno, M.Y.; Reid, F.C.; Stringfellow, W.T. Succession of hydrocarbon-degrading bacteria in the aftermath of the Deepwater Horizon oil spill in the Gulf of Mexico. Environ. Sci. Technol. 2013, 47, 10860–10867.
- Elango, V.; Urbano, M.; Lemelle, K.R.; Pardue, J.H. Biodegradation of MC252 oil in oil: Sand aggregates in a coastal headland beach environment. Front. Microbiol. 2014, 5, 161.
- Alabresm, A.; Chen, Y.P.; Decho, A.W.; Lead, J. A novel method for the synergistic remediation of oil-water mixtures using nanoparticles and oil-degrading bacteria. Sci. Total Environ. 2018, 630, 1292–1297.
- Xu, J.; Du, J.; Li, L.; Zhang, Q.; Chen, Z. Fast-stimulating bioremediation of macro crude oil in soils using matching Fenton pre-oxidation. Chemosphere 2020, 252, 126622.
- Zahid, M.R.; Khamaruddin, P.F.M.; Him, N.R.N.; Othman, N.H.; Yahya, E. Effect of Organic Nutrient Addition to the Biodegradation of Hydrocarbon Contaminated Marine Sediment in Malaysia. In Key Engineering Materials; Trans Tech Publications Ltd.: Kepellwag, Switzerland, 2019; pp. 74–83.
- Chaillan, F.; Le Flèche, A.; Bury, E.; Phantavong, Y.-h.; Grimont, P.; Saliot, A.; Oudot, J. Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res. Microbiol. 2004, 155, 587–595.
- Oudot, J.; Merlin, F.; Pinvidic, P. Weathering rates of oil components in a bioremediation experiment in estuarine sediments. Mar. Environ. Res. 1998, 45, 113–125.
- Chaineau, C.; Rougeux, G.; Yepremian, C.; Oudot, J. Effects of nutrient concentration on the biodegradation of crude oil and associated microbial populations in the soil. Soil Biol. Biochem. 2005, 37, 1490–1497.
- Thavasi, R.; Jayalakshmi, S.; Banat, I.M. Application of biosurfactant produced from peanut oil cake by Lactobacillus delbrueckii in biodegradation of crude oil. Bioresour. Technol. 2011, 102, 3366–3372.
- Farag, S.; Soliman, N.A.; Abdel-Fattah, Y.R. Statistical optimization of crude oil bio-degradation by a local marine bacterium isolate Pseudomonas sp. sp48. J. Genet. Eng. Biotechnol. 2018, 16, 409–420.
- Souza, E.C.; Vessoni-Penna, T.C.; de Souza Oliveira, R.P. Biosurfactant-enhanced hydrocarbon bioremediation: An overview. Int. Biodeterior. Biodegrad. 2014, 89, 88–94.
- Waikhom, D.; Ngasotter, S.; Soniya Devi, L.; Devi, S.; Singh, A.S. Role of Microbes in Petroleum Hydrocarbon Degradation in the Aquatic Environment: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 2890–2903.
- Brakstad, O.G.; Lofthus, S.; Ribicic, D.; Netzer, R. Biodegradation of petroleum oil in cold marine environments. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 613–644.
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. Wiley Interdiscip. Rev. 2019, 6, e1373.
- Huettel, M.; Berg, P.; Kostka, J.E. Benthic exchange and biogeochemical cycling in permeable sediments. Annu. Rev. Mar. Sci. 2014, 6, 23–51.
- Joye, S.B.; Teske, A.P.; Kostka, J.E. Microbial dynamics following the Macondo oil well blowout across Gulf of Mexico environments. Bioscience 2014, 64, 766–777.
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L. Deep-sea oil plume enriches indigenous oil-degrading bacteria. Science 2010, 330, 204–208.
- Walworth, J.; Pond, A.; Snape, I.; Rayner, J.; Ferguson, S.; Harvey, P. Nitrogen requirements for maximizing petroleum bioremediation in a sub-Antarctic soil. Cold Reg. Sci. Technol. 2007, 48, 84–91.
- Aeppli, C.; Carmichael, C.A.; Nelson, R.K.; Lemkau, K.L.; Graham, W.M.; Redmond, M.C.; Valentine, D.L.; Reddy, C.M. Oil weathering after the Deepwater Horizon disaster led to the formation of oxygenated residues. Environ. Sci. Technol. 2012, 46, 8799–8807.
- Tarr, M.A.; Zito, P.; Overton, E.B.; Olson, G.M.; Adhikari, P.L.; Reddy, C.M. Weathering of oil spilled in the marine environment. Oceanography 2016, 29, 126–135.
- King, S.M.; Leaf, P.A.; Olson, A.C.; Ray, P.Z.; Tarr, M.A. Photolytic and photocatalytic degradation of surface oil from the Deepwater Horizon spill. Chemosphere 2014, 95, 415–422.
- D’Auria, M.; Emanuele, L.; Racioppi, R.; Velluzzi, V. Photochemical degradation of crude oil: Comparison between direct irradiation, photocatalysis, and photocatalysis on zeolite. J. Hazard. Mater. 2009, 164, 32–38.
- Ward, C.P.; Overton, E.B. How the 2010 Deepwater Horizon spill reshaped our understanding of crude oil photochemical weathering at sea: A past, present, and future perspective. Environ. Sci. 2020, 22, 1125–1138.
- Ziolli, R.L.; Jardim, W.F. Photochemical transformations of water-soluble fraction (WSF) of crude oil in marine waters: A comparison between photolysis and accelerated degradation with TiO2 using GC–MS and UVF. J. Photochem. Photobiol. A 2003, 155, 243–252.
- Griffiths, M.T.; Da Campo, R.; O’Connor, P.B.; Barrow, M.P. Throwing light on petroleum: Simulated exposure of crude oil to sunlight and characterization using atmospheric pressure photoionization Fourier transform ion cyclotron resonance mass spectrometry. Anal. Chem. 2014, 86, 527–534.
- Shankar, R.; Shim, W.J.; An, J.G.; Yim, U.H. A practical review on photooxidation of crude oil: Laboratory lamp setup and factors affecting it. Water Res. 2015, 68, 304–315.
- Hall, G.J.; Frysinger, G.S.; Aeppli, C.; Carmichael, C.A.; Gros, J.; Lemkau, K.L.; Nelson, R.K.; Reddy, C.M. Oxygenated weathering products of Deepwater Horizon oil come from surprising precursors. Mar. Pollut. Bull. 2013, 75, 140–149.
- Radović, J.R.; Aeppli, C.; Nelson, R.K.; Jimenez, N.; Reddy, C.M.; Bayona, J.M.; Albaiges, J. Assessment of photochemical processes in marine oil spill fingerprinting. Mar. Pollut. Bull. 2014, 79, 268–277.
- Dutta, T.K.; Harayama, S. Fate of crude oil by the combination of photooxidation and biodegradation. Environ. Sci. Technol. 2000, 34, 1500–1505.
- Bacosa, H.P.; Liu, Z.; Erdner, D.L. Natural sunlight shapes crude oil-degrading bacterial communities in Northern Gulf of Mexico surface waters. Front. Microbiol. 2015, 6, 1325.
- Ray, P.Z.; Tarr, M.A. Solar production of singlet oxygen from crude oil films on water. J. Photochem. Photobiol. A 2014, 286, 22–28.
- Ray, P.Z.; Tarr, M.A. Petroleum films exposed to sunlight produce hydroxyl radical. Chemosphere 2014, 103, 220–227.
- Plata, D.L.; Sharpless, C.M.; Reddy, C.M. Photochemical degradation of polycyclic aromatic hydrocarbons in oil films. Environ. Sci. Technol. 2008, 42, 2432–2438.
- Bacosa, H.P.; Kamalanathan, M.; Chiu, M.-H.; Tsai, S.-M.; Sun, L.; Labonté, J.M.; Schwehr, K.A.; Hala, D.; Santschi, P.H.; Chin, W.-C. Extracellular polymeric substances (EPS) producing and oil degrading bacteria isolated from the northern Gulf of Mexico. PLoS ONE 2018, 13, e0208406.
- Sun, L.; Chiu, M.-H.; Xu, C.; Lin, P.; Schwehr, K.A.; Bacosa, H.; Kamalanathan, M.; Quigg, A.; Chin, W.-C.; Santschi, P.H. The effects of sunlight on the composition of exopolymeric substances and subsequent aggregate formation during oil spills. Mar. Chem. 2018, 203, 49–54.
- Passow, U.; Ziervogel, K.; Asper, V.; Diercks, A. Marine snow formation in the aftermath of the Deepwater Horizon oil spill in the Gulf of Mexico. Environ. Res. Lett. 2012, 7, 035301.
- Bacosa, H.P.; Kamalanathan, M.; Cullen, J.; Shi, D.; Xu, C.; Schwehr, K.A.; Hala, D.; Wade, T.L.; Knap, A.H.; Santschi, P.H. Marine snow aggregates are enriched in polycyclic aromatic hydrocarbons (PAHs) in oil contaminated waters: Insights from a mesocosm study. J. Mar. Sci. Eng. 2020, 8, 781.
- Shiu, R.-F.; Chiu, M.-H.; Vazquez, C.I.; Tsai, Y.-Y.; Le, A.; Kagiri, A.; Xu, C.; Kamalanathan, M.; Bacosa, H.P.; Doyle, S.M. Protein to carbohydrate (P/C) ratio changes in microbial extracellular polymeric substances induced by oil and Corexit. Mar. Chem. 2020, 223, 103789.
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734.
- Sun, L.; Xu, C.; Zhang, S.; Lin, P.; Schwehr, K.A.; Quigg, A.; Chiu, M.-H.; Chin, W.-C.; Santschi, P.H. Light-induced aggregation of microbial exopolymeric substances. Chemosphere 2017, 181, 675–681.
- Venosa, A.D.; Zhu, X. Biodegradation of crude oil contaminating marine shorelines and freshwater wetlands. Spill Sci. Technol. Bull. 2003, 8, 163–178.
- Meng, Q.; Wang, X.; Wang, X.; Shi, B.; Luo, X.; Zhang, L.; Lei, Y.; Jiang, C.; Liu, P. Gas geochemical evidences for biodegradation of shale gases in the Upper Triassic Yanchang Formation, Ordos Basin, China. Int. J. Coal Geol. 2017, 179, 139–152.
- Sierra-Garcia, I.N.; de Oliveira, V.M. Microbial hydrocarbon degradation: Efforts to understand biodegradation in petroleum reservoirs. Biodegrad.-Eng. Technol. 2013, 10, 55920.
- McNally, D.L.; Mihelcic, J.R.; Lueking, D.R. Biodegradation of three-and four-ring polycyclic aromatic hydrocarbons under aerobic and denitrifying conditions. Environ. Sci. Technol. 1998, 32, 2633–2639.
- Grishchenkov, V.; Townsend, R.; McDonald, T.; Autenrieth, R.; Bonner, J.; Boronin, A. Degradation of petroleum hydrocarbons by facultative anaerobic bacteria under aerobic and anaerobic conditions. Process Biochem. 2000, 35, 889–896.
- Olajire, A.; Essien, J. Aerobic degradation of petroleum components by microbial consortia. J. Pet. Environ. Biotechnol. 2014, 5, 1.
- Mille, G.; Mulyono, M.; El Jammal, T.; Bertrand, J.-C. Effects of oxygen on hydrocarbon degradation studies in vitro in surficial sediments. Estuar. Coast. Shelf Sci. 1988, 27, 283–295.
- Kostka, J.E.; Joye, S.B.; Overholt, W.; Bubenheim, P.; Hackbusch, S.; Larter, S.R.; Liese, A.; Lincoln, S.A.; Marietou, A.; Müller, R. Biodegradation of petroleum hydrocarbons in the deep sea. In Deep Oil Spills; Springer: Berlin/Heidelberg, Germany, 2020; pp. 107–124.
- Nicholson, C.A.; Fathepure, B.Z. Biodegradation of benzene by halophilic and halotolerant bacteria under aerobic conditions. Appl. Environ. Microbiol. 2004, 70, 1222–1225.
- Yakimov, M.M.; Golyshin, P.N.; Lang, S.; Moore, E.R.; Abraham, W.-R.; Lünsdorf, H.; Timmis, K.N. Alcanivorax borkumensis gen. nov., sp. nov., a new, hydrocarbon-degrading and surfactant-producing marine bacterium. Int. J. Syst. Evol. Microbiol. 1998, 48, 339–348.
- Dyksterhouse, S.E.; Gray, J.P.; Herwig, R.P.; Lara, J.C.; Staley, J.T. Cycloclasticus pugetii gen. nov., sp. nov., an aromatic hydrocarbon-degrading bacterium from marine sediments. Int. J. Syst. Evol. Microbiol. 1995, 45, 116–123.
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266.
- Atlas, R.M. Petroleum biodegradation and oil spill bioremediation. Mar. Pollut. Bull. 1995, 31, 178–182.
- Lee, W.; Morris, A.; Boatwright, D. Mexican oil spill: A toxicity study of oil accommodated in seawater on marine invertebrates. Mar. Pollut. Bull. 1980, 11, 231–234.
- Teramoto, M.; Suzuki, M.; Okazaki, F.; Hatmanti, A.; Harayama, S. Oceanobacter-related bacteria are important for the degradation of petroleum aliphatic hydrocarbons in the tropical marine environment. Microbiology 2009, 155, 3362–3370.
- Maruyama, A.; Ishiwata, H.; Kitamura, K.; Sunamura, M.; Fujita, T.; Matsuo, M.; Higashihara, T. Dynamics of microbial populations and strong selection for Cycloclasticus pugetii following the Nakhodka oil spill. Microb. Ecol. 2003, 46, 442–453.
- Jimeson, R.; Radosevich, M.; Stevens, R.; Giardino-Radosevich, G. Mixed Alcohol Fuels for Internal Combustion Engines, Furnaces, Boilers, Kilns and Gasifiers. U.S. Patent 8,277,522 B2, 2 October 2012.
- Chou, C.-C.; Riviere, J.E.; Monteiro-Riviere, N.A. Differential relationship between the carbon chain length of jet fuel aliphatic hydrocarbons and their ability to induce cytotoxicity vs. interleukin-8 release in human epidermal keratinocytes. Toxicol. Sci. 2002, 69, 226–233.
- Istoto, E.H.; Saptadi, S. Production of Fuels From HDPE and LDPE Plastic Waste via Pyrolysis Methods. In E3S Web of Conferences; EDP Sciences: Ulis, France, 2019; p. 14011.
- Bayat, H.; Cheng, F.; Dehghanizadeh, M.; Soliz, N.; Brewer, C.E.; Jena, U. Hydrothermal Liquefaction of Food Waste: Bio-crude oil Characterization, Mass and Energy Balance. In 2019 ASABE Annual International Meeting; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2019.
- Cerniglia, C. Microbial degradation of polycyclic aromatic hydrocarbons (PAH) in the aquatic environment. Metab. Polycycl. Aromat. Hydrocarb. Aquat. Environ. 1989, 38, 41–68.
- Solano-Serena, F.; Marchal, R.; Casarégola, S.; Vasnier, C.; Lebeault, J.-M.; Vandecasteele, J.-P. A Mycobacterium strain with extended capacities for degradation of gasoline hydrocarbons. Appl. Environ. Microbiol. 2000, 66, 2392–2399.
- Azevedo, L.B.; van Zelm, R.; Elshout, P.M.; Hendriks, A.J.; Leuven, R.S.; Struijs, J.; de Zwart, D.; Huijbregts, M.A. Species richness–phosphorus relationships for lakes and streams worldwide. Glob. Ecol. Biogeogr. 2013, 22, 1304–1314.
- Zhou, Y.; Gu, X.; Zhang, R.; Chen, L.; Lu, J. Microbial degradation of diesel oil and heavy oil in the presence of modified clay. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 326–331.
- Adebusoye, S.A.; Ilori, M.O.; Amund, O.O.; Teniola, O.D.; Olatope, S. Microbial degradation of petroleum hydrocarbons in a polluted tropical stream. World J. Microbiol. Biotechnol. 2007, 23, 1149–1159.
- Mandri, T.; Lin, J. Isolation and characterization of engine oil degrading indigenous microrganisms in Kwazulu-Natal, South Africa. Afr. J. Biotechnol. 2007, 6, 23–27.
- Darvishi, P.; Mowla, D.; Ayatollahi, S.; Niazi, A. Biodegradation of heavy crude oil in wastewater by an efficient strain, ERCPPI-1. Desalination Water Treat. 2011, 28, 46–54.
- Rajasekar, A.; Maruthamuthu, S.; Ting, Y.-P.; Balasubramanian, R.; Rahman, P.K. Bacterial degradation of petroleum hydrocarbons. In Microbial Degradation of Xenobiotics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 339–369.
- Bacosa, H.P.; Mabuhay-Omar, J.A.; Balisco, R.A.T.; Omar, D.M.; Inoue, C. Biodegradation of binary mixtures of octane with benzene, toluene, ethylbenzene or xylene (BTEX): Insights on the potential of Burkholderia, Pseudomonas and Cupriavidus isolates. World J. Microbiol. Biotechnol. 2021, 37, 122.
- Rahman, K.; Thahira-Rahman, J.; Lakshmanaperumalsamy, P.; Banat, I.M. Towards efficient crude oil degradation by a mixed bacterial consortium. Bioresour. Technol. 2002, 85, 257–261.
- Tzarkova, E.; Groudeva, V. Bioremediation of Petroleum—Polluted Soils by Adsorptive Immobilized Corynebacterium Sp. RB—96. Biotechnol. Biotechnol. Equip. 2000, 14, 72–75.
- Sathishkumar, M.; Binupriya, A.R.; Baik, S.H.; Yun, S.E. Biodegradation of crude oil by individual bacterial strains and a mixed bacterial consortium isolated from hydrocarbon contaminated areas. CLEAN—Soil Air Water 2008, 36, 92–96.
- Nguyen, U.T.; Lincoln, S.A.; Valladares Juárez, A.G.; Schedler, M.; Macalady, J.L.; Müller, R.; Freeman, K.H. The influence of pressure on crude oil biodegradation in shallow and deep Gulf of Mexico sediments. PLoS ONE 2018, 13, e0199784.
- Schedler, M.; Hiessl, R.; Juárez, A.G.V.; Gust, G.; Müller, R. Effect of high pressure on hydrocarbon-degrading bacteria. AMB Express 2014, 4, 77.
- Scoma, A.; Boon, N. Osmotic stress confers enhanced cell integrity to hydrostatic pressure but impairs growth in Alcanivorax borkumensis SK2. Front. Microbiol. 2016, 7, 729.
- Scoma, A.; Yakimov, M.M.; Boon, N. Challenging oil bioremediation at deep-sea hydrostatic pressure. Front. Microbiol. 2016, 7, 1203.
- Hackbusch, S.; Noirungsee, N.; Viamonte, J.; Sun, X.; Bubenheim, P.; Kostka, J.E.; Müller, R.; Liese, A. Influence of pressure and dispersant on oil biodegradation by a newly isolated Rhodococcus strain from deep-sea sediments of the Gulf of Mexico. Mar. Pollut. Bull. 2020, 150, 110683.
- Dutta, A.; Peoples, L.M.; Gupta, A.; Bartlett, D.H.; Sar, P. Exploring the piezotolerant/piezophilic microbial community and genomic basis of piezotolerance within the deep subsurface Deccan traps. Extremophiles 2019, 23, 421–433.
- Yayanos, A.A. Microbiology to 10,500 meters in the deep sea. Annu. Rev. Microbiol. 1995, 49, 777–805.
- Oger, P.M.; Jebbar, M. The many ways of coping with pressure. Res. Microbiol. 2010, 161, 799–809.
- Mason, O.U.; Han, J.; Woyke, T.; Jansson, J.K. Single-cell genomics reveals features of a Colwellia species that was dominant during the Deepwater Horizon oil spill. Front. Microbiol. 2014, 5, 332.
- Marietou, A.; Chastain, R.; Beulig, F.; Scoma, A.; Hazen, T.C.; Bartlett, D.H. The effect of hydrostatic pressure on enrichments of hydrocarbon degrading microbes from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 2018, 9, 808.
- Grossi, V.; Yakimov, M.M.; Al Ali, B.; Tapilatu, Y.; Cuny, P.; Goutx, M.; La Cono, V.; Giuliano, L.; Tamburini, C. Hydrostatic pressure affects membrane and storage lipid compositions of the piezotolerant hydrocarbon-degrading Marinobacter hydrocarbonoclasticus strain# 5. Environ. Microbiol. 2010, 12, 2020–2033.
- Perez Calderon, L.J.; Gontikaki, E.; Potts, L.D.; Shaw, S.; Gallego, A.; Anderson, J.A.; Witte, U. Pressure and temperature effects on deep-sea hydrocarbon-degrading microbial communities in subarctic sediments. Microbiologyopen 2019, 8, e00768.
- ZoBell, C.E.; Johnson, F.H. The influence of hydrostatic pressure on the growth and viability of terrestrial and marine bacteria. J. Bacteriol. 1949, 57, 179–189.
- Prince, R.C.; Lessard, R.R. Crude oil releases to the environment: Natural fate and remediation options. Encycl. Energy 2004, 1, 727–736.
- Scoma, A.; Barbato, M.; Hernandez-Sanabria, E.; Mapelli, F.; Daffonchio, D.; Borin, S.; Boon, N. Microbial oil-degradation under mild hydrostatic pressure (10 MPa): Which pathways are impacted in piezosensitive hydrocarbonoclastic bacteria? Sci. Rep. 2016, 6, 23526.
- Kimes, N.E.; Callaghan, A.V.; Aktas, D.F.; Smith, W.L.; Sunner, J.; Golding, B.T.; Drozdowska, M.; Hazen, T.C.; Suflita, J.M.; Morris, P.J. Metagenomic analysis and metabolite profiling of deep–sea sediments from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 2013, 4, 50.
- Prince, R.C.; Nash, G.W.; Hill, S.J. The biodegradation of crude oil in the deep ocean. Mar. Pollut. Bull. 2016, 111, 354–357.
- Aulenta, F.; Palma, E.; Marzocchi, U.; Cruz Viggi, C.; Rossetti, S.; Scoma, A. Enhanced Hydrocarbons Biodegradation at Deep-Sea Hydrostatic Pressure with Microbial Electrochemical Snorkels. Catalysts 2021, 11, 263.
- Margesin, R.; Schinner, F. Biodegradation and bioremediation of hydrocarbons in extreme environments. Appl. Microbiol. Biotechnol. 2001, 56, 650–663.
- Louvado, A.; Gomes, N.; Simões, M.M.; Almeida, A.; Cleary, D.F.; Cunha, A. Polycyclic aromatic hydrocarbons in deep sea sediments: Microbe–pollutant interactions in a remote environment. Sci. Total Environ. 2015, 526, 312–328.
- Evans, M.; Liu, J.; Bacosa, H.; Rosenheim, B.E.; Liu, Z. Petroleum hydrocarbon persistence following the Deepwater Horizon oil spill as a function of shoreline energy. Mar. Pollut. Bull. 2017, 115, 47–56.
- Michel, J.; Hayes, M.O. Weathering patterns of oil residues eight years after the Exxon Valdez oil spill. Mar. Pollut. Bull. 1999, 38, 855–863.
- Sauer, T.C.; Michel, J.; Hayes, M.O.; Aurand, D.V. Hydrocarbon characterization and weathering of oiled intertidal sediments along the Saudi Arabian Coast two years after the Gulf War oil spill. Environ. Int. 1998, 24, 43–60.
- Peterson, C.H.; Rice, S.D.; Short, J.W.; Esler, D.; Bodkin, J.L.; Ballachey, B.E.; Irons, D.B. Long-term ecosystem response to the Exxon Valdez oil spill. Science 2003, 302, 2082–2086.
- Owens, E.H.; Taylor, E.; Humphrey, B. The persistence and character of stranded oil on coarse-sediment beaches. Mar. Pollut. Bull. 2008, 56, 14–26.
- Hayes, M.O.; Gundlach, E.R.; D‘Ozouville, L. Role of dynamic coastal processes in the impact and dispersal of the Amoco Cadiz oil spill (March 1978) Brittany, France. In International Oil Spill Conference; American Petroleum Institute: Washington, DC, USA, 1979; pp. 193–198.
- Hayes, M.O. An exposure index for oiled shorelines. Spill Sci. Technol. Bull. 1996, 3, 139–147.
- Pendergraft, M.A.; Rosenheim, B.E. Varying relative degradation rates of oil in different forms and environments revealed by ramped pyrolysis. Environ. Sci. Technol. 2014, 48, 10966–10974.
- Rashid, M. Degradation of Bunker C oil under different coastal environments of Chedabucto Bay, Nova Scotia. Estuar. Coast. Mar. Sci. 1974, 2, 137–144.
- Cui, F.; Daskiran, C.; King, T.; Robinson, B.; Lee, K.; Katz, J.; Boufadel, M.C. Modeling oil dispersion under breaking waves. Part I: Wave hydrodynamics. Environ. Fluid Mech. 2020, 20, 1527–1551.
- Tkalich, P.; Chan, E.S. Vertical mixing of oil droplets by breaking waves. Mar. Pollut. Bull. 2002, 44, 1219–1229.
- Adzigbli, L.; Bacosa, H.P.; Deng, Y. Response of microbial communities to oil spill in the Gulf of Mexico: A review. Afr. J. Microbiol. Res. 2018, 12, 536–545.
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Lee, K.; Greer, C.W. Chemical dispersants enhance the activity of oil-and gas condensate-degrading marine bacteria. ISME J. 2017, 11, 2793–2808.
- Chaerun, S.K.; Tazaki, K.; Asada, R.; Kogure, K. Interaction between clay minerals and hydrocarbon-utilizing indigenous microorganisms in high concentrations of heavy oil: Implications for bioremediation. Clay Miner. 2005, 40, 105–114.
- Varjani, S.J.; Upasani, V.N. A new look on factors affecting microbial degradation of petroleum hydrocarbon pollutants. Int. Biodeterior. Biodegrad. 2017, 120, 71–83.
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 2018, 9, 2885.
- Deppe, U.; Richnow, H.-H.; Michaelis, W.; Antranikian, G. Degradation of crude oil by an arctic microbial consortium. Extremophiles 2005, 9, 461–470.
- Bacosa, H.P.; Inoue, C. Heavy oil degrading Burkholderia and Pseudomonas strains: Insights on the degradation potential of isolates and microbial consortia. Palawan Sci. 2020, 12, 74–89.
- Tang, X.; He, L.; Tao, X.; Dang, Z.; Guo, C.; Lu, G.; Yi, X. Construction of an artificial microalgal-bacterial consortium that efficiently degrades crude oil. J. Hazard. Mater. 2010, 181, 1158–1162.
- Pelz, O.; Tesar, M.; Wittich, R.M.; Moore, E.R.; Timmis, K.N.; Abraham, W.R. Towards elucidation of microbial community metabolic pathways: Unravelling the network of carbon sharing in a pollutant-degrading bacterial consortium by immunocapture and isotopic ratio mass spectrometry. Environ. Microbiol. 1999, 1, 167–174.
- Fitzpatrick, F.A.; Boufadel, M.C.; Johnson, R.; Lee, K.; Graan, T.P.; Bejarano, A.C.; Zhu, Z.; Waterman, D.; Capone, D.M.; Hayter, E. Oil-Particle Interactions and Submergence from Crude Oil Spills in Marine and Freshwater Environments: Review of the Science and Future Science Needs; U.S. Geological Survey: Reston, VA, USA, 2015.
- Adhikari, P.; Maiti, K.; Bam, W. Fate of particle-bound polycyclic aromatic hydrocarbons in the river-influenced continental margin of the northern Gulf of Mexico. Mar. Pollut. Bull. 2019, 141, 350–362.
- Adhikari, P.L.; Maiti, K.; Overton, E.B. Vertical fluxes of polycyclic aromatic hydrocarbons in the northern Gulf of Mexico. Mar. Chem. 2015, 168, 60–68.
- Adhikari, P.L.; Maiti, K.; Bosu, S.; Jones, P.R. 234Th as a tracer of vertical transport of polycyclic aromatic hydrocarbons in the northern Gulf of Mexico. Mar. Pollut. Bull. 2016, 107, 179–187.
- Bragg, J.R.; Owens, E.H. Shoreline cleansing by interactions between oil and fine mineral particles. In International Oil Spill Conference; American Petroleum Institute: Columbia, WA, USA, 1995; pp. 219–227.
- Lee, K.; Weise, A.M.; St-Pierre, S. Enhanced oil biodegradation with mineral fine interaction. Spill Sci. Technol. Bull. 1996, 3, 263–267.
- Owens, E.H. The interaction of fine particles with stranded oil. Pure Appl. Chem. 1999, 71, 83–93.
- Daly, K.L.; Passow, U.; Chanton, J.; Hollander, D. Assessing the impacts of oil-associated marine snow formation and sedimentation during and after the Deepwater Horizon oil spill. Anthropocene 2016, 13, 18–33.
- Warr, L.N.; Friese, A.; Schwarz, F.; Schauer, F.; Portier, R.J.; Basirico, L.M.; Olson, G.M. Bioremediating oil spills in nutrient poor ocean waters using fertilized clay mineral flakes: Some experimental constraints. Biotechnol. Res. Int. 2013, 2013, 704806.
- Loh, A.; Yim, U.H. A review of the effects of particle types on oil-suspended particulate matter aggregate formation. Ocean Sci. J. 2016, 51, 535–548.
- Wang, W.; Zheng, Y.; Li, Z.; Lee, K. PIV investigation of oil–mineral interaction for an oil spill application. Chem. Eng. J. 2011, 170, 241–249.
- Giani, M.; Rinaldi, A.; Degobbis, D. Mucilages in the Adriatic and Tyrrhenian Sea: An introduction. Sci. Total Environ. 2005, 353, 3–9.
- Alldredge, A.L.; Silver, M.W. Characteristics, dynamics and significance of marine snow. Prog. Oceanogr. 1988, 20, 41–82.
- Patton, J.S.; Rigler, M.W.; Boehm, P.D.; Fiest, D.L. Ixtoc 1 oil spill: Flaking of surface mousse in the Gulf of Mexico. Nature 1981, 290, 235–238.
- Dell’Amore, C. “Sea Snot” Explosion Caused by Gulf Oil Spill? 2010. Available online: https://www.nationalgeographic.com/science/article/100916-sea-snot-gulf-bp-oil-spill-marine-snow-science-environment (accessed on 30 January 2022).
- Ramachandran, S.D.; Sweezey, M.J.; Hodson, P.V.; Boudreau, M.; Courtenay, S.C.; Lee, K.; King, T.; Dixon, J.A. Influence of salinity and fish species on PAH uptake from dispersed crude oil. Mar. Pollut. Bull. 2006, 52, 1182–1189.
- Thavasi, R.; Jayalakshmi, S.; Balasubramanian, T.; Banat, I.M. Effect of salinity, temperature, pH and crude oil concentration on biodegradation of crude oil by Pseudomonas aeruginosa. J. Biol. Environ. Sci. 2007, 1, 51–57.
- Si-Zhong, Y.; Hui-Jun, J.; Zhi, W.; Rui-Xia, H.; Yan-Jun, J.; Xiu-Mei, L.; Shao-Peng, Y. Bioremediation of oil spills in cold environments: A review. Pedosphere 2009, 19, 371–381.
- Cao, Y.; Zhang, B.; Zhu, Z.; Song, X.; Cai, Q.; Chen, B.; Dong, G.; Ye, X. Microbial eco-physiological strategies for salinity-mediated crude oil biodegradation. Sci. Total Environ. 2020, 727, 138723.
- Jamal, M.T.; Pugazhendi, A. Degradation of petroleum hydrocarbons and treatment of refinery wastewater under saline condition by a halophilic bacterial consortium enriched from marine environment (Red Sea), Jeddah, Saudi Arabia. 3 Biotech 2018, 8, 276.
- Duran, R.; Cravo-Laureau, C. Role of environmental factors and microorganisms in determining the fate of polycyclic aromatic hydrocarbons in the marine environment. FEMS Microbiol. Rev. 2016, 40, 814–830.
- Dong, C.; Bai, X.; Sheng, H.; Jiao, L.; Zhou, H.; Shao, Z. Distribution of PAHs and the PAH-degrading bacteria in the deep-sea sediments of the high-latitude Arctic Ocean. Biogeosciences 2015, 12, 2163–2177.
- Al-Awadhi, H.; Dashti, N.; Kansour, M.; Sorkhoh, N.; Radwan, S. Hydrocarbon-utilizing bacteria associated with biofouling materials from offshore waters of the Arabian Gulf. Int. Biodeterior. Biodegrad. 2012, 69, 10–16.
- Somee, M.R.; Shavandi, M.; Dastgheib, S.M.M.; Amoozegar, M.A. Bioremediation of oil-based drill cuttings by a halophilic consortium isolated from oil-contaminated saline soil. 3 Biotech 2018, 8, 229.
- Mnif, S.; Chamkha, M.; Sayadi, S. Isolation and characterization of Halomonas sp. strain C2SS100, a hydrocarbon-degrading bacterium under hypersaline conditions. J. Appl. Microbiol. 2009, 107, 785–794.
- Zhao, B.; Wang, H.; Mao, X.; Li, R. Biodegradation of phenanthrene by a halophilic bacterial consortium under aerobic conditions. Curr. Microbiol. 2009, 58, 205–210.
- Dastgheib, S.M.M.; Amoozegar, M.A.; Khajeh, K.; Shavandi, M.; Ventosa, A. Biodegradation of polycyclic aromatic hydrocarbons by a halophilic microbial consortium. Appl. Microbiol. Biotechnol. 2012, 95, 789–798.
- Al-Mailem, D.; Sorkhoh, N.; Al-Awadhi, H.; Eliyas, M.; Radwan, S. Biodegradation of crude oil and pure hydrocarbons by extreme halophilic archaea from hypersaline coasts of the Arabian Gulf. Extremophiles 2010, 14, 321–328.
- Khemili-Talbi, S.; Kebbouche-Gana, S.; Akmoussi-Toumi, S.; Angar, Y.; Gana, M.L. Isolation of an extremely halophilic arhaeon Natrialba sp. C21 able to degrade aromatic compounds and to produce stable biosurfactant at high salinity. Extremophiles 2015, 19, 1109–1120.
- Okpokwasili, G.; Odokuma, L. Effect of salinity on biodegradation of oil spill dispersants. Waste Manag. 1990, 10, 141–146.
- Sun, J.; Zheng, X. A review of oil-suspended particulate matter aggregation—a natural process of cleansing spilled oil in the aquatic environment. J. Environ. Monit. 2009, 11, 1801–1809.
- Wells, P.G.; Butler, J.N.; Hughes, J.S. Exxon Valdez Oil Spill: Fate and Effects in Alaskan Waters; ASTM: West Conshohocken, PA, USA, 1995.
- Díaz, M.P.; Boyd, K.G.; Grigson, S.J.; Burgess, J.G. Biodegradation of crude oil across a wide range of salinities by an extremely halotolerant bacterial consortium MPD-M, immobilized onto polypropylene fibers. Biotechnol. Bioeng. 2002, 79, 145–153.
- Megharaj, M.; Venkateswarlu, K.; Naidu, R. Bioremediation. In Encyclopedia of Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1, pp. 485–489.
- Megharaj, M.; Naidu, R. Soil and brownfield bioremediation. Microb. Biotechnol. 2017, 10, 1244–1249.
- Kuppusamy, S.; Thavamani, P.; Megharaj, M.; Lee, Y.B.; Naidu, R. Isolation and characterization of polycyclic aromatic hydrocarbons (PAHs) degrading, pH tolerant, N-fixing and P-solubilizing novel bacteria from manufactured gas plant (MGP) site soils. Environ. Technol. Innov. 2016, 6, 204–219.
- Piontek, J.; Lunau, M.; Händel, N.; Borchard, C.; Wurst, M.; Engel, A. Acidification increases microbial polysaccharide degradation in the ocean. Biogeosciences 2010, 7, 1615–1624.
- Radwan, S.; Al-Hasan, R.; Ali, N.; Salamah, S.; Khanafer, M. Oil-consuming microbial consortia floating in the Arabian Gulf. Int. Biodeterior. Biodegrad. 2005, 56, 28–33.
- Hui, L.; Zhang, Y.; Kravchenko, I.; Hui, X.; Zhang, C.-g. Dynamic changes in microbial activity and community structure during biodegradation of petroleum compounds: A laboratory experiment. J. Environ. Sci. 2007, 19, 1003–1013.
- Okoh, A.I. Biodegradation alternative in the cleanup of petroleum hydrocarbon pollutants. Biotechnol. Mol. Biol. Rev. 2006, 1, 38–50.
- Fathepure, B.Z. Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environments. Front. Microbiol. 2014, 5, 173.
- Pawar, R.M. The effect of soil pH on bioremediation of polycyclic aromatic hydrocarbons (PAHS). J. Bioremediat. Biodegrad. 2015, 6, 291–304.
- Obahiagbon, K.O.; Amenaghawon, A.N.; Agbonghae, E.O. Effect of initial pH on the bioremediation of crude oil polluted water using a consortium of microbes. Pasific J. Sci. Technol. 2014, 15, 452–457.
More
Information
Subjects:
Environmental Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
5.0K
Revisions:
3 times
(View History)
Update Date:
06 Apr 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No