 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Larisa Koleva | + 4679 word(s) | 4679 | 2022-03-17 04:09:04 | | | |
| 2 | Peter Tang | Meta information modification | 4679 | 2022-03-23 07:54:18 | | | | |
| 3 | Peter Tang | Meta information modification | 4679 | 2022-03-23 07:56:04 | | |
Video Upload Options
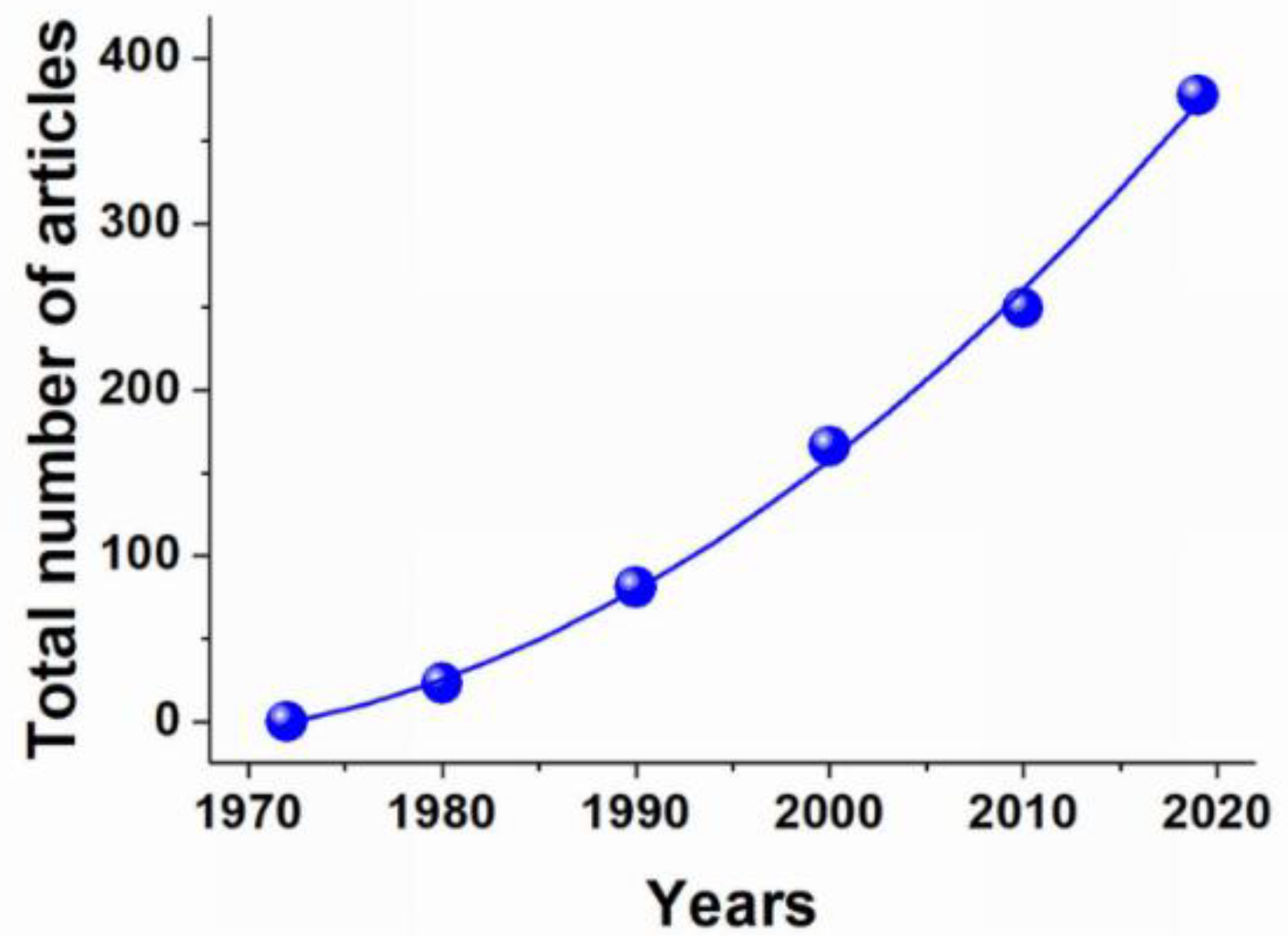
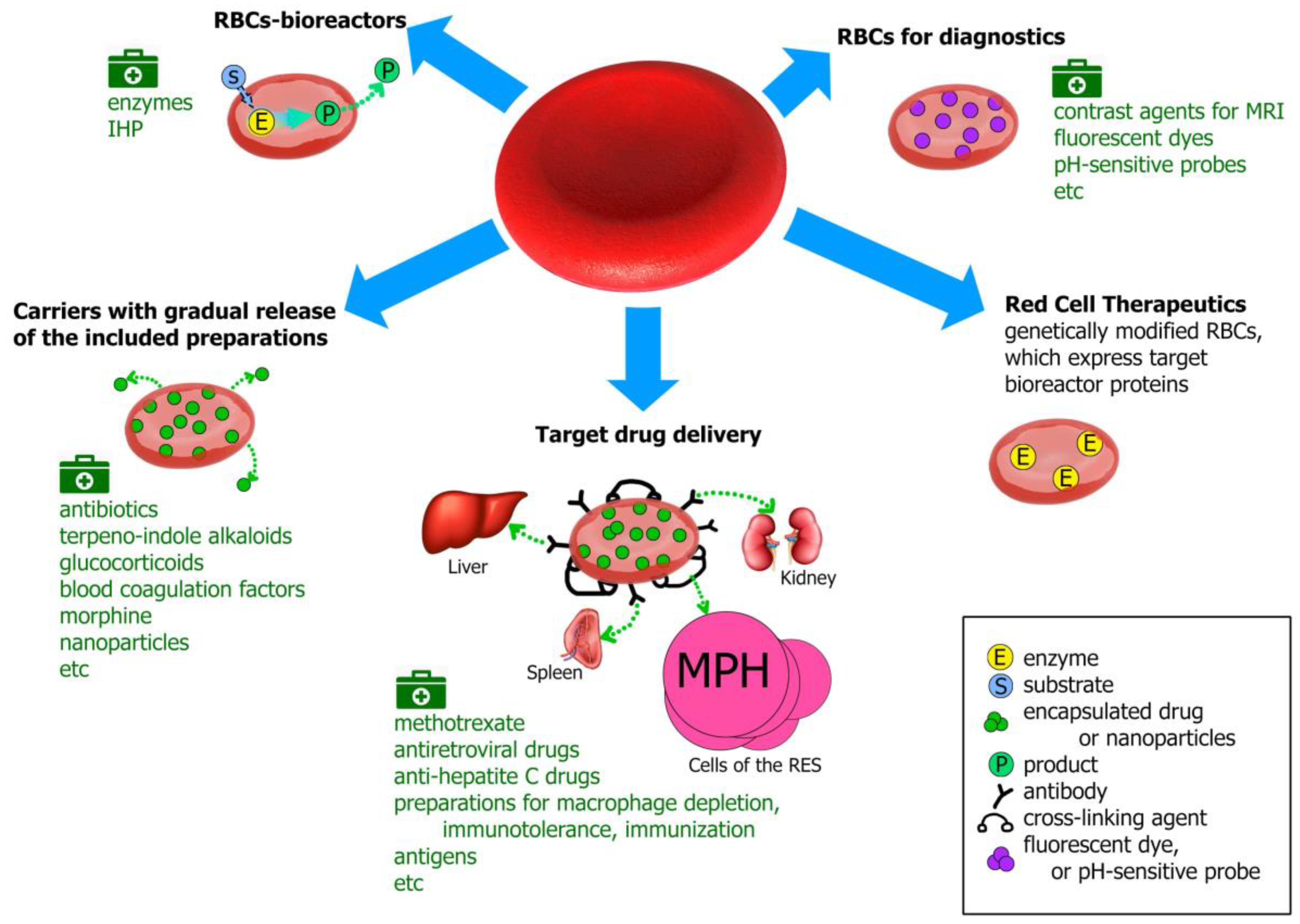
Erythrocytes (red blood cells, RBCs) are the largest population of blood cells in mammals. Their main function is oxygen transfer to cells and body tissues. The lifetime of erythrocytes in the bloodstream is 100–120 days, after which they are removed by the spleen. Due to the unique biophysical properties RBCs can be used as drug carriers in two different ways: by incorporating the drug into the cells or by binding it (using non-specific adsorption or a specific association, involving antibodies or various chemical cross-linking compounds) on the RBCs’ surface. Erythrocytes can act as carriers that prolong the drug’s action due to its gradual release from the carrier; as bioreactors with encapsulated enzymes performing the necessary reactions, while remaining inaccessible to the immune system and plasma proteases; or as a tool for targeted drug delivery to target organs, primarily to cells of the reticuloendothelial system, liver and spleen. To date, erythrocytes have been studied as carriers for a wide range of drugs, such as enzymes, antibiotics, anti-inflammatory, antiviral drugs, etc., and for diagnostic purposes.
1. Erythrocytes as Drug Carriers
- - biocompatibility (human, both autologous and donor erythrocytes are used to treat patients);
- - biodegradability (old or damaged erythrocytes are naturally removed by the reticuloendothelial system);
- - long life in the bloodstream (the drug has an extended lifetime inside the cells because RBCs protect it from the immune system and plasma proteases and the cells survive in the body for a long time; thus, the pharmacokinetics and pharmacodynamics of the drug in RBCs can significantly increase the desired therapeutic effect);
- - decreasing side effects of drugs (due to preventing allergic reactions, and the decrease in the peak concentrations of free drug in the blood to safer levels);
- - ease of cell isolation in large quantities and the ability to scale production.
2. Erythrocytes for Targeted Drug Delivery
2.1. Methotrexate
2.2. Erythrocytes-Carriers for Treatment of Retroviral Infection
2.3. Drugs Loaded into RBCs for the Treatment of Hepatitis C
2.4. Macrophage Depletion
2.5. Antigens Loaded into Erythrocytes or Associated with Their Surface
2.5.1. Immunization
2.5.2. Cancer Immunotherapy
2.5.3. Induction of Immune Tolerance
References
- Ataullakhanov, F.I.; Borsakova, D.V.; Protasov, E.S.; Sinauridze, E.I.; Zeynalov, A.M. Erythrocyte: A bag with hemoglobin, or a living active cell? Pediatrics Hematol. Oncol. Immunopathol. 2018, 17, 108–116, (In Russian, abstract in English).
- Muzykantov, V.R.; Murciano, J.C.; Taylor, R.P.; Atochina, E.N.; Herraez, A. Regulation of the complement-mediated elimination of red blood cells modified with biotin and streptavidin. Anal. Biochem. 1996, 241, 109–119.
- Muzykantov, V.R.; Zaltsman, A.B.; Smirnov, M.D.; Samokhin, G.P.; Morgan, B.P. Target-sensitive immunoerythrocytes: Interaction of biotinylated red blood cells with immobilized avidin induces their lysis by complement. Biochim. Biophys. Acta (Biomembranes) 1996, 1279, 137–143.
- Murciano, J.C.; Medinilla, S.; Eslin, D.; Atochina, E.; Cines, D.B.; Muzykantov, V.R. Prophylactic fibrinolysis through selective dissolution of nascent clots by tPA-carrying erythrocytes. Nat. Biotechnol. 2003, 21, 891–896.
- Murciano, J.C.; Muzykantov, V.R. Coupling of anti-thrombotic agents to red blood cells offers safer and more effective management of thrombosis. Discov. Med. 2003, 3, 28–29.
- Ji, W.; Smith, P.N.; Koepse, R.R.; Andersen, J.D.; Baker, S.L.; Zhang, L.; Carmali, S.; Myerson, J.W.; Muzykantov, V.; Russell, A.J. Erythrocytes as carriers of immunoglobulin-based therapeutics. Acta Biomater. 2020, 101, 422–435.
- Muzykantov, V.R.; Murciano, J.-C. Streptavidin-mediated coupling of therapeutic proteins to carrier erythrocytes. In Erythrocyte Engineering for Drug Delivery and Targeting. Biotechnology Intelligence; Unit 6; Magnani, M., Ed.; Landes Bioscience: Georgetown, TX, USA, 2002; Chapter 4; pp. 37–67. ISBN 1-58706-061-2.
- Villa, C.H.; Pan, D.C.; Zaitsev, S.; Cines, D.B.; Siegel, D.L.; Muzykantov, V.R. Delivery of drugs bound to erythrocytes: New avenues for an old intravascular carrier. Ther. Deliv. 2015, 6, 795–826.
- Villa, C.H.; Anselmo, A.C.; Mitragotri, S.; Muzykantov, V. Red blood cells: Supercarriers for drugs, biologicals, and nanoparticles and inspiration for advanced delivery systems. Adv. Drug Deliv. Rev. 2016, 106, 88–103.
- Tsong, T.Y.; Kinosita, K., Jr. Use of voltage pulses for the pore opening and drug loading and the subsequent resealing of red blood cells. In Red Blood Cells as Carriers for Drugs; Karger Publishers: Berlin, Germany, 1985; pp. 108–114.
- Lizano, C.; Sanz, S.; Luque, J.; Pinilla, M. In vitro study of alcohol dehydrogenase and acetaldehyde dehydrogenase encapsulated into human erythrocytes by an electroporation procedure. Biochim. Biophys. Acta (Gen. Subj.) 1998, 1425, 328–336.
- Yamagata, K.; Kawasaki, E.; Kawarai, H.; Iino, M. Encapsulation of concentrated protein into erythrocyte porated by continuous-wave ultrasound. Ultrasound. Med. Biol. 2008, 34, 1924–1933.
- Ginn, F.L.; Hochstein, P.; Trump, B.F. Membrane alterations in hemolysis: Internalization of plasmalemma induced by primaquine. Science 1969, 164, 843–845.
- Ben-Bassat, I.; Bensch, K.G.; Schrier, S.L. Drug-induced erythrocyte membrane internalization. J. Clin. Investig. 1972, 51, 1833–1844.
- Matovcik, L.M.; Junga, I.G.; Schrie, S.L. Drug-induced endocytosis of neonatal erythrocytes. Blood 1985, 65, 1056–1063. Available online: https://ashpublications.org/blood/article/65/5/1056/164002/Drug-induced-endocytosis-of-neonatal-erythrocytes (accessed on 10 March 2020).
- Kwon, Y.M.; Chung, H.S.; Moon, C.; Yockman, J.; Park, Y.J.; Gitlin, S.D.; David, A.E.; Yang, V.C. l-Asparaginase encapsulated intact erythrocytes for treatment of acute lymphoblastic leukemia (ALL). J. Control. Release 2009, 139, 182–189.
- Franco, R.S.; Weiner, M.; Wagner, K.; Martelo, O.J. Incorporation of inositol hexaphosphate into red blood cells mediated by dimethyl sulfoxide. Life Sci. 1983, 32, 2763–2768.
- Mosca, A.; Paleari, R.; Russo, V.; Rosti, E.; Nano, R.; Boicelli, A.; Villa, S.; Zanella, A. IHP entrapment into human erythrocytes: Comparison between hypotonic dialysis and DMSO osmotic pulse. Adv. Exp. Med. Biol. 1992, 326, 19–26.
- Wang, X.; Ge, W.; Xu, X.; Kang, X.; Luo, X. Investigation on the preparative method of morphine loaded in erythrocyte and encapsulating effect of carrier erythrocytes. Chin. J. Clin. Pharm. 2003, 06, 335–338, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-LCZZ200306003.htm (accessed on 20 February 2020).
- Ge, W.-h.; Lian, Y.-s.; Wang, X.-h.; Luo, X.; Xie, P.-h. Morphological observation of erythrocyte during the preparation of morphine carrier by a hyperosmotic method. Chin. Pharm. J. 2004, 04, 270–272, (In Chinese, Abstract in English). Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-ZGYX200404011.htm (accessed on 20 February 2020).
- Ihler, G.M.; Glew, R.H.; Schnure, F.W. Enzyme loading of erythrocytes. Proc. Natl. Acad. Sci. USA 1973, 70, 2663–2666.
- Updike, S.J.; Wakamiya, R.T. Infusion of red blood cell-loaded asparaginase in monkey. Immunologic, metabolic, and toxicologic consequences. J. Lab. Clin. Med. 1983, 101, 679–691.
- Rechsteiner, M.C. Uptake of proteins by red blood cells. Exp. Cell Res. 1975, 93, 487–492.
- Alpar, H.O.; Lewis, D.A. Therapeutic efficacy of asparaginase encapsulated in intact erythrocytes. Biochem. Pharmacol. 1985, 34, 257–261.
- Magnani, M.; Rossi, L.; D’ascenzo, M.; Panzani, I.; Bigi, L.; Zanella, A. Erythrocyte engineering for drug delivery and targeting. Biotechnol. Appl. Biochem. 1998, 28, 1–6.
- DeLoach, J.R.; Ihler, G. A dialysis procedure for loading erythrocytes with enzymes and lipids. Biochim. Biophys. Acta (Gener. Sub.) 1977, 496, 136–145.
- Bax, B.E.; Bain, M.D.; Fairbanks, L.D.; Webster, A.D.; Chalmers, R.A. In vitro and in vivo studies with human carrier erythrocytes loaded with polyethylene glycol-conjugated and native adenosine deaminase. Br. J. Haematol. 2000, 109, 549–554.
- DeLoach, J.R.; Harris, R.L.; Ihler, G.M. An erythrocyte encapsulator dialyzer used in preparing large quantities of erythrocyte ghosts and encapsulation of a pesticide in erythrocyte ghosts. Anal. Biochem. 1980, 102, 220–227.
- Ropars, C.; Nicolau, C.; Chassaigne, M. Process and Device for the Encapsulation in Erythrocytes of at Least One Biologically Active Substance, in Particular Hemoglobin Allosteric Effectors, and Erythrocytes So Obtained. EP 0101341. Data of Publication: 1 June 1983. Available online: https://europepmc.org/article/pat/ep0101341 (accessed on 10 March 2020).
- Godfrin, Y. Lysis/Resealing Process and Device for Incorporating an Active Ingredient, in Particular Asparaginase or Inositol Hexaphosphate, in Erythrocytes. US Patent 2008261262, 4 August 2008. Available online: https://europepmc.org/article/pat/us2008261262 (accessed on 10 March 2020).
- Millan, C.G.; Marinero, M.L.S.; Castaneda, A.Z.; Lanao, J.M. Drug, enzyme and peptide delivery using erythrocytes as carriers. J. Control Release 2004, 95, 27–49.
- Pierige, F.; Serafini, S.; Rossi, L.; Magnani, M. Cell-based drug delivery. Adv. Drug Deliv. Rev. 2008, 60, 286–295.
- Milanick, M.A.; Ritter, S.; Meissner, K. Engineering erythrocytes to be erythrosensors: First steps. Blood Cells Mol. Dis. 2011, 47, 100–106.
- Ritter, S.C.; Milanick, M.A.; Meissner, K.E. Encapsulation of FITC to monitor extracellular pH: A step towards the development of red blood cells as circulating blood analyte biosensors. Biomed. Opt. Express 2011, 2, 2012–2021.
- Ritter, S.C.; Shao, X.; Cooley, N.; Milanick, M.A.; Glass, T.E.; Meissner, K.E. Blood analyte sensing using fluorescent dye-loaded red blood cells. In Proceedings of the Optical Diagnostics and Sensing XIV: Toward Point-of-Care Diagnostics, San Francisco, CA, USA, 3–6 February 2014; Coté, G.L., Ed.; SPIE: San Francisco, CA, USA, ; 2014; Volume 8951.
- Pierigè, F.; Bigini, N.; Rossi, L.; Magnani, M. Reengineering red blood cells for cellular therapeutics and diagnostics. WIREs Nanomed. Nanobiotechnol. 2017, 9, e1454:1–e1454:17.
- Eichler, H.G.; Gasic, S.; Bauer, K.; Korn, A.; Bacher, S. In vivo clearance of antibody-sensitized human drug carrier erythrocytes. Clin. Pharmacol. Ther. 1986, 40, 300–303.
- Delaby, C.; Pilard, N.; Hetet, G.; Driss, F.; Grandchamp, B.; Beaumont, C.; Canonne-Hergaux, F. A physiological model to study iron recycling in macrophages. Exp. Cell Res. 2005, 310, 43–53.
- Chiarantini, L.; Rossi, L.; Fraternale, A.; Magnani, M. Modulated red blood cell survival by membrane protein clustering. Mol. Cell. Biochem. 1995, 144, 53–59.
- Bratosin, D.; Mazurier, J.; Tissier, J.P.; Slomianny, C.; Estaquier, J.; Russo-Marie, F.; Huart, J.J.; Freyssinet, J.M.; Aminoff, D.; Ameisen, J.C.; et al. Molecular mechanisms of erythrophagocytosis. Characterization of the senescent erythrocytes that are phagocytized by macrophages. C. R. Acad. Sci. III. Sciences de la vie/Life Sciences 1997, 320, 811–818.
- Magnani, M.; Rossi, L.; Brandit, G.; Schiavano, G.F.; Montroni, M.; Piedimonte, G. Targeting antiretroviral nucleoside analogues in phosphorylated form to macrophages : In vitro and in vivo studies. Proc. Nati. Acad. Sci. USA 1992, 89, 6477–6481.
- Chiarantini, L.; Antonelli, A.; Rossi, L.; Fraternale, A.; Magnani, M. Red blood cell phagocytosis following hexokinase inactivation. CELL Biochem. Funct. 1994, 12, 217–220.
- Zimmermann, U.; Pilwat, G.; Esser, B. The effect of encapsulation in red blood cells on the distribution of methotrexate in mice. Clin. Chem. Lab. Med. 1978, 16, 135–144.
- DeLoach, J.R.; Barton, C. Glutaraldehyde-treated carrier erythrocytes for organ targeting of methotrexate in dogs. Am. J. Vet. Res. 1981, 42, 1971–1974.
- DeLoach, J.R.; Tangner, C.H.; Barton, C. Hepatic pharmacokinetics of glutaraldehyde-treated methotrexate-loaded carrier erythrocytes in dogs. Res. Exp. Med. 1983, 183, 167–175.
- Yuan, S.-H.; Ge, W.-H.; Huo, J.; Wang, X.-H. Slow release properties and liver-targeting characteristics of methotrexate erythrocyte carriers. Fundam. Clin. Pharmacol. 2009, 23, 189–196.
- Mishra, P.R.; Jain, N.K. Biotinylated methotrexate loaded erythrocytes for enhanced liver uptake. ‘A study on the rat’. Int. J. Pharm. 2002, 231, 145–153.
- Mishra, P.R.; Jain, N.K. Surface modified methotrexate loaded erythrocytes for enhanced macrophage uptake. J. Drug Target. 2000, 8, 217–224.
- Perno, C.F.; Yarcoan, R.; Cooney, D.A.; Hartman, N.R.; Gartner, G.; Popovich, M.; Hao, Z.; Gerrard, T.L.; Wilson, Y.A. Inhibition of human immunodeficiency virus (HIV-1/HTLV-IIIBa-L) replication in fresh and cultured human peripheral blood monocytes/macrophages by azidothymidine and related 2’,3’-dideoxynucleosides. J. Exp. Med. 1988, 168, 1111–1125.
- Fraternale, A.; Tonelli, A.; Casabianca, A.; Vallanti, G.; Chiarantini, L.; Schiavano, G.F.; Benatti, U.; De Flora, A.; Magnani, M. Role of macrophage protection in the development of murine AIDS. J. Acquir. Immune Defic. Syndr. 1999, 21, 81–89.
- Fraternale, A.; Casabianca, A.; Rossi, L.; Chiarantini, L.; Schiavano, G.F.; Palamara, A.T.; Garaci, E.; Magnani, M. Erythrocytes as carriers of reduced glutathione (GSH) in the treatment of retroviral infections. J. Antimicrob. Chemother. 2003, 52, 551–554.
- Rossi, L.; Casabianca, A.; Fraternale, A.; Schiavano, G.F.; Brandi, G.; Antonelli, A.; Magnani, M. Macrophage protection by nucleoside and nucleotide analogue administration. In Erythrocytes as Drug Carriers in Medicine; Sprandel, U., Way, J.L., Eds.; Springer: Boston, MA, USA, 1997; pp. 63–71. ISBN 978-1-4899-0046-3.
- Magnani, M.; Rossi, L.; Fraternale, A.; Casabianca, A.; Brandi, G.; Benatti, U.; De Flora, A. Targeting antiviral nucleotide analogues to macrophages. J. Leukoc. Biol. 1997, 62, 133–137.
- Magnani, M.; Rossi, L.; Fraternale, A.; Silvotti, L.; Quintavalla, F.; Piedimonte, G.; Matteucci, D.; Baldinotti, F.; Bendinelli, M. Feline immunodeficiency virus infection of macrophages: In vitro and in vivo inhibition by dideoxycytidine-5′-triphosphate-loaded erythrocytes. AIDS Res. Hum. Retrovir. 1994, 10, 1179–1186.
- Rossi, L.; Brandi, G.; Fraternale, A.; Schiavano, G.F.; Chiarantini, L.; Magnani, M. Inhibition of murine retrovirus-induced immunodeficiency disease by dideoxycytidine and dideoxycytidine 5′-triphosphate. J. Acquir. Immune Defic. Syndr. 1993, 6, 1179–1186.
- Magnani, M.; Rossi, L.; Fraternale, A.; Silvotti, L.; Quintavalla, F.; Piedimonte, G.; Matteucci, D.; Baldinotti, F.; Bendinelli, M. FIV infection of macrophages: In vitro and in vivo inhibition by dideoxycytidine 5′-triphosphate. Vet. Immunol. Immunopathol. 1995, 46, 151–158.
- Benatti, U.; Giovine, M.; Damonte, G.; Gasparini, A.; Scarfi, S.; De Flora, A.; Fraternale, A.; Rossi, L.; Magnani, M. Azidothymidine homodinucleotide-loaded erythrocytes as bioreactors for slow delivery of the antiretroviral drug azidothymidine. Biochem. Biophys. Res. Commun. 1996, 220, 20–25.
- Magnani, M.; Fraternale, A.; Casabianca, A.; Schiavano, G.F.; Chiarantini, L.; Palamara, A.T.; Chiriolo, M.R.; Rotilio, G.; Garaci, E. Antiretroviral effect of combined zidovudine and reduced glutathione therapy in murine AIDS. AIDS Res. Hum. Retroviruses 1997, 13, 1093–1099.
- Fraternale, A.; Casabianca, A.; Orlandi, C.; Cerasi, A.; Chiarantini, L.; Brandi, G.; Magnani, M. Macrophage protection by addition of glutathione (GSH)-loaded erythrocytes to AZT and DDI in a murine AIDS model. Antiviral Res. 2002, 56, 263–272.
- Magnani, M.; Balestra, E.; Fraternale, A.; Aquaro, S.; Paiardini, M.; Cervasi, B.; Casabianca, A.; Garaci, E.; Perno, C.-F. Drug-loaded red blood cell-mediated clearance of HIV-1 macrophage reservoir by selective inhibition of STAT1 expression. J. Leukoc. Biol. 2003, 74, 764–771.
- Franco, R.; Dufour, E.; Kosenko, E.; Bax, B.E.; Banz, A.; Skorokhod, O.A.; Lanao, M.; Vitvitsky, V.; Sinauridze, E.; Bourgeaux, V.; et al. International seminar on the red blood cells as vehicles for drugs. Expert Opin. Biol. Ther. 2012, 12, 127–133.
- Foroozesh, M.; Zarrin, A. A novel combinatory paradigm for chronic hepatitis C treatment using liver-targeted carrier erythrocytes co-encapsulated with inter-feron alpha-2b, ribavirin and boceprevir. Irn. J. Med. Hypotheses Ideas 2010, 4, 10:1–10:8. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.921.5785&rep=rep1&type=pdf (accessed on 20 February 2020).
- Sabatino, R.; Antonelli, A.; Battistelli, S.; Schwendener, R.; Magnani, M.; Rossi, L. Macrophage depletion by free bisphosphonates and zoledronate-loaded red blood cells. PLoS ONE 2014, 9, e101260:1–e101260:12.
- Shih, J.-Y.; Yuan, A.; Chen, J.J.-W.; Yang, P.-C. Tumor-associated macrophage : Its role in cancer invasion and metastasis. J. Cancer Mol. 2006, 2, 101–106. Available online: http://www.oalib.com/paper/2766208#.Xk6P9kpn2Ul (accessed on 20 February 2020).
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073.
- Fukuda, K.; Kobayashi, A.; Watabe, K. The role of tumor-associated macrophage in tumor progression. Front. Biosci. (Schol. Ed.). 2012, 4, 787–798.
- Mac, J.T.; Nuñez, V.; Burns, J.M.; Guerrero, Y.A.; Vullev, V.I.; Anvari, B. Erythrocyte-derived nano-probes functionalized with antibodies for targeted near infrared fluorescence imaging of cancer cells. Biomed. Opt. Express 2016, 7, 1311–1322.
- Rossi, L.; Migliavacca, B.; Pierigé, F.; Serafini, S.; Sanvito, F.; Olivieri, S.; Nano, R.; Antonioli, B.; Magnani, M.; Bertuzzi, F. Prolonged islet allograft survival in diabetic mice upon macrophage depletion by clodronate-loaded erythrocytes. Transplantation 2008, 85, 648–650.
- Pozzi, L.-A.M.; Maciaszek, J.W.; Rock, K.L. Both dendritic cells and macrophages can stimulate naive CD8 T cells in vivo to proliferate, develop effector function, and differentiate into memory cells. J. Immunol. 2005, 175, 2071–2081.
- Magnani, M.; Chiarantini, L.; Vittoria, E.; Mancini, U.; Rossi, L.; Fazi, A. Red blood cells as an antigen-delivery system. Biotechnol. Appl. Biochem. 1992, 16, 188–194.
- Chiarantini, L.; Argnanit, R.; Zucchinit, S.; Stevanatot, L.; Grossi, M.P.; Magnani, M.; Manservigi, R. Red blood cells as delivery system for recombinant HSV-1 glycoprotein B: Immunogenicity and protection in mice. Vaccine 1997, 15, 276–280.
- Dominici, S.; Laguardia, M.E.; Serafini, G.; Chiarantini, L.; Fortini, C.; Tripiciano, A.; Scoglio, A.; Caputo, A.; Fiorelli, V.; Gavioli, R.; et al. Red blood cell-mediated delivery of recombinant HIV-1 Tat protein in mice induces anti-Tat neutralizing antibodies and CTL. Vaccine 2003, 21, 2073–2081.
- Polvani, C.; Gasparini, A.; Benatti, U.; DeFlora, A.; Silvestri, S.; Volpini, G.; Nencioni, L. Murine red blood cells as efficient carriers of three bacterial antigens for the production of specific and neutralizing antibodies. Biotechnol. Appl. Biochem. 1991, 14, 347–356.
- Murray, A.M.; Pearson, I.F.S.; Fairbanks, L.D.; Chalmers, R.A.; Bain, M.D.; Bax, B.E. The mouse immune response to carrier erythrocyte entrapped antigens. Vaccine 2006, 24, 6129–6139.
- Renno, T.; Lebecque, S.; Renard, N.; Saeland, S.; Vicari, A. What’s new in the field of cancer vaccines? Cell. Mol. Life Sci. 2003, 60, 1296–1310.
- Melief, C.J. Cancer immunotherapy by dendritic cells. Immunity 2008, 29, 372–383.
- Banz, A.; Cremel, M.; Rembert, A.; Godfrin, Y. In situ targeting of dendritic cells by antigen-loaded red blood cells: A novel approach to cancer immunotherapy. Vaccine 2010, 28, 2965–2972.
- Hendrickson, J.E.; Chadwick, T.E.; Roback, J.D.; Hillyer, C.D.; Zimring, J.C. Inflammation enhances consumption and presentation of transfused RBC antigens by dendritic cells. Blood 2007, 110, 2736–2743.
- Hendrickson, J.E.; Roback, J.D.; Hillyer, C.D.; Easley, K.A.; Zimring, J.C. Discrete Toll-like receptor agonists have differential effects on alloimmunization to transfused red blood cells. Transfusion 2008, 48, 1869–1877.
- Banz, A.; Cremel, M.; Mouvant, A.; Guerin, N.; Horand, F.; Godfrin, Y. Tumor growth control using red blood cells as the antigen delivery system and poly(I:C). J. Immunother. 2012, 35, 409–417.
- Cremel, M.; Guerin, N.; Campello, G.; Barthe, Q.; Berlier, W.; Horand, F.; Godfrin, Y. Innovative approach in Pompe disease therapy: Induction of immune tolerance by antigen-encapsulated red blood cells. Int. J. Pharm. 2015, 491, 69–77.
- Cremel, M.; Guérin, N.; Horand, F.; Banz, A.; Godfrin, Y. Red blood cells as innovative antigen carrier to induce specific immune tolerance. Int. J. Pharm. 2013, 443, 39–49.
- Khubutiya, M.S.; Gulyaev, V.A.; Khvatov, V.B.; Lemenev, V.L.; Kabanova, S.A.; Novruzbekov, M.S.; Lutsyk, K.N.; Olisov, O.D.; Zhuravel’, S.V.; Bulava, G.V.; et al. Immunological tolerance in organ transplantation. Transplantologiya (Russia) 2017, 9, 211–225.
- Yamazaki, S.; Dudziak, D.; Heidkamp, G.F.; Fiorese, C.; Bonito, A.J.; Inaba, K.; Nussenzweig, M.C.; Steinman, R.M. CD8+ CD205+ splenic dendritic cells are specialized to induce Foxp3+ regulatory T cells. J. Immunol. 2008, 181, 6923–6933.


