Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sergey Sedykh | + 3972 word(s) | 3972 | 2022-03-08 04:07:34 | | | |
| 2 | Yvaine Wei | + 2 word(s) | 3974 | 2022-03-22 06:49:58 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Sedykh, S. Antibodies in HIV-infection and HIV-vaccine development. Encyclopedia. Available online: https://encyclopedia.pub/entry/20804 (accessed on 08 August 2026).
Sedykh S. Antibodies in HIV-infection and HIV-vaccine development. Encyclopedia. Available at: https://encyclopedia.pub/entry/20804. Accessed August 08, 2026.
Sedykh, Sergey. "Antibodies in HIV-infection and HIV-vaccine development" Encyclopedia, https://encyclopedia.pub/entry/20804 (accessed August 08, 2026).
Sedykh, S. (2022, March 21). Antibodies in HIV-infection and HIV-vaccine development. In Encyclopedia. https://encyclopedia.pub/entry/20804
Sedykh, Sergey. "Antibodies in HIV-infection and HIV-vaccine development." Encyclopedia. Web. 21 March, 2022.
Copy Citation
The structure of HIV-envelope changes as the infection progresses and is one of the biggest obstacles in developing of HIV-vaccine. HIV-infection can cause the production of various natural autoantibodies, including catalytic antibodies hydrolyzing DNA, myelin basic protein, histones, HIV-integrase, HIV-reverse transcriptase, β-casein, serum albumin, and some other natural substrates. Currently, there are various directions for the development of HIV vaccines: stimulation of the immune response on the mucous membranes; induction of cytotoxic T cells, which lyse infected cells and hold back HIV-infection; immunization with recombinant Env proteins or vectors encoding Env; mRNA-based vaccines and some others.
HIV
antibodies
IgG
neutralizing antibodies
vaccine
HIV-vaccine
catalytic antibodies
COVID-19
1. Introduction
Since the first clinical detection of AIDS (acquired immunodeficiency syndrome) and the subsequent isolation of HIV (human immunodeficiency virus) in the early 1980s, the HIV epidemic continues to be one of the major health threats in the world, despite four decades of intensive research [1].
A high level of genetic variability in HIV is one of the biggest obstacles in developing a safe and effective vaccine. Unlike many other viruses, HIV is highly variable, with many subtypes and recombinant forms [2][3]. Low fidelity of HIV reverse transcriptase leads to the rapid generation of mutants carrying base substitutions, insertions, and deletions. Combined with the addition and loss of glycosylation sites, this results in tremendous viral diversity [4][5][6]. A key source of genetic diversity is the viral gene env, which encodes envelope glycoprotein (Env), gp120, and gp41. As is known, gp120 and gp41 subunits on the surface of the virion are trimerized, form spikes, and together are involved in penetration into the target cell. Env has a complex conformation and undergoes significant rearrangements when binding to CD4 and coreceptors [7][8][9]. Glycans make up half the mass of the HIV envelope glycoprotein, forming a topographic landscape that alters the availability of antibody (Ab) binding to the immunogen [10].
Env is also a major target for neutralizing antibodies (nAbs) [11][12]. The inability of the adaptive immune system to prevent and control infection is explained by the structural variability of the HIV envelope. Protein gp120 consists of five relatively constant (C) regions and five highly variable (V) regions. Most adaptive responses target V-domain epitopes that mutate rapidly [13]. Cytotoxic T-lymphocytes and nAbs provide only temporary protection [14][15].
Progression of HIV-infection to AIDS occurs over 1 year to more than 2 decades [16]. Some HIV-infected patients develop Abs that can neutralize a wide range of HIV strains [17][18][19]. HIV has mechanisms preventing the production of broadly neutralizing antibodies (bNAbs) (Figure 1), including intense glycosylation of Env glycoproteins, instability of these glycoproteins, and conformational masking of receptor binding sites [20][21][22].
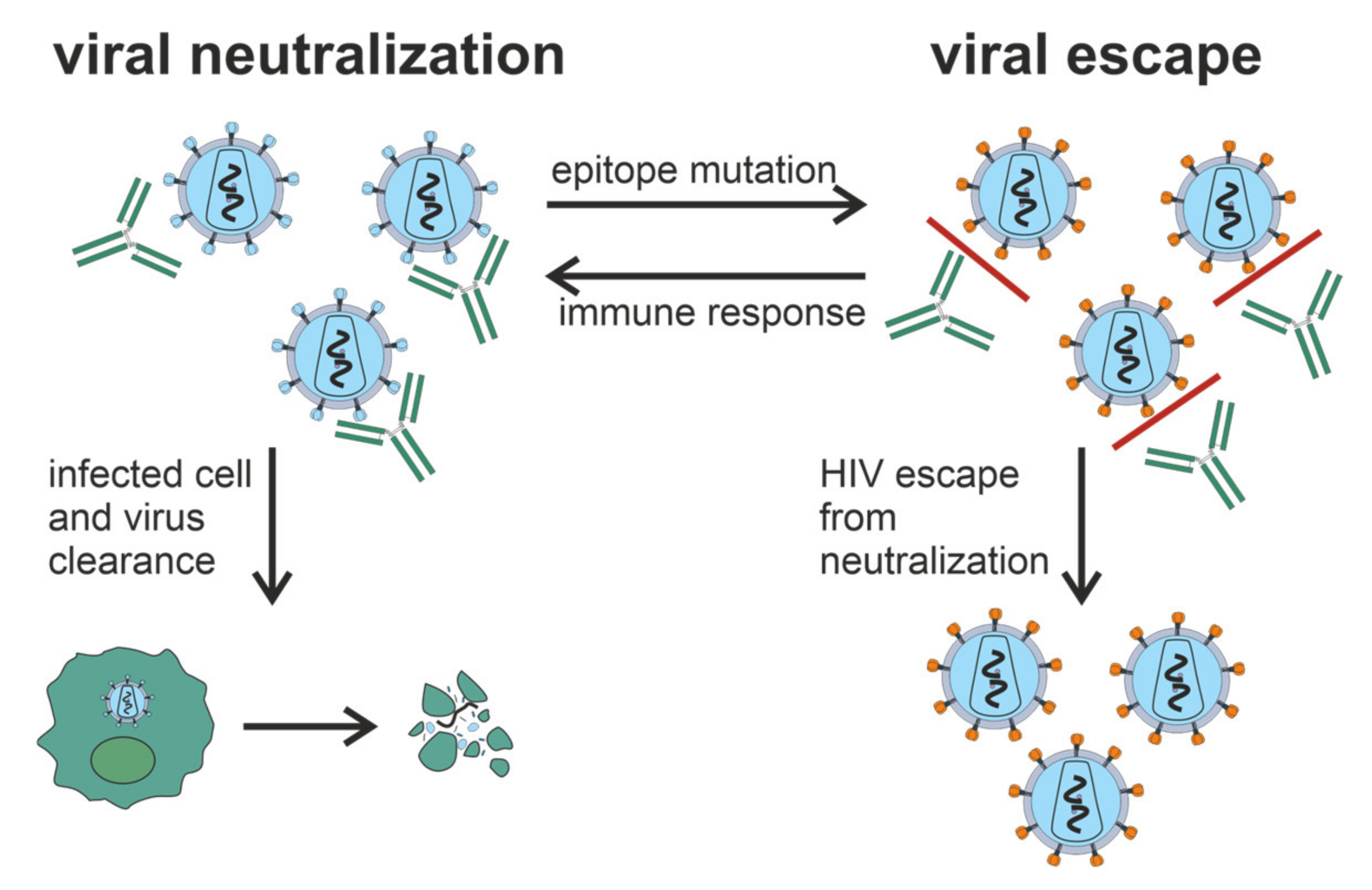
Figure 1. Generation of bNAbs, which neutralize the virus through binding to viral spikes and blocking the entry of the virus into sensitive cells such as CD4+ T cells, is related to the evasion of virus from the immune response due to the low precision of HIV reverse transcriptase and rapid generation of viral mutants. Glycan shield, immunodominant variable loops, and conformational masking of key viral epitopes protect Env from Ab-response.
Despite the enormous diversity of HIV, there are described a few bNAbs such as b12 [23][24], 2G12 [24][25], 4E10 [26], 2F5 [24][27], VRC01 [28], PG9/PG16, 447-52D [29]. As shown in [30][31][32], b12 Ab isolated from the phage display library can neutralize about 40% of known HIV strains. Another monoclonal antibody (mAb) HJ16, also neutralizes about 40% of viral isolates [33]. Most of the structures of such monoclonal bNAbs are resolved in complexes with Env [8][34]. These structures can be used to develop immunogens capable of eliciting bNAbs.
Although the detection of circulating autoantibodies (autoAbs) to self-antigens in patients infected by HIV or other viruses does not necessarily reflect the presence of an autoimmune disease, such autoAbs can certainly complicate viral infections. These responses may vary due to the infection of autologous cells by the virus and their consequent targeting by pre-existing and/or induced autoAbs against the virus. Several natural autoAbs were described in HIV-infection [35]: Abs against small nuclear ribonucleoproteins (snRNP) [36], Abs to anticardiolipin (aCL) and antiglycoprotein 1 (aβ2GP1), anti-DNA, and antinuclear Abs [37][38]. Common to some viral diseases and autoimmune pathologies, catalytic Abs (Abzymes) hydrolyzing DNA [39] and a variety of proteins were described in HIV/AIDS patients: histones [40], myelin basic protein (MBP) [41], HIV integrase [42][43] and HIV reverse transcriptase [39], β-casein and serum albumin [39]. Such autoAbs may be beneficial for patients, as they expel autologous cells expressing viral antigens from the body. Moreover, one cannot exclude that HIV-infected patients produce natural Abs that recognize both microbial and autologous antigens.
Several vaccines in preclinical tests consisted of single coat proteins or in combination with other HIV proteins and multiepitope synthetic peptides and polypeptides expressed by noninfectious vectors. Numerous HIV vaccine candidates are directed to the induction of nAbs and/or cytotoxic T cells [44]. Despite the induction of robust immune responses, the recombinant glycoprotein gp120 VaxGen [45] and Merck gag/pol/nef (STEP) adenoviral vaccine [46] did not reduce the risk of infection.
2. Neutralizing Antibodies in HIV-Infection
Neutralizing Abs protect cells from a viral particle by binding it. These Abs surround the virus, after which the entire complex containing the virus is removed with the immune system. The induction of nAbs is a key goal of vaccination strategies [47][48].
During HIV-infection, almost all patients produce Abs to Env, but only a small fraction of these Abs can neutralize the virus [49][50]. Interestingly, nAbs against viral Env are produced within the first weeks of infection. Still, this early Ab response targets an autologous virus that circulates within each person and is ineffective against heterologous (unrelated) viruses [5].
Among the nAbs, a specific group of Abs neutralizing a wide spectrum of HIV strains are distinguished. These broadly neutralizing antibodies (bNAbs) arise after several years of virus–antibody coevolution in infected patients [51][52][53]; bNAbs usually recognize conserved epitopes on the highly glycosylated envelope glycoprotein (Env) [54][55].
2.1. General Characteristics of nAbs
The first B cell response to HIV-infection appears within 8 days after detectable viremia [56]. After the next 5 days, circulating Abs against gp41 are detected, and after another 2 weeks, Abs against gp120 are detected, which primarily target the V3 loop. Autologous nAbs are developed over several months [57] and target variable HIV regions by potent but particular molecules [5][58][59]. Over the years, Abs with neutralizing cross-specific potential developed in one third of chronically infected patients and targeted the more conserved regions of Env [60]. Heterologous nAbs appear in some patients 1 year after infection reach their peak after 4 years, with no increase after that [19]. High viral load is not a general predictor of Ab neutralization capacity since some patients with high viremia do not develop cross-neutralizing Abs [19][61].
The breadth of Ab neutralization is usually increased to the end of 2.5 years after infection [62]. In the long-term nonprogressor patients, all neutralizing Abs against gp120 were clonally associated with the antibody-diversifying process of somatic hypermutation [63]. B cells producing bNAbs have been shown to undergo several rounds of affinity maturation in germinal centers to achieve cross-neutralizing activity [64][65]. BNAbs develop over time and are maintained by chronic antigen exposure.
2.2. Antibodies against Env Epitopes
Several papers are devoted to mapping the Ab-specificity responsible for cross-neutralizing activity [66][67][68]. These nAbs recognize epitopes of monomeric gp120; in some cases, cross-neutralizing activity can be attributed to Abs recognizing linear epitopes in the membrane-proximal outer region (MPER) of gp41 [69][70]. It has been shown that the quaternary epitope at the end of the trimeric envelope structure, including loops V2 and V3, is often the target of cross-neutralizing Abs [17][32][71].
Figure 2 shows vulnerable targets of HIV: nAbs react with the HIV Env spike, which is composed of three highly glycosylated gp120 molecules, each noncovalently linked to a transmembrane gp41 molecule. To initiate entry of the virus into cells, gp120 binds to the CD4 receptor on the cell surface [72]. One of the structurally conserved regions of Env among the various isolates is the CD4 binding site on gp120, which is required for binding with the CD4 receptor on the surface of immune cells, such as T helper cells and monocytes. The second structurally conserved region of Env is the coreceptor binding site of gp120. Both the CD4 binding site and the coreceptor binding site are immunogenic, and numerous mAbs against these Env regions have been isolated from HIV-infected patients. However, most human mAbs against these regions cannot bind the native viral Env trimer and, therefore, cannot neutralize HIV [60].
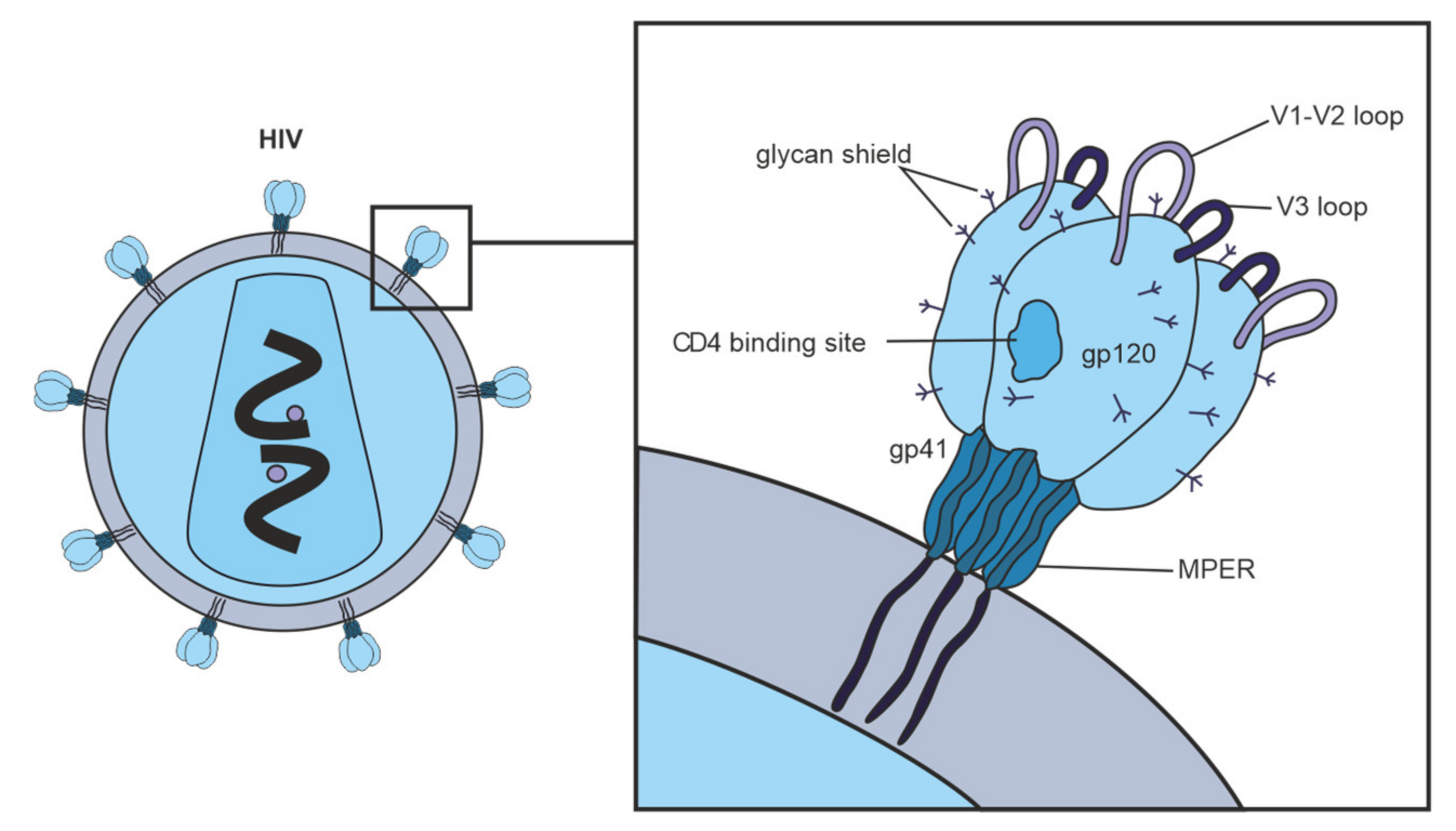
Figure 2. Vulnerable targets on trimeric glycoproteins of HIV spike (gp120 and gp41). Widely neutralizing Abs target the CD4 binding site on gp120, the proximal outer membrane region of gp41, the glycan shield, and epitopes in variable loops 1, 2, and 3 on gp120.
2.3. Hypervariable Domains gp120
HIV gp120 contains five “hypervariable” domains. The first two domains (V1V2) include dramatic insertions and deletions and different glycosylation patterns [73]. The V1V2 region regulates the neutralization sensitivity of conserved epitopes such as the coreceptor binding site [8][74][75]. Among viruses of the B subtype, a high ratio of nonsynonymous and synonymous substitutions is characteristic of the V3 region, while in the C subtype, this region remains relatively conserved [76]. Abs against V3 play a minimal role in neutralizing primary viruses [31][77] because the V3 loop is clogged with trimeric Env [75][78][79]. In contrast to V1V2, the role of V4 and V5 in neutralization resistance is unclear, although these regions affect Env conformation and glycan packaging, thereby sterically limiting the availability of neutralization determinants [5][80][81].
3. Autoantibodies in HIV-infection
Autoantibodies can interact with antigens specific to their organism. Pathogenesis of HIV/AIDS involves several factors, not a single virus-controlled destruction of CD4+ T cells [82]; among various pathogenic mechanisms there is autoimmunity [35] directed against lymphocytes [83][84][85], platelets [86][87] and peripheral nerves [88][89]. Up to 40% of HIV-infected patients are positive for autoAbs against red blood cells [90][91], which are involved in HIV-associated autoimmune hemolytic anemia [92][93].
Circulating Abs to small nuclear ribonucleoproteins are presented in the serum of HIV-infected patients [36]. Blood serum of HIV-infected patients contains Abs against anticardiolipin (aCL) and antiglycoprotein 1 (aβ2GP1), anti-DNA, and antinuclear Abs [37][38]. Exact frequency of autoimmune manifestations and their pathogenesis during HIV-infection remain unknown. Still, it was shown that anti-aCL Abs among HIV-infected patients is much higher than among patients with autoimmune diseases [94]. Content of these and similar Abs among HIV-infected patients before antiretroviral therapy ranges from 36% to 67% [37][95]. The presence of anti-aCL Abs in blood serum is not associated with any clinical manifestations of antiphospholipid syndrome, and it is not easy to explain why [96]. Moreover, spontaneous control of HIV replication or subsequent after antiretroviral therapy is associated with lower production of anti-aCL Abs [97][98].
4. Catalytic Antibodies in HIV/AIDS
Autoimmune reactions and the appearance of autoAbs in HIV/AIDS and some other viral infections may be associated with the activation of polyclonal B cells, molecular mimicry between viral and/or microbial antigens and host antigens [99][100], abnormal expression of immunoregulatory molecules, and anti-idiotypic network [38][101]. One can propose several physiological functions of the hydrolysis of DNA, histones, and MBP by Abs. It is suggested that the most relevant function of such catalytic Abs is the elimination of autoantigens formed due to the destruction of infected cells from the blood. However, it cannot be excluded the possibility that such autoAbs may also explain some of the observed autoimmune pathologies found in patients with HIV/AIDS.
Activation of B-lymphocytes in HIV-infected patients leads to the production of Abs to viral components and autoAbs to various components of human cells [38]. It has been shown that IgG and/or IgM of patients with AIDS hydrolyze not only autoantigens as DNA [102], MBP [41], histones [40][103][104][105], but also viral enzymes HIV integrase [42][43][106][107] and HIV reverse transcriptase [39] and corresponding peptides. Interestingly, IgG of HIV-infected patients efficiently cleaves five human histones (H1 [41][103], H2a, H2b [104], H3, and H4 [105]).
4.1. Catalytic Antibodies Cleave gp120
Hydrolysis of gp120 is possessed by V domains of the heavy (VH) and light (VL) chains of natural IgG, IgA, and IgM [108][109]. A rare light chain of Abs, isolated by phage display, can bind and hydrolyze gp120 independently of the heavy chain [110]. The most described Abs catalytic sites are located in the VL domains [111].
Long-term HIV-infection for 5 years is associated with a modest increase in catalytic IgA against gp120 [109]. No accumulation of such Abs was observed at the earlier stages of disease (6 months) [112].
4.2. Antibodies Hydrolyze HIV Integrase, Reverse Transcriptase, and Corresponding Oligopeptides
IgG and IgM were isolated from the blood serum of HIV-infected patients and subsequently separated on columns with immobilized reverse transcriptase or integrase, respectively, specifically hydrolyzed only recombinant viral reverse transcriptase [39] or integrase [42][43]. Abs that specifically hydrolyze these HIV enzymes were of potential interest in developing new anti-HIV-drugs for many years.
Nearly 40 sites of HIV integrase cleavage by Abs isolated from HIV-infected patients were found. Most sites are located in seven known immunodominant integrase sequences [106]. Two 20 mer oligopeptides corresponding to immunogenic integrase sequences contained 9 to 10 clusters of major, moderate, and minor cleavage sites [106][113]. Each individual Ab preparation from HIV-infected patients had a distinct ratio of hydrolysis sites [106].
4.3. Antibodies Hydrolyze DNA, Histones, and Myelin Basic Protein
Relative DNase activity of Abs isolated from the blood serum of HIV-infected donors varies significantly from patient to patient, but 96% of the preparations show a detectable level of DNase activity [102], which is an intrinsic feature of Abs [39]. Abs against H1, H3, H4, and MBP possess enzymatic cross-reactivity [114]: anti-histone H1 IgG hydrolyze H1 as well as MBP, and vice versa, anti-MBP Abs hydrolyze this H1 histone [41]. Similarly, IgG against H2a and H2b histones efficiently hydrolyze these histones and MBP, and anti-MBP Abs cleave MBP, H2a, and H2b histones, but not other control proteins [114]. Near 100% of IgG of HIV-infected patients effectively hydrolyze between one and five human histones [40].
5. HIV-vaccines: Production of Neutralizing Antibodies
Current HIV prevention and treatment strategies include using antiretroviral drugs for pre-exposure prophylaxis and antiretroviral therapy. This made it possible to transform HIV from a life-threatening disease to a manageable chronic disease [115]. However, drugs are expensive, require strict dosing to be effective, and cause side effects. Some HIV-infected patients develop drug resistance; furthermore, drug access remains a significant barrier, especially in low- and middle-income countries.
HIV replicates chronically in the host and evades an Ab response, unlike other viral pathogens. Immunity evasion and large genetic variation among HIV strains are significant obstacles to vaccine development. The use of vaccines should provide generation of Abs that can adapt to the Env glycan screen and bind to various viral strains to offer complete protection [116][117]. An ideal HIV-vaccine would elicit serum Abs that are highly effective and can prevent viral infection at low concentrations and have broad action—making a high percentage of hard-to-neutralize viruses inactive [118].
5.1. HIV-vaccine Development Strategies
The induction of a protective Ab response remains the main priority in developing HIV-vaccines [119]. Unfortunately, most current HIV-vaccine candidates do not elicit nAbs against most circulating viral strains. To replicate chronically in the host, the HIV uses several mechanisms to defend itself against Ab recognition: Env glycoprotein is protected by the shield of glycans. Different variations occur in immunodominant loops, and key viral epitopes are due to the conformational lability [5][21][49] (Figure 3). These defense mechanisms, although very effective, have vulnerabilities; information on the precise location and molecular structure of these vulnerable regions can be useful for the rational design of improved vaccine immunogens.
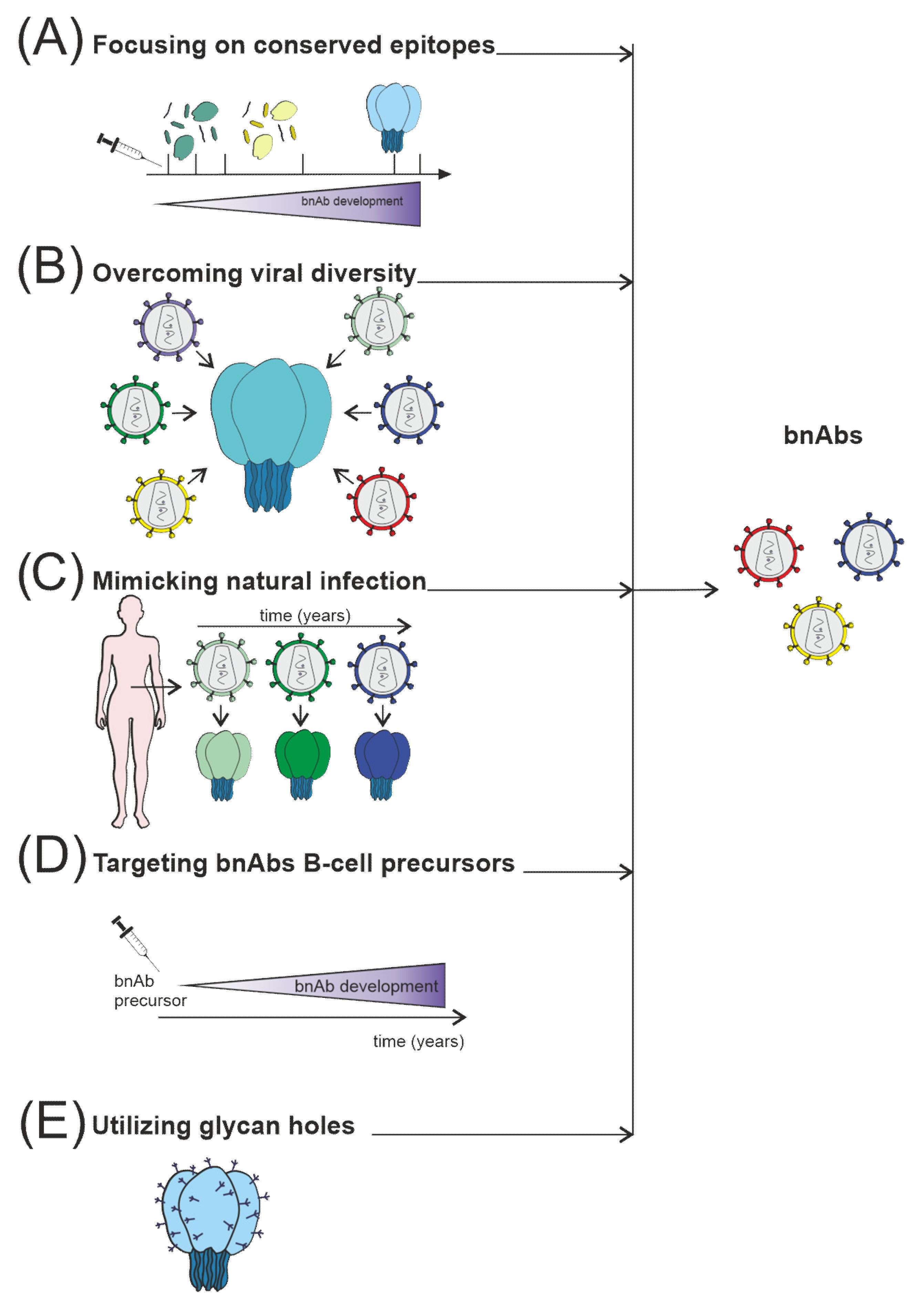
Figure 3. Several strategies are currently used to stimulate nAb responses towards bnAb generation, which consist of (A) directing of Ab response to conserved sites, for example, using fusion peptides; (B) use of mosaic or consensus antigens to overcome the viral diversity of circulating HIV worldwide; (C) clonal immunogens resembling the evolution of the virus in HIV-infected individuals modeling natural infection; (D) targeting immunogens to putative bnAb germline progenitors, followed by immunization leading to the affinity maturation pathway to bnAb developm€; (E) selective removing of glycans to focus Ab responses to a specific site of interest, such as CD4 binding site immunogens.
5.2. Immunogen Structure Design
HIV predominantly induces non-neutralizing or strain-specific Abs during the first months after infection [5][57]. It was found that approximately 10–20% of HIV-infected patients had bNAbs after a few years of disease [17][60][120], which is the humoral immune response that a vaccine should elicit.
Viral epitopes conserved among most viral strains are more likely to generate cross-reactive Abs. In this regard, research is focused on small numbers of human mAbs isolated from HIV-infected individuals that possess cross-reactive neutralizing activity [49][121]. However, these conserved viral epitopes are weakly immunogenic or produce Abs with limited reactivity. Minor structural changes may improve specific binding [20][122]. The crystal structure of b12 mAb bound to the CD4 receptor binding site of the gp120 molecule provides insight into how nAbs gain access to the functionally conserved regions of the Env glycoprotein [34].
5.3. Vaccination Inducing Immune Response at Mucous Membranes
Mucosal tissues of the gastrointestinal tract and vagina are the main reservoirs for the initial replication and reproduction of HIV, as well as sites of rapid depletion of CD4+ T cells [123]. Such viral reservoirs are considered significant difficulties in eradicating HIV in infected hosts. A successful vaccine should prevent the creation of these reservoirs at a very early stage of HIV-infection [124]. Protection of mucous membranes from HIV is mainly provided due to the Abs. Vaccination is not necessarily resulting in extremely high levels of nAb to protect the mucosal surface. Still, the Ab response must be robust, and nAb must cross-react with the genetically diverse spectra of HIV [125].
5.4. Covalent Epitope-Based Vaccines
Although the virus mutates rapidly, it must retain specific surface protein epitopes to maintain its infectivity. Inducing a robust immune response against structurally conserved epitopes essential to the viral life cycle is a logical route to a vaccine development that will be effective worldwide and minimize the possibility of viral mutants’ escape. The target site should be expressed on the surface of free virions in a form that is sterically accessible to Abs. The main problem is that the vulnerable epitopes of HIV are weakly immunogenic. Peptide immunogens with sequences identical to the linear conserved epitopes of gp120 can be easily synthesized, but the conformation of such peptides may also differ from that of the native epitope.
5.5. HIV mRNA Vaccines
The SARS-CoV-2 pandemic introduced the world to a new type of vaccine—mRNA encapsulated in lipid nanoparticles. mRNA-based vaccines rely on the production of individual immunogens by the host cells, which in turn are targets for Ab responses and cytotoxic T cells [126]. Traditional subunit vaccines deliver immunogens in a protein that degrades immediately after injection. In contrast, mRNA-based vaccines induce weekly production of immunogens in situ [127], increasing the possibility that rare bnAb progenitor B cells will become activated and proliferate after immunization. Prolonged production of mRNA-encoded immunogens in vivo works much like antigen release via an osmotic pump. Slow antigen delivery increases the activity of the germinal center and the induction of HIV nAbs [128].
6. Immune Exhaustion and Antibody Response
Immune exhaustion is the loss of effector functions and proliferative capacity of memory T cells [129]. The process of immune exhaustion in the context of persistent viral infections was first described in a mouse model of lymphocytic choriomeningitis (LCMV). It has been shown that LCMV-specific CD8+ T cells are preserved during chronic infection but are not cytotoxic [130]. Subsequently, T cell exhaustion has been described in patients with chronic viral infections such as HIV, hepatitis B, and hepatitis C [131][132][133].
Immune exhaustion was described for both CD8+ and CD4+ T cells [134][135]. Antigen-specific exhaustion of effector and memory cells includes a gradual loss of effector function and proliferative capacity, which progress until such cells are eliminated [136]. Exhausted CD8+ T cells significantly differ from terminally differentiated or memory CD8+ T cells: they secrete fewer cytokines and are characterized by cell surface expression of inhibitory receptors such as PD-1, CTLA4, Tim3, TIGIT, CD160, and LAG-3 [137][138][139]. Such surface expression of inhibitory receptors increases upon activation and limits the overactivation of T cells, providing the so-called immune checkpoints (IC) [134].
7. COVID-19 and HIV-infection
There is little evidence on the contribution of HIV to previous epidemics of respiratory viruses. HIV is associated with a higher risk of severe respiratory infections, including seasonal influenza [140][141]. However, the contribution of HIV-infection to outcomes during the 2009 H1N1 influenza pandemic was unclear. There would be no substantial evidence that HIV-infected individuals were at increased risk of infection or had worse outcomes unless they were in advanced immunosuppression [142]. HIV-infection was not associated with increased disease severity during previous SARS and MERS outbreaks; there were only a few reports of mild illness among people living with HIV [143][144].
The SARS-CoV-2 pandemic has become the greatest threat to global health in the modern era. It should be recognized that HIV/AIDS and COVID-19 are completely different diseases with different modes of transmission and disease course. First, HIV spreads through body fluids, while COVID-19 is currently considered an acute airborne infection [145]. People with chronic illnesses are at risk of relatively more severe COVID-19 symptoms, including those with weakened immune systems [146][147][148]. Therefore, HIV-infected patients may experience heightened feelings of anxiety about being infected with COVID-19 [145][149][150]. The US Centers for Disease Control and Prevention classifies immunocompromised people as high-risk, focusing on people living with uncontrolled HIV/AIDS.
7.1. Immunological Features of COVID-19 in HIV-infected Patients
If untreated, HIV-infection leads to a decrease in the CD4 T cell count, leading to AIDS. AIDS is defined as a CD4 cell count <200 cells/μL [151]. If HIV-infection is controlled, the risk of serious complications from COVID-19, and therefore poor outcomes, is likely to be low. However, the same cannot be said for poorly controlled HIV-infection or AIDS [152].
7.2. SARS-CoV-2 and HIV-vaccines
In the case of HIV, the virus integrates into the genome, forms a CD4+ T cell reservoir, leading to chronic infection [153]. The target of nAbs in the case of SARS-CoV-2 is the receptor-binding domain of the S-glycoprotein. Neutralizing Abs block the binding of SARS-CoV-2 virion to the Ace2-receptor [154][155][156]. Abs neutralizing SARS-CoV-2 are easily formed in the body after COVID-19 infection or after vaccination [157][158][159]; on the opposite, HIV can evade immunity and possess significant genetic variation among strains. This poses serious obstacles to vaccine development; the vaccine must produce Abs that can adapt to the Env glycan screen and bind to various viral strains to provide protection [116][117].
8. Conclusions
The highest priority in HIV research remains the development of a preventive vaccine. Despite many years of attempts to develop a HIV-vaccine based on classical strategies, it has not been possible so far. The search for a vaccine is still ongoing. One of the main focuses of a preventive vaccine is the induction of protective immune responses in the early stages of HIV-infection. Among the main obstacles encountered in developing an effective vaccine are mutational variability and global viral diversity, making it easy to avoid cellular and humoral host responses. One of the goals of studying Abs in HIV-infection is to determine how to safely induce broadly reactive protective Abs against HIV. At the same time, such Abs should trigger a cascade of autoimmune reactions. Since HIV is transmitted primarily through mucous membranes, understanding antiviral immunity in mucosal sites is of great importance.
The first step in developing effective vaccines is understanding HIV evolutionary strategies. One of these strategies is the ability of HIV to suppress the adaptive immune response to vulnerable envelope epitopes, which must be maintained in the most conservative form since they are necessary to maintain the infectivity of the virus. Because of HIV ability to elude Ab responses, the vaccine must induce the production of multiple bNAbs that will target various conserved sites on the Env glycoprotein. Thus, several batches of immunogens may be required.
Various Abs against antigens were described in HIV, including autoAbs with catalytic activities. Biological functions of natural Abs in HIV-infection, and in the first place, polyreactive and autoAbs, have not been sufficiently studied. Thus, the task of analyzing natural and artificial Abs against HIV in the context of developing an HIV-vaccine is unlikely to be completed soon. When creating a vaccine against HIV, it is necessary to consider the design of the immunogen and the method of its delivery, and the production of nAbs and avoid the production of harmful autoAbs. Among the priorities is the solution of the following problems: generation of bNAbs after vaccination and production of monoclonal bNAbs with broad prospects for therapeutic use.
References
- Barré-Sinoussi, F.; Ross, A.L.; Delfraissy, J.-F. Past, present and future: 30 years of HIV research. Nat. Rev. Microbiol. 2013, 11, 877–883.
- Taylor, B.S.; Sobieszczyk, M.E.; McCutchan, F.E.; Hammer, S.M. The Challenge of HIV-1 Subtype Diversity. N. Engl. J. Med. 2008, 358, 1590–1602.
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. AIDS 2006, 20, W13–W23.
- Korber, B.; Gaschen, B.; Yusim, K.; Thakallapally, R.; Kesmir, C.; Detours, V. Evolutionary and immunological implications of contemporary HIV-1 variation. Br. Med. Bull. 2001, 58, 19–42.
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody neutralization and escape by HIV-1. Nature 2003, 422, 307–312.
- Richman, D.D.; Wrin, T.; Little, S.J.; Petropoulos, C.J. Rapid evolution of the neutralizing antibody response to HIV type 1 infection. Proc. Natl. Acad. Sci. USA 2003, 100, 4144–4149.
- Liu, J.; Bartesaghi, A.; Borgnia, M.J.; Sapiro, G.; Subramaniam, S. Molecular architecture of native HIV-1 gp120 trimers. Nature 2008, 455, 109–113.
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 1998, 393, 648–659.
- Chen, B.; Vogan, E.M.; Gong, H.; Skehel, J.J.; Wiley, D.C.; Harrison, S.C. Structure of an unliganded simian immunodeficiency virus gp120 core. Nature 2005, 433, 834–841.
- Yu, W.-H.; Zhao, P.; Draghi, M.; Arevalo, C.; Karsten, C.B.; Suscovich, T.J.; Gunn, B.; Streeck, H.; Brass, A.L.; Tiemeyer, M.; et al. Exploiting glycan topography for computational design of Env glycoprotein antigenicity. PLoS Comput. Biol. 2018, 14, e1006093.
- Barin, F.; McLane, M.F.; Allan, J.S.; Lee, T.H.; Groopman, J.E.; Essex, M. Virus Envelope Protein of HTLV-III Represents Major Target Antigen for Antibodies in AIDS Patients. Science 1985, 228, 1094–1096.
- Allan, J.S.; Coligan, J.E.; Barin, F.; McLane, M.F.; Sodroski, J.G.; Rosen, C.A.; Haseltine, W.A.; Lee, T.H.; Essex, M. Major Glycoprotein Antigens That Induce Antibodies in AIDS Patients Are Encoded by HTLV-III. Science 1985, 228, 1091–1094.
- Okabe, M.; Kubara, K.; Kawaguchi, H.; Kawano, T.; Nakashima, Y.; Fukuda, K.; Hiroki, T.; Arakawa, K.; Kikuchi, M. A case of myxedema with diffuse myocardial fibrosis proven by endomyocardial biopsy. Kokyu Junkan Respir. Circ. 1990, 38, 1159–1163.
- Moore, J.P.; Cao, Y.; Ho, D.D.; Koup, R.A. Development of the anti-gp120 antibody response during seroconversion to human immunodeficiency virus type 1. J. Virol. 1994, 68, 5142–5155.
- Soudeyns, H.; Pantaleo, G. The moving target: Mechanisms of HIV persistence during primary infection. Immunol. Today 1999, 20, 446–450.
- Cao, Y.; Qin, L.; Zhang, L.; Safrit, J.; Ho, D.D. Virologic and Immunologic Characterization of Long-Term Survivors of Human Immunodeficiency Virus Type 1 Infection. N. Engl. J. Med. 1995, 332, 201–208.
- Walker, L.M.; Simek, M.D.; Priddy, F.; Gach, J.S.; Wagner, D.; Zwick, M.B.; Phogat, S.K.; Poignard, P.; Burton, D.R. A Limited Number of Antibody Specificities Mediate Broad and Potent Serum Neutralization in Selected HIV-1 Infected Individuals. PLoS Pathog. 2010, 6, e1001028.
- Doria-Rose, N.A.; Klein, R.M.; Daniels, M.G.; O’Dell, S.; Nason, M.; Lapedes, A.; Bhattacharya, T.; Migueles, S.A.; Wyatt, R.T.; Korber, B.T.; et al. Breadth of Human Immunodeficiency Virus-Specific Neutralizing Activity in Sera: Clustering Analysis and Association with Clinical Variables. J. Virol. 2010, 84, 1631–1636.
- Gray, E.S.; Madiga, M.C.; Hermanus, T.; Moore, P.L.; Wibmer, C.K.; Tumba, N.L.; Werner, L.; Mlisana, K.; Sibeko, S.; Williamson, C.; et al. The Neutralization Breadth of HIV-1 Develops Incrementally over Four Years and Is Associated with CD4+ T Cell Decline and High Viral Load during Acute Infection. J. Virol. 2011, 85, 4828–4840.
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Koff, W.C.; Kwong, P.D.; Moore, J.P.; Nabel, G.J.; Sodroski, J.; Wilson, I.A.; Wyatt, R.T. HIV vaccine design and the neutralizing antibody problem. Nat. Immunol. 2004, 5, 233–236.
- Kwong, P.D.; Doyle, M.L.; Casper, D.J.; Cicala, C.; Leavitt, S.A.; Majeed, S.; Steenbeke, T.D.; Venturi, M.; Chaiken, I.; Fung, M.; et al. HIV-1 evades antibody-mediated neutralization through conformational masking of receptor-binding sites. Nature 2002, 420, 678–682.
- Mascola, J.R.; Montefiori, D.C. HIV-1: Nature’s master of disguise. Nat. Med. 2003, 9, 393–394.
- Kessler, J.A.; McKenna, P.M.; Emini, E.A.; Chan, C.P.; Patel, M.D.; Gupta, S.K.; Mark, G.E.; Barbas, C.F.; Burton, D.R.; Conley, A.J. Recombinant Human Monoclonal Antibody IgG1b12 Neutralizes Diverse Human Immunodeficiency Virus Type 1 Primary Isolates. AIDS Res. Hum. Retroviruses 1997, 13, 575–582.
- Parren, P.W.H.I.; Wang, M.; Trkola, A.; Binley, J.M.; Purtscher, M.; Katinger, H.; Moore, J.P.; Burton, D.R. Antibody Neutralization-Resistant Primary Isolates of Human Immunodeficiency Virus Type 1. J. Virol. 1998, 72, 10270–10274.
- Trkola, A.; Purtscher, M.; Muster, T.; Ballaun, C.; Buchacher, A.; Sullivan, N.; Srinivasan, K.; Sodroski, J.; Moore, J.P.; Katinger, H. Human monoclonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J. Virol. 1996, 70, 1100–1108.
- Zwick, M.B.; Labrijn, A.F.; Wang, M.; Spenlehauer, C.; Saphire, E.O.; Binley, J.M.; Moore, J.P.; Stiegler, G.; Katinger, H.; Burton, D.R.; et al. Broadly Neutralizing Antibodies Targeted to the Membrane-Proximal External Region of Human Immunodeficiency Virus Type 1 Glycoprotein gp41. J. Virol. 2001, 75, 10892–10905.
- Muster, T.; Steindl, F.; Purtscher, M.; Trkola, A.; Klima, A.; Himmler, G.; Rüker, F.; Katinger, H. A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J. Virol. 1993, 67, 6642–6647.
- Wu, X.; Yang, Z.-Y.; Li, Y.; Hogerkorp, C.-M.; Schief, W.R.; Seaman, M.S.; Zhou, T.; Schmidt, S.D.; Wu, L.; Xu, L.; et al. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 2010, 329, 856–861.
- Conley, A.J.; Gorny, M.K.; Kessler, J.A.; Boots, L.J.; Ossorio-Castro, M.; Koenig, S.; Lineberger, D.W.; Emini, E.A.; Williams, C.; Zolla-Pazner, S. Neutralization of primary human immunodeficiency virus type 1 isolates by the broadly reactive anti-V3 monoclonal antibody, 447–452D. J. Virol. 1994, 68, 6994–7000.
- Burton, D.R.; Pyati, J.; Koduri, R.; Sharp, S.J.; Thornton, G.B.; Parren, P.W.H.I.; Sawyer, L.S.W.; Hendry, R.M.; Dunlop, N.; Nara, P.L.; et al. Efficient Neutralization of Primary Isolates of HIV-1 by a Recombinant Human Monoclonal Antibody. Science 1994, 266, 1024–1027.
- Binley, J.M.; Wrin, T.; Korber, B.; Zwick, M.B.; Wang, M.; Chappey, C.; Stiegler, G.; Kunert, R.; Zolla-Pazner, S.; Katinger, H.; et al. Comprehensive Cross-Clade Neutralization Analysis of a Panel of Anti-Human Immunodeficiency Virus Type 1 Monoclonal Antibodies. J. Virol. 2004, 78, 13232–13252.
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.-Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and Potent Neutralizing Antibodies from an African Donor Reveal a New HIV-1 Vaccine Target. Science 2009, 326, 285–289.
- Corti, D.; Langedijk, J.P.M.; Hinz, A.; Seaman, M.S.; Vanzetta, F.; Fernandez-Rodriguez, B.M.; Silacci, C.; Pinna, D.; Jarrossay, D.; Balla-Jhagjhoorsingh, S.; et al. Analysis of Memory B Cell Responses and Isolation of Novel Monoclonal Antibodies with Neutralizing Breadth from HIV-1-Infected Individuals. PLoS ONE 2010, 5, e8805.
- Zhou, T.; Xu, L.; Dey, B.; Hessell, A.J.; Van Ryk, D.; Xiang, S.-H.; Yang, X.; Zhang, M.-Y.; Zwick, M.B.; Arthos, J.; et al. Structural definition of a conserved neutralization epitope on HIV-1 gp120. Nature 2007, 445, 732–737.
- Morrow, W.J.W.; Isenberg, D.A.; Sobol, R.E.; Stricker, R.B.; Kieber-Emmons, T. AIDS virus infection and autoimmunity: A perspective of the clinical, immunological, and molecular origins of the autoallergic pathologies associated with HIV disease. Clin. Immunol. Immunopathol. 1991, 58, 163–180.
- González, C.M.; López-LONGO, F.J.; Samson, J.; Monteagudo, I.; Grau, R.; Rodríguez-Mahou, M.; ST-Cyr, C.; Lapointe, N.; Carreño, L. Antiribonucleoprotein Antibodies in Children with HIV Infection: A Comparative Study with Childhood-Onset Systemic Lupus Erythematosus. AIDS Patient Care STDS 1998, 12, 21–28.
- Argov, S.; Schattner, A.; Burstein, R.; Handzel, Z.T.; Shoenfeld, Y.; Bentwich, Z. Autoantibodies in male homosexuals and HIV infection. Immunol. Lett. 1991, 30, 31–35.
- Zandman-Goddard, G.; Shoenfeld, Y. HIV and autoimmunity. Autoimmun. Rev. 2002, 1, 329–337.
- Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A.; Odintsova, E.S.; Kharitonova, M.A.; Baranovskii, A.G.; Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A. Proteolytic activity of IgG antibodies from blood of acquired immunodeficiency syndrome patients. Biochemistry 2006, 71, 251–261.
- Baranova, S.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies from the sera of HIV-infected patients efficiently hydrolyze all human histones. J. Mol. Recognit. 2016, 29, 346–362.
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Nevinsky, G.A. Autoantibodies in HIV-infected patients: Cross site-specific hydrolysis of H1 histone and myelin basic protein. BioFactors 2019, 45, 211–222.
- Baranova, S.V.; Buneva, V.N.; Kharitonova, M.A.; Sizyakina, L.P.; Calmels, C.; Andreola, M.-L.; Parissi, V.; Nevinsky, G. HIV-1 integrase-hydrolyzing antibodies from sera of HIV-infected patients. Biochimie 2009, 91, 1081–1086.
- Baranova, S.V.; Buneva, V.N.; Kharitonova, M.A.; Sizyakina, L.P.; Calmels, C.; Andreola, M.-L.M.-L.; Parissi, V.; Zakharova, O.D.; Nevinsky, G.A. HIV-1 integrase-hydrolyzing IgM antibodies from sera of HIV-infected patients. Int. Immunol. 2010, 22, 671–680.
- Miedema, F. A brief history of HIV vaccine research: Stepping back to the drawing board? AIDS 2008, 22, 1699–1703.
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W. Bangkok Vaccine Evaluation Group Randomized, Double-Blind, Placebo-Controlled Efficacy Trial of a Bivalent Recombinant Glycoprotein 120 HIV-1 Vaccine among Injection Drug Users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671.
- Sekaly, R.-P. The failed HIV Merck vaccine study: A step back or a launching point for future vaccine development? J. Exp. Med. 2008, 205, 7–12.
- Fauci, A.S.; Marston, H.D. Toward an HIV vaccine: A scientific journey. Science 2015, 349, 386–387.
- Mascola, J.R.; Montefiori, D.C. The Role of Antibodies in HIV Vaccines. Annu. Rev. Immunol. 2010, 28, 413–444.
- Pantophlet, R.; Burton, D.R. GP120: Target for Neutralizing HIV-1 Antibodies. Annu. Rev. Immunol. 2006, 24, 739–769.
- Montefiori, D.C.; Morris, L.; Ferrari, G.; Mascola, J.R. Neutralizing and other antiviral antibodies in HIV-1 infection and vaccination. Curr. Opin. HIV AIDS 2007, 2, 169–176.
- Cheng-Mayer, C.; Homsy, J.; Evans, L.A.; Levy, J.A. Identification of human immunodeficiency virus subtypes with distinct patterns of sensitivity to serum neutralization. Proc. Natl. Acad. Sci. USA 1988, 85, 2815–2819.
- Brown, B.K.; Wieczorek, L.; Sanders-Buell, E.; Borges, A.R.; Robb, M.L.; Birx, D.L.; Michael, N.L.; McCutchan, F.E.; Polonis, V.R. Cross-clade neutralization patterns among HIV-1 strains from the six major clades of the pandemic evaluated and compared in two different models. Virology 2008, 375, 529–538.
- Kostrikis, L.G.; Cao, Y.; Ngai, H.; Moore, J.P.; Ho, D.D. Quantitative analysis of serum neutralization of human immunodeficiency virus type 1 from subtypes A, B, C, D, E, F, and I: Lack of direct correlation between neutralization serotypes and genetic subtypes and evidence for prevalent serum-dependent infecti. J. Virol. 1996, 70, 445–458.
- Haynes, B.F.; Burton, D.R.; Mascola, J.R. Multiple roles for HIV broadly neutralizing antibodies. Sci. Transl. Med. 2019, 11, eaaz2686.
- Burton, D.R.; Hangartner, L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol. 2016, 34, 635–659.
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-Cell Responses to Transmitted Human Immunodeficiency Virus Type 1: Virion-Binding Immunoglobulin M (IgM) and IgG Antibodies Followed by Plasma Anti-gp41 Antibodies with Ineffective Control of Initial Viremia. J. Virol. 2008, 82, 12449–12463.
- Gray, E.S.; Moore, P.L.; Choge, I.A.; Decker, J.M.; Bibollet-Ruche, F.; Li, H.; Leseka, N.; Treurnicht, F.; Mlisana, K.; Shaw, G.M.; et al. Neutralizing Antibody Responses in Acute Human Immunodeficiency Virus Type 1 Subtype C Infection. J. Virol. 2007, 81, 6187–6196.
- Li, B.; Decker, J.M.; Johnson, R.W.; Bibollet-Ruche, F.; Wei, X.; Mulenga, J.; Allen, S.; Hunter, E.; Hahn, B.H.; Shaw, G.M.; et al. Evidence for Potent Autologous Neutralizing Antibody Titers and Compact Envelopes in Early Infection with Subtype C Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 5211–5218.
- Moore, P.L.; Gray, E.S.; Choge, I.A.; Ranchobe, N.; Mlisana, K.; Abdool Karim, S.S.; Williamson, C.; Morris, L. The C3-V4 Region Is a Major Target of Autologous Neutralizing Antibodies in Human Immunodeficiency Virus Type 1 Subtype C Infection. J. Virol. 2008, 82, 1860–1869.
- Stamatatos, L.; Morris, L.; Burton, D.R.; Mascola, J.R. Neutralizing antibodies generated during natural HIV-1 infection: Good news for an HIV-1 vaccine? Nat. Med. 2009, 15, 866–870.
- Euler, Z.; van Gils, M.J.; Bunnik, E.M.; Phung, P.; Schweighardt, B.; Wrin, T.; Schuitemaker, H. Cross-Reactive Neutralizing Humoral Immunity Does Not Protect from HIV Type 1 Disease Progression. J. Infect. Dis. 2010, 201, 1045–1053.
- Mikell, I.; Sather, D.N.; Kalams, S.A.; Altfeld, M.; Alter, G.; Stamatatos, L. Characteristics of the Earliest Cross-Neutralizing Antibody Response to HIV-1. PLoS Pathog. 2011, 7, e1001251.
- Torán, J.L.; Kremer, L.; Sánchez-Pulido, L.; de Alborán, I.M.; del Real, G.; Llorente, M.; Valencia, A.; de Mon, M.A.; Martínez, A.C. Molecular analysis of HIV-1 gp120 antibody response using isotype IgM and IgG phage display libraries from a long-term non-progressor HIV-1-infected individual. Eur. J. Immunol. 1999, 29, 2666–2675.
- Pancera, M.; McLellan, J.S.; Wu, X.; Zhu, J.; Changela, A.; Schmidt, S.D.; Yang, Y.; Zhou, T.; Phogat, S.; Mascola, J.R.; et al. Crystal Structure of PG16 and Chimeric Dissection with Somatically Related PG9: Structure-Function Analysis of Two Quaternary-Specific Antibodies That Effectively Neutralize HIV-1. J. Virol. 2010, 84, 8098–8110.
- Zhou, T.; Georgiev, I.; Wu, X.; Yang, Z.-Y.; Dai, K.; Finzi, A.; Do Kwon, Y.; Scheid, J.F.; Shi, W.; Xu, L.; et al. Structural Basis for Broad and Potent Neutralization of HIV-1 by Antibody VRC01. Science 2010, 329, 811–817.
- Binley, J. Specificities of broadly neutralizing anti-HIV-1 sera. Curr. Opin. HIV AIDS 2009, 4, 364–372.
- Gray, E.S.; Taylor, N.; Wycuff, D.; Moore, P.L.; Tomaras, G.D.; Wibmer, C.K.; Puren, A.; DeCamp, A.; Gilbert, P.B.; Wood, B.; et al. Antibody Specificities Associated with Neutralization Breadth in Plasma from Human Immunodeficiency Virus Type 1 Subtype C-Infected Blood Donors. J. Virol. 2009, 83, 8925–8937.
- Li, Y.; Svehla, K.; Louder, M.K.; Wycuff, D.; Phogat, S.; Tang, M.; Migueles, S.A.; Wu, X.; Phogat, A.; Shaw, G.M.; et al. Analysis of Neutralization Specificities in Polyclonal Sera Derived from Human Immunodeficiency Virus Type 1-Infected Individuals. J. Virol. 2009, 83, 1045–1059.
- Gray, E.S.; Madiga, M.C.; Moore, P.L.; Mlisana, K.; Abdool Karim, S.S.; Binley, J.M.; Shaw, G.M.; Mascola, J.R.; Morris, L. Broad Neutralization of Human Immunodeficiency Virus Type 1 Mediated by Plasma Antibodies against the gp41 Membrane Proximal External Region. J. Virol. 2009, 83, 11265–11274.
- Sather, D.N.; Stamatatos, L. Epitope specificities of broadly neutralizing plasmas from HIV-1 infected subjects. Vaccine 2010, 28, B8–B12.
- Moore, P.L.; Gray, E.S.; Sheward, D.; Madiga, M.; Ranchobe, N.; Lai, Z.; Honnen, W.J.; Nonyane, M.; Tumba, N.; Hermanus, T.; et al. Potent and Broad Neutralization of HIV-1 Subtype C by Plasma Antibodies Targeting a Quaternary Epitope Including Residues in the V2 Loop. J. Virol. 2011, 85, 3128–3141.
- Wyatt, R.; Kwong, P.D.; Desjardins, E.; Sweet, R.W.; Robinson, J.; Hendrickson, W.A.; Sodroski, J.G. The antigenic structure of the HIV gp120 envelope glycoprotein. Nature 1998, 393, 705–711.
- Rong, R.; Bibollet-Ruche, F.; Mulenga, J.; Allen, S.; Blackwell, J.L.; Derdeyn, C.A. Role of V1V2 and Other Human Immunodeficiency Virus Type 1 Envelope Domains in Resistance to Autologous Neutralization during Clade C Infection. J. Virol. 2007, 81, 1350–1359.
- Krachmarov, C.; Pinter, A.; Honnen, W.J.; Gorny, M.K.; Nyambi, P.N.; Zolla-Pazner, S.; Kayman, S.C. Antibodies That Are Cross-Reactive for Human Immunodeficiency Virus Type 1 Clade A and Clade B V3 Domains Are Common in Patient Sera from Cameroon, but Their Neutralization Activity Is Usually Restricted by Epitope Masking. J. Virol. 2005, 79, 780–790.
- Krachmarov, C.P.; Honnen, W.J.; Kayman, S.C.; Gorny, M.K.; Zolla-Pazner, S.; Pinter, A. Factors Determining the Breadth and Potency of Neutralization by V3-Specific Human Monoclonal Antibodies Derived from Subjects Infected with Clade A or Clade B Strains of Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 7127–7135.
- Gaschen, B.; Taylor, J.; Yusim, K.; Foley, B.; Gao, F.; Lang, D.; Novitsky, V.; Haynes, B.; Hahn, B.H.; Bhattacharya, T.; et al. Diversity Considerations in HIV-1 Vaccine Selection. Science 2002, 296, 2354–2360.
- Li, M.; Gao, F.; Mascola, J.R.; Stamatatos, L.; Polonis, V.R.; Koutsoukos, M.; Voss, G.; Goepfert, P.; Gilbert, P.; Greene, K.M.; et al. Human Immunodeficiency Virus Type 1 env Clones from Acute and Early Subtype B Infections for Standardized Assessments of Vaccine-Elicited Neutralizing Antibodies. J. Virol. 2005, 79, 10108–10125.
- Hartley, O.; Klasse, P.J.; Sattentau, Q.J.; Moore, J.P. V3: HIV’s Switch-Hitter. AIDS Res. Hum. Retroviruses 2005, 21, 171–189.
- Pinter, A.; Honnen, W.J.; He, Y.; Gorny, M.K.; Zolla-Pazner, S.; Kayman, S.C. The V1/V2 Domain of gp120 Is a Global Regulator of the Sensitivity of Primary Human Immunodeficiency Virus Type 1 Isolates to Neutralization by Antibodies Commonly Induced upon Infection. J. Virol. 2004, 78, 5205–5215.
- Kinsey, N.E.; Anderson, M.G.; Unangst, T.J.; Joag, S.V.; Narayan, O.; Zink, M.C.; Clements, J.E. Antigenic Variation of SIV: Mutations in V4 Alter the Neutralization Profile. Virology 1996, 221, 14–21.
- Teeraputon, S.; Louisirirojchanakul, S.; Auewarakul, P. N-Linked Glycosylation in C2 Region of HIV-1 Envelope Reduces Sensitivity to Neutralizing Antibodies. Viral Immunol. 2005, 18, 343–353.
- Fauci, A.S. Multifactorial Nature of Human Immunodeficiency Virus Disease: Implications for Therapy. Science 1993, 262, 1011–1018.
- Tomar, R.H.; John, P.A.; Hennig, A.K.; Kloster, B. Cellular targets of antilymphocyte antibodies in AIDS and LAS. Clin. Immunol. Immunopathol. 1985, 37, 37–47.
- Tausk, F.A.; McCutchan, A.; Spechko, P.; Schreiber, R.D.; Gigli, I. Altered erythrocyte C3b receptor expression, immune complexes, and complement activation in homosexual men in varying risk groups for acquired immune deficiency syndrome. J. Clin. Investig. 1986, 78, 977–982.
- Ameglio, F.; Dolei, A.; Benedetto, A.; Sorrentino, R.; Tanigaki, N.; Tosi, R. Antibodies Reactive with Nonpolymorphic Epitopes on HLA Molecules Interfere in Screening Tests for the Human Immunodeficiency Virus. J. Infect. Dis. 1987, 156, 1034–1035.
- Morris, L.; Distenfeld, A.; Amorosi, E.; Karpatkin, S. Autoimmune Thrombocytopenic Purpura in Homosexual Men. Ann. Intern. Med. 1982, 96, 714–717.
- Stricker, R.B.; Abrams, D.I.; Corash, L.; Shuman, M.A. Target Platelet Antigen in Homosexual Men with Immune Thrombocytopenia. N. Engl. J. Med. 1985, 313, 1375–1380.
- Kiprov, D.; Pfaeffl, W.; Parry, G.; Lippert, R.; Lang, W.; Miller, R. Antibody-mediated peripheral neuropathies associated with ARC and AIDS: Successful treatment with plasmapheresis. J. Clin. Apher. 1988, 4, 3–7.
- de La Monte, S.M.; Gabuzda, D.H.; Ho, D.D.; Brown, R.H.; Hedley-Whyte, E.T.; Schooley, R.T.; Hirsch, M.S.; Bhan, A.K. Peripheral neuropathy in the acquired immunodeficiency syndrome. Ann. Neurol. 1988, 23, 485–492.
- De Angelis, V.; Biasinutto, C.; Pradella, P.; Vaccher, E.; Spina, M.; Tirelli, U. Clinical significance of positive direct antiglobulin test in patients with HIV infection. Infection 1994, 22, 92–95.
- Lai, M.; Visconti, E.; D’Onofrio, G.; Tamburrini, E.; Cauda, R.; Leone, G. Lower hemoglobin levels in human immunodeficiency virus-infected patients with a positive direct antiglobulin test (DAT): Relationship with DAT strength and clinical stages. Transfusion 2006, 46, 1237–1243.
- Yen, Y.-F.; Lan, Y.-C.; Huang, C.-T.; Jen, I.-A.; Chen, M.; Lee, C.-Y.; Chuang, P.-H.; Lee, Y.; Morisky, D.E.; Chen, Y.-M.A. Human Immunodeficiency Virus Infection Increases the Risk of Incident Autoimmune Hemolytic Anemia: A Population-Based Cohort Study in Taiwan. J. Infect. Dis. 2017, 216, 1000–1007.
- Yen, Y.-F.; Chuang, P.-H.; Jen, I.-A.; Chen, M.; Lan, Y.-C.; Liu, Y.-L.; Lee, Y.; Chen, Y.-H.; Chen, Y.-M.A. Incidence of autoimmune diseases in a nationwide HIV/AIDS patient cohort in Taiwan, 2000–2012. Ann. Rheum. Dis. 2017, 76, 661–665.
- Schved, J.F.; Dupuy-Fons, C.; Biron, C.; Quéré, I.; Janbon, C. A Prospective Epidemiological Study on the Occurrence of Antiphospholipid Antibody: The Montpellier Antiphospholipid (MAP) Study. Pathophysiol. Haemost. Thromb. 1994, 24, 175–182.
- Silvestris, F.; Frassanito, M.; Cafforio, P.; Potenza, D.; Di Loreto, M.; Tucci, M.; Grizzuti, M.; Nico, B.; Dammacco, F. Antiphosphatidylserine antibodies in human immunodeficiency virus-1 patients with evidence of T-cell apoptosis and mediate antibody- dependent cellular cytotoxicity . Blood 1996, 87, 5185–5195.
- Girón-González, J.A.; García del Río, E.; Rodríguez, C.; Rodríguez-Martorell, J.; Serrano, A. Antiphospholipid syndrome and asymptomatic carriers of antiphospholipid antibody: Prospective analysis of 404 individuals. J. Rheumatol. 2004, 31, 1560–1567.
- Calabrese, L.H.; Kirchner, E.; Shrestha, R. Rheumatic Complications of Human Immunodeficiency Virus Infection in the Era of Highly Active Antiretroviral Therapy: Emergence of a New Syndrome of Immune Reconstitution and Changing Patterns of Disease. Semin. Arthritis Rheum. 2005, 35, 166–174.
- Reveille, J.D. The changing spectrum of rheumatic disease in human immunodeficiency virus infection. Semin. Arthritis Rheum. 2000, 30, 147–166.
- Ternynck, T.; Falanga, P.B.; Unterklrscher, C.; Gregolre, J.; da Silva, L.P.; Avrameas, S. Induction of high levels of IgG autoantibodies in mice infected with Plasmodium chabaudi. Int. Immunol. 1991, 3, 29–37.
- Hentati, B.; Sato, M.N.; Payelle-Brogard, B.; Avrameas, S.; Ternynck, T. Beneficial effect of polyclonal immunoglobulins from malaria-infected BALB/c mice on the lupus-like syndrome of (NZB × NZW)F1 mice. Eur. J. Immunol. 1994, 24, 8–15.
- Barzilai, O.; Ram, M.; Shoenfeld, Y. Viral infection can induce the production of autoantibodies. Curr. Opin. Rheumatol. 2007, 19, 636–643.
- Odintsova, E.S.; Kharitonova, M.A.; Baranovskii, A.G.; Sizyakina, L.P.; Buneva, V.N.; Nevinsky, G.A. DNA-hydrolyzing IgG antibodies from the blood of patients with acquired immune deficiency syndrome. Mol. Biol. 2006, 40, 770–777.
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H1 histone from the sera of HIV-infected patients recognize and catalyze site-specific degradation of this histone. J. Mol. Recognit. 2017, 30, e2588.
- Baranova, S.V.; Dmitrienok, P.S.; Ivanisenko, N.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies to H2a and H2b histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. Mol. Biosyst. 2017, 13, 1090–1101.
- Baranova, S.V.; Dmitrenok, P.S.; Zubkova, A.D.; Ivanisenko, N.V.; Odintsova, E.S.; Buneva, V.N.; Nevinsky, G.A. Antibodies against H3 and H4 histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones. J. Mol. Recognit. 2018, 31, e2703.
- Odintsova, E.S.; Baranova, S.V.; Dmitrenok, P.S.; Rasskazov, V.A.; Calmels, C.; Parissi, V.; Andreola, M.-L.M.-L.; Buneva, V.N.; Zakharova, O.D.; Nevinsky, G.A. Antibodies to HIV integrase catalyze site-specific degradation of their antigen. Int. Immunol. 2011, 23, 601–612.
- Odintsova, E.S.; Baranova, S.V.; Dmitrenok, P.S.; Calmels, C.; Parissi, V.; Andreola, M.-L.; Buneva, V.N.; Nevinsky, G.A. Anti-integrase abzymes from the sera of HIV-infected patients specifically hydrolyze integrase but nonspecifically cleave short oligopeptides. J. Mol. Recognit. 2012, 25, 193–207.
- Townsley-Fuchs, J.; Kam, L.; Fairhurst, R.; Gange, S.J.; Goodglick, L.; Giorgi, J.V.; Sidell, N.; Detels, R.; Braun, J. Human immunodeficiency virus-1 (HIV-1) gp120 superantigen-binding serum antibodies. A host factor in homosexual HIV-1 transmission. J. Clin. Investig. 1996, 98, 1794–1801.
- Planque, S.; Mitsuda, Y.; Taguchi, H.; Salas, M.; Morris, M.-K.; Nishiyama, Y.; Kyle, R.; Okhuysen, P.; Escobar, M.; Hunter, R.; et al. Characterization of gp120 Hydrolysis by IgA Antibodies from Humans without HIV Infection. AIDS Res. Hum. Retroviruses 2007, 23, 1541–1554.
- Paul, S.; Karle, S.; Planque, S.; Taguchi, H.; Salas, M.; Nishiyama, Y.; Handy, B.; Hunter, R.; Edmundson, A.; Hanson, C. Naturally occurring proteolytic antibodies: Selective immunoglobulin M-catalyzed hydrolysis of HIV gp120. J. Biol. Chem. 2004, 279, 39611–39619.
- Gao, Q.-S.; Sun, M.; Rees, A.R.; Paul, S. Site-directed Mutagenesis of Proteolytic Antibody Light Chain. J. Mol. Biol. 1995, 253, 658–664.
- Chorny, A.; Puga, I.; Cerutti, A. Innate Signalling Networks in Mucosal IgA Class Switching. Adv Immunol 2010, 107, 31–69.
- Odintsova, E.S.; Dmitrenok, P.S.; Buneva, V.N.; Nevinsky, G.A. Specific anti-integrase abzymes from HIV-infected patients: A comparison of the cleavage sites of intact globular HIV integrase and two 20-mer oligopeptides corresponding to its antigenic determinants. J. Mol. Recognit. 2013, 26, 121–135.
- Baranova, S.V.; Dmitrenok, P.S.; Buneva, V.N.; Sedykh, S.E.; Nevinsky, G.A. HIV-Infected Patients: Cross Site-Specific Hydrolysis of H3 and H4 Histones and Myelin Basic Protein with Antibodies against These Three Proteins. Molecules 2021, 26, 316.
- Gulick, R.M.; Flexner, C. Long-Acting HIV Drugs for Treatment and Prevention. Annu. Rev. Med. 2019, 70, 137–150.
- Seaman, M.S.; Janes, H.; Hawkins, N.; Grandpre, L.E.; Devoy, C.; Giri, A.; Coffey, R.T.; Harris, L.; Wood, B.; Daniels, M.G.; et al. Tiered Categorization of a Diverse Panel of HIV-1 Env Pseudoviruses for Assessment of Neutralizing Antibodies. J. Virol. 2010, 84, 1439–1452.
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 2014, 409, 131–146.
- Montefiori, D.C.; Roederer, M.; Morris, L.; Seaman, M.S. Neutralization tiers of HIV-1. Curr. Opin. HIV AIDS 2018, 13, 128–136.
- Mascola, J.R.; D’Souza, P.; Gilbert, P.; Hahn, B.H.; Haigwood, N.L.; Morris, L.; Petropoulos, C.J.; Polonis, V.R.; Sarzotti, M.; Montefiori, D.C. Recommendations for the Design and Use of Standard Virus Panels To Assess Neutralizing Antibody Responses Elicited by Candidate Human Immunodeficiency Virus Type 1 Vaccines. J. Virol. 2005, 79, 10103–10107.
- Kwong, P.D.; Mascola, J.R. Human Antibodies that Neutralize HIV-1: Identification, Structures, and B Cell Ontogenies. Immunity 2012, 37, 412–425.
- Haynes, B.F.; Montefiori, D.C. Aiming to induce broadly reactive neutralizing antibody responses with HIV-1 vaccine candidates. Expert Rev. Vaccines 2006, 5, 579–595.
- Montefiori, D.; Sattentau, Q.; Flores, J.; Esparza, J.; Mascola, J. Antibody-Based HIV-1 Vaccines: Recent Developments and Future Directions. PLoS Med. 2007, 4, e348.
- Mehandru, S.; Poles, M.A.; Tenner-Racz, K.; Horowitz, A.; Hurley, A.; Hogan, C.; Boden, D.; Racz, P.; Markowitz, M. Primary HIV-1 Infection Is Associated with Preferential Depletion of CD4+ T Lymphocytes from Effector Sites in the Gastrointestinal Tract. J. Exp. Med. 2004, 200, 761–770.
- Katlama, C.; Deeks, S.G.; Autran, B.; Martinez-Picado, J.; van Lunzen, J.; Rouzioux, C.; Miller, M.; Vella, S.; Schmitz, J.E.; Ahlers, J.; et al. Barriers to a cure for HIV: New ways to target and eradicate HIV-1 reservoirs. Lancet 2013, 381, 2109–2117.
- Pavot, V.; Rochereau, N.; Lawrence, P.; Girard, M.P.; Genin, C.; Verrier, B.; Paul, S. Recent progress in HIV vaccines inducing mucosal immune responses. AIDS 2014, 28, 1701–1718.
- Mu, Z.; Haynes, B.F.; Cain, D.W. HIV mRNA Vaccines—Progress and Future Paths. Vaccines 2021, 9, 134.
- Pardi, N.; Hogan, M.J.; Naradikian, M.S.; Parkhouse, K.; Cain, D.W.; Jones, L.; Moody, M.A.; Verkerke, H.P.; Myles, A.; Willis, E.; et al. Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses. J. Exp. Med. 2018, 215, 1571–1588.
- Cirelli, K.M.; Carnathan, D.G.; Nogal, B.; Martin, J.T.; Rodriguez, O.L.; Upadhyay, A.A.; Enemuo, C.A.; Gebru, E.H.; Choe, Y.; Viviano, F.; et al. Slow Delivery Immunization Enhances HIV Neutralizing Antibody and Germinal Center Responses via Modulation of Immunodominance. Cell 2020, 180, 206.
- Khaitan, A.; Unutmaz, D. Revisiting Immune Exhaustion During HIV Infection. Curr. HIV/AIDS Rep. 2011, 8, 4–11.
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.D.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral Immune Evasion Due to Persistence of Activated T Cells Without Effector Function. J. Exp. Med. 1998, 188, 2205–2213.
- Urbani, S.; Amadei, B.; Tola, D.; Massari, M.; Schivazappa, S.; Missale, G.; Ferrari, C. PD-1 Expression in Acute Hepatitis C Virus (HCV) Infection Is Associated with HCV-Specific CD8 Exhaustion. J. Virol. 2006, 80, 11398–11403.
- Radziewicz, H.; Ibegbu, C.C.; Fernandez, M.L.; Workowski, K.A.; Obideen, K.; Wehbi, M.; Hanson, H.L.; Steinberg, J.P.; Masopust, D.; Wherry, E.J.; et al. Liver-Infiltrating Lymphocytes in Chronic Human Hepatitis C Virus Infection Display an Exhausted Phenotype with High Levels of PD-1 and Low Levels of CD127 Expression. J. Virol. 2007, 81, 2545–2553.
- Boni, C.; Fisicaro, P.; Valdatta, C.; Amadei, B.; Di Vincenzo, P.; Giuberti, T.; Laccabue, D.; Zerbini, A.; Cavalli, A.; Missale, G.; et al. Characterization of Hepatitis B Virus (HBV)-Specific T-Cell Dysfunction in Chronic HBV Infection. J. Virol. 2007, 81, 4215–4225.
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499.
- Hashimoto, M.; Kamphorst, A.O.; Im, S.J.; Kissick, H.T.; Pillai, R.N.; Ramalingam, S.S.; Araki, K.; Ahmed, R. CD8 T Cell Exhaustion in Chronic Infection and Cancer: Opportunities for Interventions. Annu. Rev. Med. 2018, 69, 301–318.
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T cell exhaustion during persistent viral infections. Virology 2015, 479–480, 180–193.
- Trautmann, L.; Janbazian, L.; Chomont, N.; Said, E.A.; Gimmig, S.; Bessette, B.; Boulassel, M.-R.; Delwart, E.; Sepulveda, H.; Balderas, R.S.; et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat. Med. 2006, 12, 1198–1202.
- Petrovas, C.; Casazza, J.P.; Brenchley, J.M.; Price, D.A.; Gostick, E.; Adams, W.C.; Precopio, M.L.; Schacker, T.; Roederer, M.; Douek, D.C.; et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J. Exp. Med. 2006, 203, 2281–2292.
- Wherry, E.J.; Ha, S.-J.; Kaech, S.M.; Haining, W.N.; Sarkar, S.; Kalia, V.; Subramaniam, S.; Blattman, J.N.; Barber, D.L.; Ahmed, R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity 2007, 27, 670–684.
- Kenmoe, S.; Bigna, J.J.; Fatawou Modiyingi, A.; Ndangang, M.S.; Ngoupo, P.A.; Simo, F.B.N.; Tchatchouang, S.; Temfack, E.; Njouom, R. Case fatality rate and viral aetiologies of acute respiratory tract infections in HIV positive and negative people in Africa: The VARIAFRICA-HIV systematic review and meta-analysis. J. Clin. Virol. 2019, 117, 96–102.
- Neuzil, K.M.; Reed, G.W.; Mitchel, E.F., Jr.; Griffin, M.R. Influenza-Associated Morbidity and Mortality in Young and Middle-Aged Women. JAMA 1999, 281, 901–907.
- Sheth, A.N.; Patel, P.; Peters, P.J. Influenza and HIV: Lessons from the 2009 H1N1 Influenza Pandemic. Curr. HIV/AIDS Rep. 2011, 8, 181–191.
- Al-Omari, A.; Rabaan, A.A.; Salih, S.; Al-Tawfiq, J.A.; Memish, Z.A. MERS coronavirus outbreak: Implications for emerging viral infections. Diagn. Microbiol. Infect. Dis. 2019, 93, 265–285.
- Moni, M.A.; Liò, P. Network-based analysis of comorbidities risk during an infection: SARS and HIV case studies. BMC Bioinform. 2014, 15, 333.
- Edelman, E.J.; Aoun-Barakat, L.; Villanueva, M.; Friedland, G. Confronting Another Pandemic: Lessons from HIV can Inform Our COVID-19 Response. AIDS Behav. 2020, 24, 1977–1979.
- Jiang, H.; Zhou, Y.; Tang, W. Maintaining HIV care during the COVID-19 pandemic. Lancet HIV 2020, 7, e308–e309.
- Blanco, J.L.; Ambrosioni, J.; Garcia, F.; Martínez, E.; Soriano, A.; Mallolas, J.; Miro, J.M. COVID-19 in patients with HIV: Clinical case series. Lancet HIV 2020, 7, e314–e316.
- Adepoju, P. Tuberculosis and HIV responses threatened by COVID-19. Lancet HIV 2020, 7, e319–e320.
- Algarin, A.B.; Varas-Rodríguez, E.; Valdivia, C.; Fennie, K.P.; Larkey, L.; Hu, N.; Ibañez, G.E. Symptoms, Stress, and HIV-Related Care Among Older People Living with HIV During the COVID-19 Pandemic, Miami, Florida. AIDS Behav. 2020, 24, 2236–2238.
- Harkness, A.; Behar-Zusman, V.; Safren, S.A. Understanding the Impact of COVID-19 on Latino Sexual Minority Men in a US HIV Hot Spot. AIDS Behav. 2020, 24, 2017–2023.
- Jordan, R.E.; Adab, P.; Cheng, K.K. COVID-19: Risk factors for severe disease and death. BMJ 2020, 368, m1198.
- Cooper, T.; Woodward, B.; Alom, S.; Harky, A. Coronavirus disease 2019 (COVID-19) outcomes in HIV/AIDS patients: A systematic review. HIV Med. 2020, 21, 567–577.
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23.
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569.
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263.
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6.
- Guo, L.; Ren, L.; Yang, S.; Xiao, M.; Chang, D.; Yang, F.; Dela Cruz, C.S.; Wang, Y.; Wu, C.; Xiao, Y.; et al. Profiling Early Humoral Response to Diagnose Novel Coronavirus Disease (COVID-19). Clin. Infect. Dis. 2020, 71, 778–785.
- Long, Q.-X.; Liu, B.-Z.; Deng, H.-J.; Wu, G.-C.; Deng, K.; Chen, Y.-K.; Liao, P.; Qiu, J.-F.; Lin, Y.; Cai, X.-F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848.
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Yuan, M.; Leung, W.S.; Chan, J.M.; Chik, T.S.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), March 2020. Eurosurveillance 2020, 25, 2000421.
More
Information
Subjects:
Immunology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.8K
Revisions:
2 times
(View History)
Update Date:
22 Mar 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No