 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Nikoleta Bizymi | + 2248 word(s) | 2248 | 2022-02-08 04:51:51 | | | |
| 2 | Lindsay Dong | Meta information modification | 2248 | 2022-03-11 06:38:41 | | | | |
| 3 | Lindsay Dong | Meta information modification | 2248 | 2022-03-18 02:19:45 | | |
Video Upload Options
Myeloid-derived suppressor cells (MDSCs) represent a heterogeneous population of myeloid cells that suppress immune responses in cancer, infection, and trauma. They mainly act by inhibiting T-cells, natural-killer cells, and dendritic cells, and also by inducing T-regulatory cells, and modulating macrophages. Although they are mostly associated with adverse prognosis of the underlying disease entity, they may display positive effects in specific situations, such as in allogeneic hematopoietic stem cell transplantation (HSCT), where they attenuate graft-versus-host disease (GVHD). They also contribute to the feto-maternal tolerance, and in the fetus growth process, whereas several pregnancy complications have been associated with their defects. Human umbilical cord blood (UCB) is a source rich in MDSCs.
1. Myeloid-Derived Suppressor Cells
2. Immune Cells and MDSCs in Pregnancy, Fetal-Maternal Cross-Talk, and Neonatal Period
3. Immune Cells and MDSCs in the Umbilical Cord Blood (UCB)
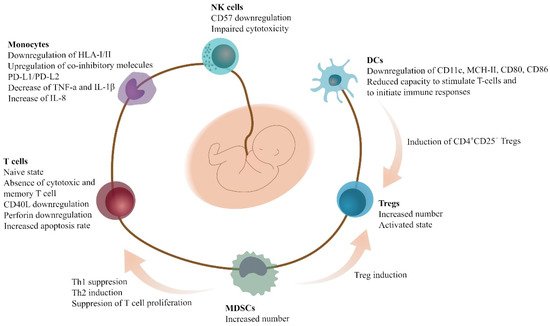
4. Conclusions
References
- Bronte, V.; Brandau, S.; Chen, S.-H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 2016, 7, 12150.
- Young, M.R.; Newby, M.; Wepsic, H.T. Hematopoiesis and suppressor bone marrow cells in mice bearing large metastatic Lewis lung carcinoma tumors. Cancer Res. 1987, 47, 100–105.
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174.
- Arocena, A.R.; Onofrio, L.I.; Pellegrini, A.V.; Silva, E.A.C.; Paroli, A.; Cano, R.C.; Aoki, M.P.; Gea, S. Myeloid-derived suppressor cells are key players in the resolution of inflammation during a model of acute infection. Eur. J. Immunol. 2013, 44, 184–194.
- Gabrilovich, D.I. MDSCs. Cancer Immunol. Res. 2018, 5, 3–8.
- Seman, B.G.; Robinson, C.M. The Enigma of Low-Density Granulocytes in Humans: Complexities in the Characterization and Function of LDGs during Disease. Pathogens 2021, 10, 1091.
- Vanhaver, C.; van der Bruggen, P.; Bruger, A. MDSC in Mice and Men: Mechanisms of Immunosuppression in Cancer. J. Clin. Med. 2021, 10, 2872.
- Millrud, C.R.; Bergenfelz, C.; Leandersson, K. On the origin of myeloid-derived suppressor cells. Oncotarget 2016, 8, 3649–3665.
- Bunt, S.K.; Yang, L.; Sinha, P.; Clements, V.K.; Leips, J.; Ostrand-Rosenberg, S. Reduced Inflammation in the Tumor Microenvironment Delays the Accumulation of Myeloid-Derived Suppressor Cells and Limits Tumor Progression. Cancer Res. 2007, 67, 10019–10026.
- Gabitass, R.F.; Annels, N.E.; Stocken, D.D.; Pandha, H.A.; Middleton, G.W. Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13. Cancer Immunol. Immunother. 2011, 60, 1419–1430.
- Ahmadi, M.; Mohammadi, M.; Ali-Hassanzadeh, M.; Zare, M.; Gharesi-Fard, B. MDSCs in pregnancy: Critical players for a balanced immune system at the feto-maternal interface. Cell. Immunol. 2019, 346, 103990.
- Umansky, V.; Blattner, C.; Gebhardt, C.; Utikal, J. The Role of Myeloid-Derived Suppressor Cells (MDSC) in Cancer Progression. Vaccines 2016, 4, 36.
- Mauti, L.A.; Le Bitoux, M.-A.; Baumer, K.; Stehle, J.-C.; Golshayan, D.; Provero, P.; Stamenkovic, I. Myeloid-derived suppressor cells are implicated in regulating permissiveness for tumor metastasis during mouse gestation. J. Clin. Investig. 2011, 121, 2794–2807.
- Poschke, I.; Mao, Y.; Adamson, L.; Salazar-Onfray, F.; Masucci, G.; Kiessling, R. Myeloid-derived suppressor cells impair the quality of dendritic cell vaccines. Cancer Immunol. Immunother. 2011, 61, 827–838.
- Fleming, V.; Hu, X.; Weber, R.; Nagibin, V.; Groth, C.; Altevogt, P.; Utikal, J.; Umansky, V. Targeting Myeloid-Derived Suppressor Cells to Bypass Tumor-Induced Immunosuppression. Front. Immunol. 2018, 9, 398.
- Tebartz, C.; Horst, S.A.; Sparwasser, T.; Huehn, J.; Beineke, A.; Peters, G.; Medina, E. A Major Role for Myeloid-Derived Suppressor Cells and a Minor Role for Regulatory T Cells in Immunosuppression during Staphylococcus aureus Infection Christina. J. Immunol. 2015, 194, 1100–1111.
- Tsukamoto, H.; Nishikata, R.; Senju, S.; Nishimura, Y. Myeloid-Derived Suppressor Cells Attenuate T H 1 Development through IL-6 Production to Promote Tumor Progression. Cancer Immunol. Res. 2013, 1, 64–76.
- Condamine, T.; Gabrilovich, D.I. Molecular mechanisms regulating myeloid-derived suppressor cell differentiation and function. Trends Immunol. 2011, 32, 19–25.
- De Sanctis, F.; Solito, S.; Ugel, S.; Molon, B.; Bronte, V.; Marigo, I. MDSCs in cancer: Conceiving new prognostic and therapeutic targets. Biochim. Biophys. Acta 2016, 1865, 35–48.
- Solito, S.; Pinton, L.; Mandruzzato, S. In Brief: Myeloid-derived suppressor cells in cancer. J. Pathol. 2017, 242, 7–9.
- Pastuła, A.; Marcinkiewicz, J. Myeloid-derived suppressor cells: A double-edged sword? Int. J. Exp. Pathol. 2011, 92, 73–78.
- Budhwar, S.; Verma, P.; Verma, R.; Rai, S.; Singh, K. The Yin and Yang of Myeloid Derived Suppressor Cells. Front. Immunol. 2018, 9, 2776.
- Pawelec, G.; Verschoor, C.P.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells: Not Only in Tumor Immunity. Front. Immunol. 2019, 10, 1099.
- Zhang, C.; Wang, S.; Yang, C.; Rong, R. The Crosstalk between Myeloid Derived Suppressor Cells and Immune Cells: To Establish Immune Tolerance in Transplantation. J. Immunol. Res. 2016, 2016, 4986797.
- Ostrand-Rosenberg, S. Myeloid derived-suppressor cells: Their role in cancer and obesity. Curr. Opin. Immunol. 2018, 51, 68–75.
- Köstlin-Gille, N.; Gille, C. Myeloid-Derived Suppressor Cells in Pregnancy and the Neonatal Period. Front. Immunol. 2020, 11, 584712.
- Wegmann, T.G.; Lin, H.; Guilbert, L.; Mosmann, T.R. Biderectional cytokine interactions in the maternal-fetal relationship: Is successful pregnancy a TH2 phenomenon ? Immunol Today. 1993, 14, 5–8.
- Zhao, A.; Xu, H.; Kang, X.; Zhao, A.; Lu, L. New insights into myeloid-derived suppressor cells and their roles in feto-maternal immune cross-talk. J. Reprod. Immunol. 2016, 113, 35–41.
- Crncic, T.B.; Laskarin, G.; Juretic, K.; Strbo, N.; Dupor, J.; Srsen, S.; Randic, L.; Bouteiller, P.L.; Tabiasco, J.; Rukavina, D. Perforin and Fas/FasL Cytolytic Pathways at the Maternal—Fetal Interface. Am. J. Reprod. Immunol. 2005, 54, 241–248.
- Raghupathy, R. Th 1-type immunity is incompatible with successful pregnancy. Immunol. Today 1997, 18, 478–482.
- Tripathi, S.; Guleria, I. Biomarkers in Fetomaternal Tolerance. Clin. Lab. Med. 2018, 39, 145–156.
- Robinson, D.P.; Klein, S.L. Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm. Behav. 2012, 62, 263–271.
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87.
- Köstlin, N.; Kugel, H.; Spring, B.; Leiber, A.; Marmé, A.; Henes, M.; Rieber, N.; Hartl, D.; Poets, C.F.; Gille, C. Granulocytic myeloid derived suppressor cells expand in human pregnancy and modulate T-cell responses. Eur. J. Immunol. 2014, 44, 2582–2591.
- Sykes, L.; MacIntyre, D.A.; Yap, X.J.; Teoh, T.G.; Bennett, P.R. The Th1:Th2 Dichotomy of Pregnancy and Preterm Labour. Mediat. Inflamm. 2012, 2012, 967629.
- Blencowe, H.; Cousens, S. Review: Addressing the challenge of neonatal mortality. Trop. Med. Int. Health 2013, 18, 303–312.
- Gantt, S.; Gervassi, A.; Jaspan, H.; Horton, H. The Role of Myeloid-Derived Suppressor Cells in Immune Ontogeny. Front. Immunol. 2014, 5, 387.
- Levy, O. Innate immunity of the newborn: Basic mechanisms and clinical correlates. Nat. Rev. Immunol. 2007, 7, 379–390.
- Philbin, V.J.; Levy, O. Developmental Biology of the Innate Immune Response: Implications for Neonatal and Infant Vaccine Development. Pediatr. Res. 2009, 65, 98R–105R.
- Wilson, C.B.; Lewis, D.B. Basis and Implications of Selectively Diminished Cytokine Production in Neonatal Susceptibility to Infection. Clin. Infect. Dis. 1990, 12, S410–S420.
- Kollman, T.R.; Levy, O.; Montgomery, R.R.; Goriely, S. Innate Immune Sensing by Toll-like Receptors in Newborns and the Elderly. Immunity 2012, 37, 771–783.
- Maródi, L. Innate cellular immune responses in newborns. Clin. Immunol. 2006, 118, 137–144.
- Zaghouani, H.; Hoeman, C.M.; Adkins, B. Neonatal immunity: Faulty T-helpers and the shortcomings of dendritic cells. Trends Immunol. 2009, 30, 585–591.
- Rieber, N.; Gille, C.; Köstlin, N.; Schäfer, I.; Spring, B.; Ost, M.; Spieles, H.; Kugel, H.A.; Pfeiffer, M.; Heininger, V.; et al. Neutrophilic myeloid-derived suppressor cells in cord blood modulate innate and adaptive immune responses. Clin. Exp. Immunol. 2013, 174, 45–52.
- Schwarz, J.; Scheckenbach, V.; Kugel, H.; Spring, B.; Pagel, J.; Härtel, C.; Pauluschke-Fröhlich, J.; Peter, A.; Poets, C.F.; Gille, C.; et al. Granulocytic myeloid-derived suppressor cells (GR-MDSC) accumulate in cord blood of preterm infants and remain elevated during the neonatal period. Clin. Exp. Immunol. 2017, 191, 328–337.
- Leiber, A.; Schwarz, J.; Köstlin, N.; Spring, B.; Fehrenbacher, B.; Katava, N.; Poets, C.F.; Gille, C. Neonatal myeloid derived suppressor cells show reduced apoptosis and immunosuppressive activity upon infection with Escherichia coli. Eur. J. Immunol. 2017, 47, 1009–1021.
- Gervassi, A.; Lejarcegui, N.; Dross, S.; Jacobson, A.; Itaya, G.; Kidzeru, E.; Gantt, S.; Jaspan, H.; Horton, H. Myeloid Derived Suppressor Cells Are Present at High Frequency in Neonates and Suppress In Vitro T Cell Responses. PLoS ONE 2014, 9, e107816.
- Muniraman, H.; Sardesai, T.; Sardesai, S. Disorders of the Umbilical Cord. Pediatr. Rev. 2018, 39, 332–341.
- Taghizadeh, R.; Cetrulo, K.; Cetrulo, C. Wharton’s Jelly stem cells: Future clinical applications. Placenta 2011, 32, S311–S315.
- Rogers, I.; Casper, R.F. Umbilical cord blood stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 893–908.
- Gluckman, E. Umbilical cord blood biology and transplantation. Curr. Opin. Hematol. 1995, 2, 413–416.
- Mayani, H. Umbilical Cord Blood: Lessons Learned and Lingering Challenges after More Than 20 Years of Basic and Clinical Research. Arch. Med Res. 2011, 42, 645–651.
- He, Y.-M.; Li, X.; Perego, M.; Nefedova, Y.; Kossenkov, A.V.; A Jensen, E.; Kagan, V.E.; Liu, Y.-F.; Fu, S.-Y.; Ye, Q.-J.; et al. Transitory presence of myeloid-derived suppressor cells in neonates is critical for control of inflammation. Nat. Med. 2018, 24, 224–231.
- Dietz, S.; Schwarz, J.; Vogelmann, M.; Spring, B.; Molnár, K.; Orlikowsky, T.W.; Wiese, F.; Holzer, U.; Poets, C.F.; Gille, C.; et al. Cord blood granulocytic myeloid-derived suppressor cells impair monocyte T cell stimulatory capacity and response to bacterial stimulation. Pediatr. Res. 2019, 86, 608–615.

