 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Stephanie Polutchko | + 4399 word(s) | 4399 | 2022-02-23 05:04:01 | | | |
| 2 | Amina Yu | + 2 word(s) | 4401 | 2022-02-28 03:56:17 | | |
Video Upload Options
Improvement of crop climate resilience will require an understanding of whole-plant adaptation to specific local environments. Arabidopsis thaliana ecotypes adapted to local environments with different climates in Sweden and Italy exhibited different responses when grown under common cool temperature. A proportionally greater emphasis on maintenance of photosynthetic activity in the Swedish ecotype contrasted with a greater emphasis on downregulation of light-harvesting and upregulation of antioxidant enzymes in the Italian ecotype under these conditions. The Swedish ecotype thus pushes forward at full speed with productivity under low temperature, whereas the Italian ecotype stays safe from harm while letting productivity decline when temperatures are transiently cold. Either strategy offer directions for the development of climate-resilient crops for specific locations of cultivation.
1. Introduction
1.1. Climate Resilience
The need to develop “climate-resilient crops for improving global food security and safety” is intensifying [1] as extreme weather events become more common, with greater heat in the summer, more frequent and lasting droughts that can extend through more than one season, as well as late-spring/early-fall cold spells [2]. Climate-resilient crops are needed that maintain high productivity despite less predictable weather patterns and under unique combinations of environmental stresses[3]. Even evasive agricultural strategies require the concomitant development of new crop lines. For example, early sowing of summer crops is used to shift crop production to cooler months of the year, thus evading the most intense summer heat/drought and improving water-use efficiency [4]. Another evasive strategy involves the relocation of agricultural operations (as well as migration of natural plant communities) to higher latitudes where plants experience cooler temperatures and longer photoperiods in the summer [5][6][7][8]. Crops utilized for these strategies must be capable of high productivity in environments with varying levels of exposure to cold temperatures. Even in regions where freezing temperatures are uncommon, occasional freezing events must be tolerated. Moreover, of specific importance for productivity is the ability to make adjustments that allow maintenance of high photosynthesis rates under cool temperatures, which is the focus of this review.
Plants that grow as winter annuals germinate in the fall and overwinter before reproducing in the spring. As such, winter annuals must be able to withstand freezing temperatures [9][10][11] as well as maintain high photosynthetic activity under cool temperatures. Photosynthetic enzymes work more slowly at lower temperature, and the same presumably applies to proteins associated with sugar export from leaves. To compensate, plant acclimation to low growth temperature involves the development of new leaves with greater maximal photosynthetic capacity as a result of greater levels of photosynthetic proteins [12][13][14] and presumably transport proteins. Such a corresponding increase in the number of sugar-export proteins was proposed to take place in winter annuals during acclimation to cool temperature [15]. This acclimation also involved a major infrastructural change at the leaf level, with thicker leaves containing additional layers of chloroplast-rich palisade cells [16][17][18], as is also seen in other herbaceous species that are active during the winter [16][19][20][21][22]. These leaf morphological adjustments are accompanied by larger (sugar-loading) minor leaf veins containing a greater number of sugar-loading cells [16][17][23][24][25][26].
These leaf-level anatomical and morphological adjustments of winter annuals in response to cool growth temperatures presumably allow maximization of the number of photosynthetic and transport proteins per unit of leaf area. In contrast, summer annuals exhibit neither pronounced photosynthetic upregulation nor an increase in palisade layer number or larger minor veins when grown under cool versus warm temperatures[16].
1.2. Comparative Ecophysiology and the Study of Phenotypic Plasticity
The use of comparative ecophysiology (a compare-and-contrast approach) to traits of species or populations adapted to contrasting environments was championed by the Department of Plant Biology of the Carnegie Institution of Washington (for example,[12][27][28][29][30][31]). The studies summarized here built on the work of Doug Schemske and Jon Ågren who had picked the specific locations (Figure 1 [32]) and the specific SW (Rodasen-47; from Sweden) and IT (Castelnuovo-12; from Italy) lines of A. thaliana that are the focus of this review. After Ågren and Schemske [32] published evidence of local adaptation from reciprocal transplants, establishing these populations as true “ecotypes” (see [33]), their teams provided valuable and detailed characterizations of these plants under field conditions [34][35][36][37][38][39][40][41], including their growing seasons with the timing of germination, flowering, and seed-set, which turned out to be quite different.
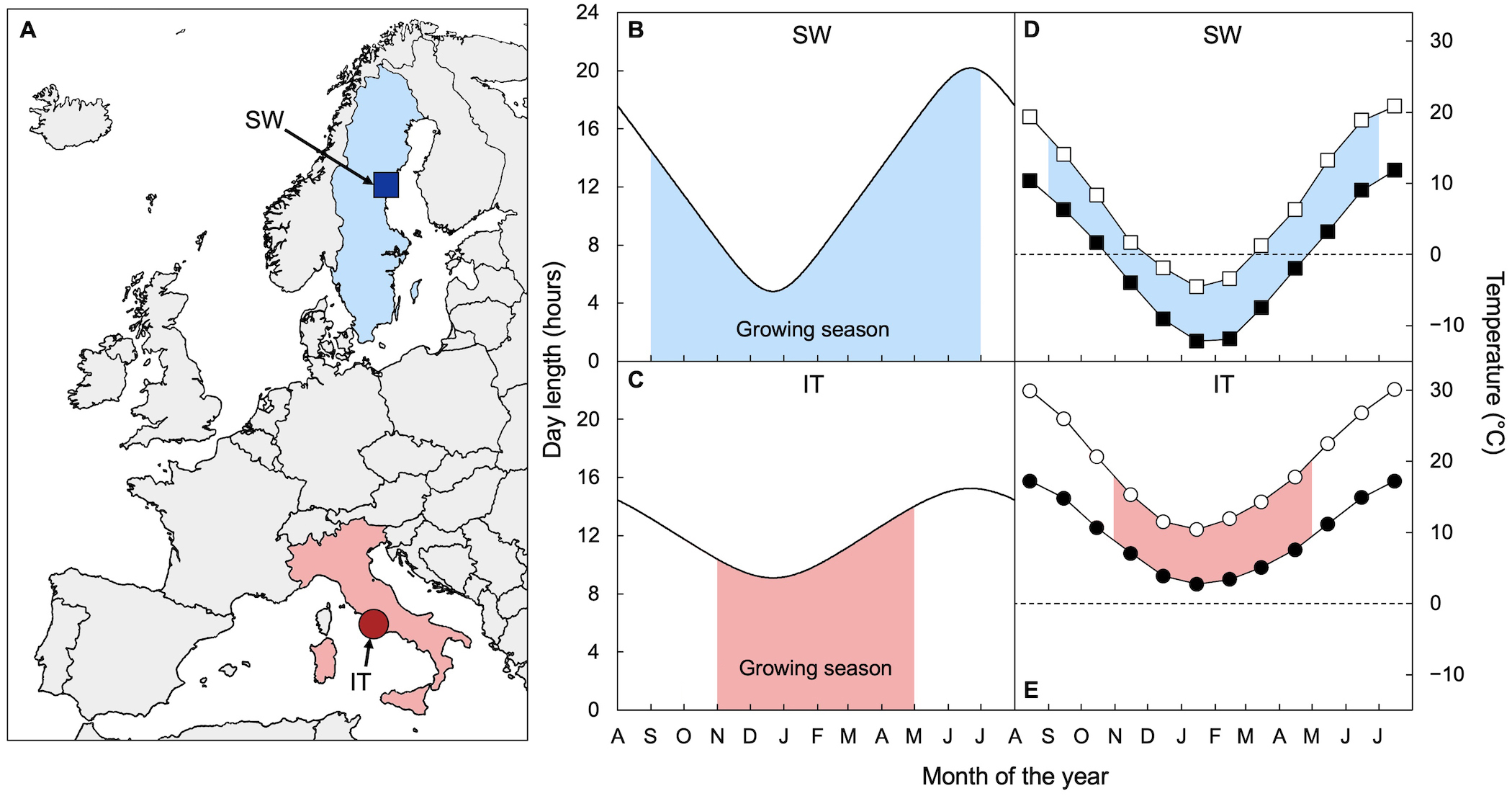
Figure 1. (A) Map showing the geographic origins of the SW (blue square; 62°48′ N, 18°12′ E) and IT (red circle; 42°07′ N, 12°29′ E) A. thaliana ecotypes (after [32]) as well as (B,C) the change in day length and (D,E) monthly minimal (closed symbols) and maximal (open symbols) temperatures across the growing season (light blue or red shading) for (B,D) SW and (C,E) IT at their respective sites of origin. Data on growing seasons for the two ecotypes from [35][41].
Overwintering annuals must contend with the prospect of freezing events in many of the environments in which they are found. The Sweden (SW) ecotype faces subfreezing average monthly minimum temperatures in its native habitat for multiple winter months, but Italy (IT) does not face any subfreezing average monthly minimum temperatures in its native habitat (Figure 1). Perhaps not surprisingly, SW exhibits significantly greater freezing tolerance than IT when grown under common controlled conditions [42][43][44][45][46][47].
The other key element to plant success is the ability to gain enough carbon to support reproduction during periods conducive to productivity [48][49][50]. IT is exposed to average monthly maximal temperatures that remain above 10°C throughout its life cycle in its native habitat (Figure 1), including the winter months between seedling establishment in early November and seed maturation in late April [41]. On the other hand, Sweden (SW) exhibits an extension of its life cycle, with seedlings establishing much earlier (early September [41]) and seeds maturing much later (late June [41]) than IT (Figure 1; see also [36]). As a result, SW and IT experience similar temperatures during corresponding developmental stages (following germination and before reproduction) despite different lifespans. These ecotypes clearly shrink the differences in environmental conditions they have to contend with for productivity by virtue of their different phenology. These differences were linked to gene loci associated with delayed flowering time in SW [33][36].
The extension of SW’s life cycle into the summer at high latitude also brings with it exposure to much longer days leading up to reproduction (Figure 1) and further exacerbates the latitude-dependent differences in daylength between the ecotypes’ native habitats. This difference represents a greater amplitude of variation in light supply (via photoperiod) in Sweden, with relatively shorter days in winter and even longer days in summer at the site of origin of SW. Agricultural strategies that involve a move to a higher latitude will likewise expose plants not only to seasonally cooler temperatures but also to an increased total light supply with longer days in the summer[5][6][7][8].
Findings related to productivity traits for SW and IT point to a greater degree of phenotypic plasticity (wider range of variation in response of individual plant form/function) in these traits in SW, as the ecotype from the habitat with the greater amplitude in temperature and light over the year/growing season when grown under contrasting light intensities and/or temperatures under controlled conditions [17][24][51][52][53][54][55]. These productivity-related traits include photosynthetic capacity and associated processes of sugar export from the leaf. However, a different suite of traits exhibited the opposite pattern—greater upregulation of genes that have antioxidant functions in removing reactive oxygen species (ROS) in IT versus SW for comparison of plants grown under low light and warm temperature (LLW) versus high light and cool temperature (HLC)[47].
The study of phenotypic plasticity in locally adapted populations of other species revealed similarly diverse patterns. It was concluded that the degree of plasticity varies trait by trait among ecotypes [56][57], indicative of the diverse strategies plants employ to survive in specific environmental contexts [58], which may make “local studies … more informative than global ones”[59]. Stearns [60] suggested revising the assessment of traits as either positive or negative by updating the concept of tradeoffs to include a more fluid view of “dynamic linkages.” Sack and Buckley [61] also emphasized the need to consider interactions among multiple genes/traits. Combining genomic/transcriptomic information with information on the response of functional traits may further aid in gaining a holistic understanding of plant response [62]. It was proposed that an essential next step in the quest to understand how plants respond to different environments is the identification of underlying themes of how genotypes interact with the environment [63].
In the present review, evidence from multiple studies on the pair of IT and SW ecotypes is summarized in a schematic model (Figure 2) that integrates the responses of suites of traits related to various aspects of photosynthetic productivity and/or protection against oxidative stress in SW versus IT. The schematic depiction in Figure 2 presents these functional features at a glance, positing that SW effectively minimizes excitation pressure and the associated formation of ROS in the chloroplast by pronounced upregulation of the capacity for photosynthetic electron transport, the capacity for preemptive dissipation of excess excitation as heat (thermal energy), and the capacity to export sugar from source leaves (Figure 2A). In contrast, it is proposed that IT shifts its emphasis, with a somewhat less pronounced capacity for photosynthetic upregulation, pre-emptive dissipation of excess excitation, and export of sugars from source leaves to a proportionally greater emphasis on alternative approaches, including a smaller light-harvesting antenna and pronounced upregulation of multiple antioxidant enzymes that may prevent disruption of cellular redox homeostasis despite formation of more ROS (Figure 2B).
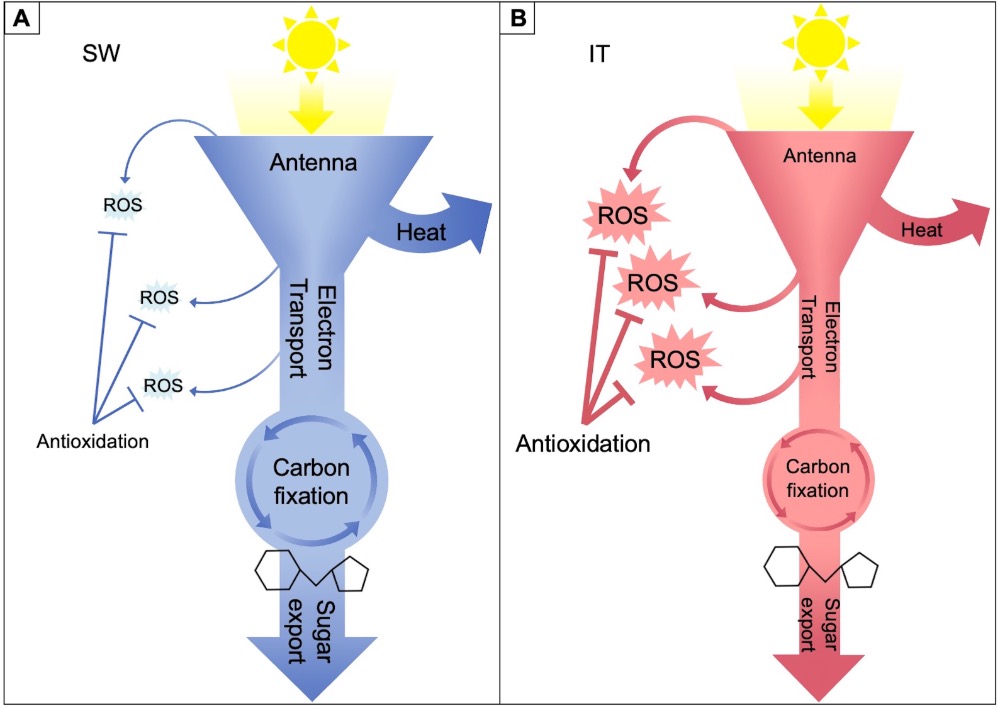
Figure 2. Schematic depiction of relative differences in capacities for light-harvesting (antenna), photoprotective dissipation of excess excitation energy as heat, electron transport, carbon dioxide fixation, sugar export (sucrose symbol), and ROS production and detoxification in leaves of the (A) SW and (B) IT ecotypes of A. thaliana.
IT’s different acclimation compared to that of SW may be well suited for IT’s native environment with relatively consistent and mild weather during the plant’s growth cycle. These warm periods may only be interrupted by occasional brief cold periods, followed by rapid resumption of photosynthetic activity when temperatures rise again. Plant productivity in such a scenario may not require the same pronounced up-regulation of maximal photosynthetic capacity as seen in SW. In contrast, the continuously cold conditions in SW’s native habitat may require maximization of the up-regulation of photosynthetic capacity and maintenance of a larger light-harvesting antenna along with a very high capacity for flexible preemptive dissipation of excess excitation as heat from this antenna as needed.
In addition, the capacity for dissipating excess absorbed light (excess excitation energy that can be potentially destructive) as harmless heat energy (Figure 2) was greater in SW versus IT [45]. It should be noted that excess light results from an imbalance in light level (for example, input of excitation) versus temperature (as affecting utilization of excitation), resulting in excess excitation pressure under very high or fluctuating light even in the presence of moderate temperature (or modestly cool temperatures above the chilling range) or, conversely, under colder temperatures in the presence of low light levels.
As a result of the greater utilization of excitation energy in photosynthesis and the greater capacity of alternative dissipation of excess excitation as heat, overall excitation pressure in the chloroplast [47] was lower in HLC-grown SW compared to IT under exposure to high light levels. This finding further supports the model shown in Figure 2 and suggests that higher levels of ROS may be formed in IT versus SW under these conditions.
2. Differential Upregulation of Photosynthesis and Photoprotection, Sugar-Export Infrastructure, and Expression of Relevant Genes
2.1. Functional and Anatomical Traits
This section focuses on traits of IT and SW associated with plant productivity, especially the maintenance of photosynthetic activity under cool temperatures in fall and spring. These features are compared in plants of SW and IT that had developed under, and were fully acclimated to, various combinations of temperature and light intensity under controlled growth conditions. As suggested in Figure 2, SW exhibits more pronounced upregulation of photosynthesis than IT (Figure 3A) when grown under a combination of cool temperature and high light [47], moderate temperature and high light [17], or cool temperature and moderate light [53]. SW also maintains a larger light-harvesting antenna than IT under the first two growth conditions, as evidenced by a lower chlorophyll a/b ratio (Figure 3B).
Expression of photosynthetic genes responds to sink activity [64], for example, the combined consumption of sugars (produced in photosynthesis) by growing and metabolizing tissues and/or storage in storage organs [50]. Maintenance of a high photosynthetic capacity thus requires a high growth rate and/or a high storage capacity. Overwintering annuals limit their above-ground vegetative growth during the winter and presumably store carbohydrates produced in photosynthesis on mild winter days to support a quick resumption of growth in the spring.
Sugar production and movement out of the leaves requires sugar loading and export as well as water transport into the leaf to replace water lost in transpiration and also move water into foliar phloem in support of the outward-flow of sugar sap [24][26][60][65][66]. When grown under cool temperature, SW exhibits more pronounced upregulation than IT of the foliar vascular infrastructure for moving sugars out of the leaf that match the trends seen for photosynthetic capacity and antenna size. Similar differences were also reported for plants of SW compared to IT grown under moderate temperatures in high light intensity [17] and cool temperatures in moderate light [53]. The finding of a greater sucrose export capacity in SW versus IT is consistent with a greater level of expression of sucrose synthase in plants of SW versus IT grown under HLC [47].
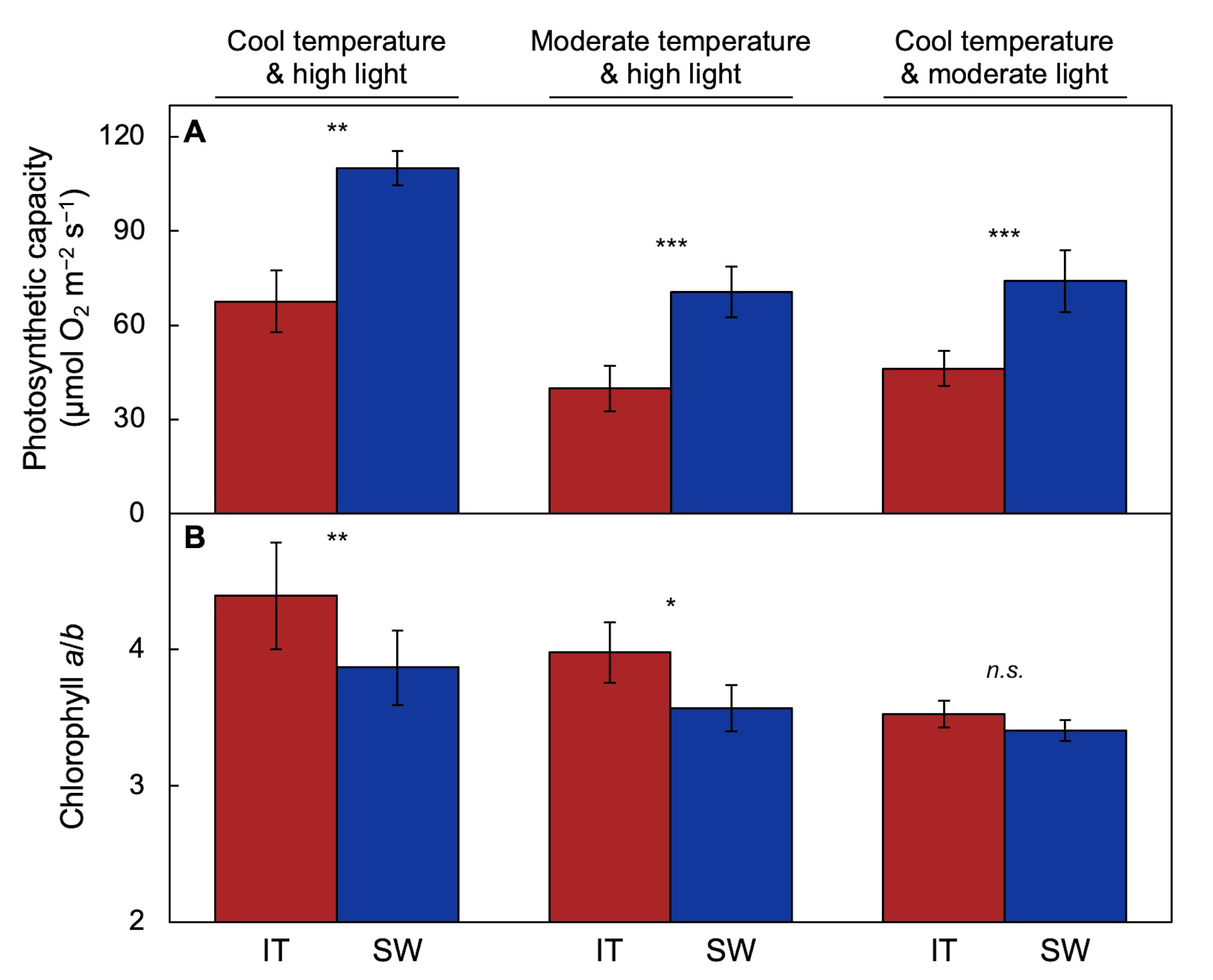
Figure 3. (A) Light- and CO2-saturated capacity of photosynthesis measured at 25°C and (B) chlorophyll a/b ratio in IT (red columns) and SW (blue columns) plants grown under cool temperature and high light (data from [47]), moderate temperature and high light (data from [54]), and cool temperature and moderate light (data from [24][52]). Mean values ± standard deviations (n = 3 or 4); statistically significant differences between ecotypes based on Student’s t-tests are indicated with asterisks (* = p < 0.05; ** = p < 0.01; *** = p < 0.001); n.s. = not significantly different.
These findings indicate that the pronounced acclimation that allowed maintenance of photosynthetic capacity and sucrose export from leaves involved not only greater numbers of photosynthetic and transport proteins but also an expansion of the available leaf infrastructure to accommodate additional chloroplasts and additional vascular cells with sugar-loading transporters.
2.2. Differential Gene Expression
Differential gene regulation patterns were identified for IT and SW in plants grown under HLC compared to LLW [47]. These differences included significant downregulation of several light-harvesting chlorophyll (a + b)-binding genes in HLC-grown IT but no significant downregulation in SW in most of these genes. This finding is consistent with the chlorophyll a/b ratios shown in Figure 4 that indicate a smaller antenna size in HLC-grown IT versus SW. Moreover, consistently more pronounced upregulation in HLC versus LLW was seen in IT for a host of genes encoding antioxidant enzymes that serve in ROS detoxification [47][67].
In contrast, in SW, some of these genes either did not exhibit significant upregulation or were upregulated to a lesser extent in HLC-grown versus LLW-grown plants. A more pronounced upregulation of antioxidant enzymes in HLC-grown versus LLW-grown IT is consistent with the model in Figure 2, suggesting higher levels of ROS production resulting from a higher excitation pressure under high light exposure in IT versus SW that is apparently addressed by pronounced ROS detoxification in IT. Several additional genes with roles in maintaining redox homeostasis are highlighted by Baker et al. [47].
These findings support a proportionally greater emphasis on the mitigation of oxidative stress in IT as the ecotype that exhibits a lesser combined capacity for utilization (in electron transport) and pre-emptive dissipation (as heat) of excitation energy than SW (see also Figure 2). This finding, furthermore, suggests that antenna size in IT is not sufficiently downsized to keep ROS formation from rising more in IT than in SW. The concomitant upregulation of ROS detoxification may offer the benefit of appreciable light absorption and utilization when possible (under warmer temperatures) while maintaining redox homeostasis by virtue of ROS detoxification when needed (during a cold spell).
One can think of this difference in the strategies of SW and IT as SW opening a pre-emptive overflow valve (dissipation of excess energy as heat) in the bathtub, while IT waits to see if the tub overflows (with ROS) and then simply mops the floor with a stack of absorbent towels (ROS-detoxifying antioxidant enzymes). It is tempting to speculate that this difference is associated with the difference in frequency of how often excitation energy tends to become excessive (for example, how frequently the bathtub threatens to overflow) in the ecotypes’ respective native habitats.
3. Growth Patterns as Affected by Growth Environment and Genotype
Winter annuals slow their growth under short days and cool temperatures, which is thought to be protective in limiting frost damage to overwintering shoots [68]. This section focuses on selected findings from transcriptomic analysis. SW and IT are characterized as differing in a transcription factor implicated in cold-temperature acclimation [43], as also noted for other A. thaliana populations from colder versus warmer regions [11][69][70].The transcription factor is a member of the C-repeat-binding factor (CBF) family of transcription factors (CBF1, CBF2, CBF3; hereafter CBF1–3) that support cold tolerance and high photosynthetic rates in plants grown under cool temperatures and/or high light [48][49] and are related to plant fitness in the native habitats of SW and IT [34] as well as other A. thaliana populations across temperature gradients [11][69].
Subsequent genetic analyses revealed that IT has a mutation in the CBF2 gene that makes the corresponding protein nonfunctional [43]. In addition, there are considerably fewer genes in the CBF1–3 regulon in IT versus SW [44]. However, from studies with CBF1–3-deficient lines of IT and SW [44][57], gene regulators other than CBF1–3 transcription factors also play a substantial role in orchestrating the acclimation of both IT and SW to cold temperatures with respect not only to freezing tolerance [44][47] but also photosynthetic acclimation [47]. Likewise, CBF2-deficient lines of SW continued to exhibit greater freezing tolerance than IT [44][46].
SW and IT both exhibited slow growth under cool temperatures (leaf temperatures of 12–16°C), with a significant, albeit rather small, difference between IT and SW (slightly smaller rosette in SW [47]). Likewise, plants grown under an even colder temperature of 4°C showed a similar difference between IT and SW both in rosette area and dry biomass [44], again with SW exhibiting slower growth. The slowing of growth under cool temperature was significantly released in the CBF1–3-deficient mutant of IT but to a lesser (albeit significant) extent in the CBF1–3-deficient mutant of SW [44][47]. Many of the phenotypic acclimatory responses to cold temperature and/or high light were still seen in CBF1–3-deficient mutants, again indicating that other regulators can fulfill the role of the CBF1–3 transcription factors [44][47]. More research is needed to assess the role of CBF2 and other regulators in the ability of IT to maintain a similar growth suppression as seen in SW under cool growth temperatures and, unlike SW, release growth suppression under hot growth temperatures.
In stark contrast, the difference in growth between wildtype IT and SW was dramatic under either low versus high growth light intensity (100 vs. 1000 µmol photons m−2 s−1) under moderate temperature [17] or under hot temperature (leaf temperature of 36°C during the light period and 25°C during the dark period, respectively) versus cool temperature (leaf temperatures of 14°C during the light period and 12.5°C during the dark period, respectively) under moderate light during plant growth [53]. In either case (low growth light or hot growth temperature), IT exhibited dramatically larger rosettes that were twice the size as those of SW. Again, no such differences were seen in high light or cool temperatures [17][53].
The take-home message from these latter findings is that both IT and SW exhibited the expected, and presumably protective, stunting of growth under cool growth temperatures, but that only IT (and not SW) released growth inhibition under hot temperatures. These differences can be viewed in the context of the different environmental conditions during the growing season of the two ecotypes in their respective native habitats. Growth suppression during the short winter season in Italy should protect IT against damage during any freezing episodes that are rare events in IT’s native habitat [32]. Resumption of rapid growth in early spring should aid in the completion of IT’s life cycle before the summer. Conversely, even mid-summer conditions in Sweden do not include any particularly warm temperatures, and an ability to speed up growth and hasten life-cycle completion in response to rising temperatures would appear to be of no benefit to SW.
4. Summary and Conclusions in the Context of Climate Resilience and Dynamic Linkages
As summarized in the schematic model shown in Figure 2, SW exhibited particularly strong upregulation of features that support the maintenance of photosynthetic productivity under cool temperatures, thereby effectively limiting increases in excitation pressure and the associated formation of excess ROS. These features of SW included a greater upregulation of the capacity to utilize excitation energy via photosynthetic electron transport, of preemptive dissipation of excess excitation as heat, and of the foliar vascular infrastructure for sugar export. In addition, SW largely maintained its antenna size when grown under HLC. Conversely, IT exhibited significant downregulation of its light-harvesting antenna size and a proportionally greater upregulation of a broad range of enzymatic antioxidation processes that counter oxidative stress under HLC compared to LLW.
Concerning growth rates, SW and IT exhibited a similar slowing of growth under cool temperature, but not under warm/hot growth temperature, where IT but not SW released the growth arrest. Each ecotype thus demonstrates specific productivity-related features of life cycle progression in their natural habitat and photosynthetic performance and growth under common environmental conditions. The adaptations in leaf form and function seen in SW presumably offer (i) particularly strong support for the maintenance of photosynthetic activity and export of sugar from leaves under continuously cool temperature, which minimizes excess-ROS formation, and (ii) protection against frost damage of above-ground tissue by capping rosette growth.
Conversely, the adaptations of IT from a native habitat with extended periods of warmer temperatures during winter involve releasing constraints on growth under warm temperature and/or low light while still maintaining growth caps under cool temperature. IT’s response to growth under HLC involves a combination of downsizing of light-harvesting antenna size (for example., less capture of light for utilization in photosynthesis) and proportionally more pronounced upregulation of ROS-detoxifying antioxidation processes.
It is to integrate traits/genes and place them into the context of plant performance in specific environments. In particular, this analysis suggests that the lesser degree of freezing tolerance and the lesser upregulation of photosynthetic capacity in IT should not be viewed as inferior to that of SW. It rather appears that the different strategies of IT could be seen as highly suitable for an environment with a low probability of freezing events and substantial periods of time for photosynthetic productivity under temperatures above the chilling range. Particularly strong upregulation of an array of ROS-detoxifying antioxidant enzymes under exposure to cool temperature can be seen as an effective and efficient way to avert oxidative damage if/when needed while tolerating a dip in photosynthetic productivity during a brief cold spell in favor of the resumption of photosynthesis in the subsequent warmer period. The acceleration of growth during rising temperatures may have benefits in environments that experience summer heat. Overall, the difference between IT and SW could be characterized metaphorically as SW pushing forward at full speed with productivity under cool temperatures and IT staying safe from harm while taking a bit of a break from productivity when temperatures are cool.
In conclusion, both ecotypes undergo acclimation to growth under cool temperature that meets the criterion for full acclimation as a transition from an experience of “stress” (with a temporary loss of redox balance resulting in oxidative stress) to a new steady-state in which little to no endogenous stress is experienced (with restored redox balance), and where excessive ROS levels are avoided by a combination of pre-emptive mechanisms that counteract ROS formation (less light absorption and/or boosted utilization/thermal dissipation of excitation energy) and removal of ROS once formed (detoxification by antioxidant enzymes). The difference between IT and SW lies in the relative proportion to which pre-emptive and detoxification mechanisms are employed to reach acclimation.
Preliminary lessons from this insight into productivity-related traits of IT and SW for agriculture include:
- The movement of agriculture to a cooler time of the year or a higher latitude, as an increasingly discussed agricultural approach [5][6][7][8], should be accompanied by crop development for each specific scenario. The design of climate-resilient crops should take into consideration the specific target latitude and associated degree of environmental variability.
- The development of improved crops for cultivation at high latitudes (with continuously low temperatures) should probably follow the acclimation pattern of SW, whereas improved crops for mid-latitudes (where cold spells are only intermittent) should probably follow the acclimation pattern of IT.
- Whereas it was focused on the comparison of two ecotypes of a winter annual, a comparison of either of these with summer annuals may be informative. It should be assessed whether IT, but not SW, combines traits of a winter annual (upregulation of freezing tolerance and photosynthetic and sugar-export capacity under cool growth temperatures) with those of a summer annual. IT’s higher growth rate in warm temperatures supports such a notion.
- More generally, environments with continuous exposure to cool temperatures that dramatically lower productivity in unacclimated plants may be best met with acclimatory adjustments of plant form and function supporting the maintenance of photosynthetic productivity and sugar export from source leaves, thereby pre-emptively minimizing shifts in cellular redox homeostasis. In contrast, environments with extended periods of warmer temperatures and the possibility of only brief interspersed cold spells may be adequately met with a relatively greater emphasis on upregulation of genes with roles in the maintenance of cellular redox homeostasis during intermittent periods of stress. Pronounced growth acceleration when temperatures rise in the spring may play a role in evasion of summer heat.
- Moreover, the trajectory from initial plant exposure to cold temperatures all the way to full acclimation (with leaves newly grown and fully developed under these conditions) likely involves a more pronounced transient upregulation of the antioxidation processes even in genotypes that eventually fully re-establish photosynthetic productivity. Leaves of winter annuals may thus rely to a relatively lesser degree on management rather than avoidance of oxidative stress in fully acclimated new leaves that have developed under cool growth temperatures.
References
- Dhankher, O.P.; Foyer, C.H. Climate resilient crops for improving global food security and safety. Plant. Cell Environ. 2018, 41, 877–884, doi:10.1111/pce.13207.
- Intergovernmental Panel on Climate Change. Available online: http://www.ipcc.ch (accessed on 23 December 2021).
- Rivero, R.M.; Mittler, R.; Blumwald, E.; Zandalinas, S.I. Developing climate‐resilient crops: Improving plant tolerance to stress combination. Plant J. 2022, 109, 373–389, doi:10.1111/tpj.15483.
- Soriano, M.A.; Orgaz, F.; Villalobos, F.J.; Fereres, E. Efficiency of water use of early plantings of sunflower. Eur. J. Agron. 2004, 21, 465–476, doi:10.1016/j.eja.2004.07.001.
- Vitt, P.; Havens, K.; Kramer, A.T.; Sollenberger, D.; Yates, E. Assisted migration of plants: Changes in latitudes, changes in attitudes. Biol. Conserv. 2010, 143, 18–27, doi:10.1016/j.biocon.2009.08.015.
- Hu, Y.; Fan, L.; Liu, Z.; Yu, Q.; Liang, S.; Chen, S.; You, L.; Wu, W.; Yang, P. Rice production and climate change in Northeast China: Evidence of adaptation through land use shifts. Environ. Res. Lett. 2019, 14, 024014, doi:10.1088/1748-9326/aafa55.
- Carlson-Nilsson, U.; Aloisi, K.; Vågen, I.M.; Rajala, A.; Mølmann, J.B.; Rasmussen, S.K.; Niemi, M.; Wojciechowska, E.; Pärs-sinen, P.; Poulsen, G.; et al. Trait expression and environmental eesponses of pea (Pisum sativum L.) genetic resources targeting cultivation in the Arctic. Front. Plant Sci. 2021, 12, 1476, doi:10.3389/fpls.2021.688067.
- Mølmann, J.B.; Dalmannsdottir, S.; Hykkerud, A.L.; Hytönen, T.; Samkumar, A.; Jaakola, L. Influence of Arctic light condi-tions on crop production and quality. Physiol. Plant 2021, 172, 1931–1940, doi:10.1111/ppl.13418.
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599, doi:10.1146/annurev.arplant.50.1.571.
- Zhen, Y.; Ungerer, M.C. Clinal variation in freezing tolerance among natural accessions of Arabidopsis thaliana. New Phytol. 2008, 177, 419–427, doi:10.1111/j.1469-8137.2007.02262.x.
- Kang, J.; Zhang, H.; Sun, T.; Shi, Y.; Wang, J.; Zhang, B.; Wang, Z.; Zhou, Y.; Gu, H. Natural variation of C‐repeat‐binding fac-tor (CBFs) genes is a major cause of divergence in freezing tolerance among a group of Arabidopsis thaliana populations along the Yangtze River in China. New Phytol. 2013, 199, 1069–1080, doi:10.1111/nph.12335.
- Berry, J.; Bjorkman, O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant. Physiol. 1980, 31, 491–543, doi:10.1146/annurev.pp.31.060180.002423.
- Hüner, N.P.A.; Öquist, G.; Sarhan, F. Energy balance and acclimation to light and cold. Trends Plant. Sci. 1998, 3, 224–230, doi:10.1016/S1360-1385(98)01248-5.
- Savitch, L.V.; Leonardos, E.D.; Krol, M.; Jansson, S.; Grodzinski, B.; Hüner, N.P.A.; Öquist, G. Two different strategies for light utilization in photosynthesis in relation to growth and cold acclimation. Plant. Cell Environ. 2002, 25, 761–771, doi:10.1046/j.1365-3040.2002.00861.x.
- Adams, W.W., III; Stewart, J.J.; Demmig-Adams, B. Photosynthetic modulation in response to plant activity and environment. In The Leaf: A Platform for Performing Photosynthesis, Advances in Photosynthesis and Respiration; Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 44, pp. 493–563, doi:10.1007/978-3-319-93594-2_18.
- Cohu, C.M.; Muller, O.; Adams, W.W., III; Demmig-Adams, B. Leaf anatomical and photosynthetic acclimation to cool tem-perature and high light in two winter versus two summer annuals. Physiol. Plant. 2014, 152, 164–173, doi:10.1111/ppl.12154.
- Stewart, J.J.; Polutchko, S.K.; Adams, W.W., III; Cohu, C.M.; Wenzl, C.A.; Demmig‐Adams, B. Light, temperature and tocoph-erol status influence foliar vascular anatomy and leaf function in Arabidopsis thaliana. Physiol. Plant. 2017, 160, 98–110, doi:10.1111/ppl.12543.
- Gorsuch, P.A.; Pandey, S.; Atkin, O.K. Temporal heterogeneity of cold acclimation phenotypes in Arabidopsis leaves. Plant. Cell Environ. 2010, 33, 244–258, doi:10.1111/j.1365-3040.2009.02074.x.
- Boese, S.R.; Hüner, N.P.A. Effect of growth temperature and temperature shifts on spinach leaf morphology and photosynthe-sis. Plant. Physiol. 1990, 94, 1830–1836, doi:10.1104/pp.94.4.1830.
- Adams, W.W., III; Hoehn, A.; Demmig-Adams, B. Chilling temperatures and the xanthophyll cycle. A comparison of warm-grown and overwintering spinach. Funct. Plant. Biol. 1995, 22, 75–85, doi:10.1071/pp9950075.
- Dumlao, M.R.; Darehshouri, A.; Cohu, C.M.; Muller, O.; Mathias, J.; Adams, W.W., III; Demmig-Adams, B. Low temperature acclimation of photosynthetic capacity and leaf morphology in the context of phloem loading type. Photosynth. Res. 2012, 113, 181–189, doi:10.1007/s11120-012-9762-5.
- Muller, O.; Stewart, J.J.; Cohu, C.M.; Polutchko, S.K.; Demmig-Adams, B.; Adams, W.W. III. Leaf architectural, vascular and photosynthetic acclimation to temperature in two biennials. Physiol. Plant. 2014, 152, 763–772, doi:10.1111/ppl.12226.
- Adams, W.W., III; Cohu, C.M.; Muller, O.; Demmig-Adams, B. Foliar phloem infrastructure in support of photosynthesis. Front. Plant. Sci. 2013, 4, 194, doi:10.3389/fpls.2013.00194.
- Cohu, C.M.; Muller, O.; Demmig-Adams, B.; Adams, W.W. III. Minor loading vein acclimation for three Arabidopsis thaliana ecotypes in response to growth under different temperature and light regimes. Front. Plant. Sci. 2013, 4, 240, doi:10.3389/fpls.2013.00240.
- Muller, O.; Cohu, C.M.; Stewart, J.J.; Protheroe, J.A.; Demmig-Adams, B.; Adams, W.W. III. Association between photosynthe-sis and contrasting features of minor veins in leaves of summer annuals loading phloem via symplastic versus apoplastic routes. Physiol. Plant. 2014, 152, 174–183, doi:10.1111/ppl.12155.
- Cohu, C.M.; Muller, O.; Stewart, J.J.; Demmig-Adams, B.; Adams, W.W. III. Association between minor loading vein architec-ture and light- and CO2-saturated rates of photosynthetic oxygen evolution among Arabidopsis thaliana ecotypes from differ-ent latitudes. Front. Plant. Sci. 2013, 4, 264, doi:10.3389/fpls.2013.00264.
- Clausen, J.; Keck, D.D.; Hiesey, W.M. Experimental Studies on the Nature of Species. I. Effect of Varied Environments on Western North. American Plants; Carnegie Institution of Washington: Washington, DC, USA, 1940.
- Clausen, J.; Keck, D.D.; Hiesey, W.M. Regional differentiation in plant species. Am. Nat. 1941, 75, 231–250, doi:10.1086/280955.
- Clausen, J.; Keck, D.D.; Hiesey, W.M. Heredity of geographically and ecologically isolated races. Am. Nat. 1947, 81, 114–133, doi:10.1086/281507
- Hiesey, W.M.; Nobs, M.A.; Björkman, O. Experimental Studies on the Nature of Species. Volume V: Biosystematics, Genetics, and Physiological Ecology of the Erythranthe Section of Mimulus; Carnegie Institution of Washington: Washington, DC, USA, 1971.
- Björkman, O. Comparative studies on photosynthesis in higher plants. In Photophysiology, Current Topics in Photobiology and Photochemistry; Giese, A.C., Ed.; Academic Press: New York, USA, 1973; Volume VIII, pp. 1–63, doi:10.1016/B978-0-12-282608-5.50007-2.
- Ågren, J.; Schemske, D.W. Reciprocal transplants demonstrate strong adaptive differentiation of the model organism Ara-bidopsis thaliana in its native range. New Phytol. 2012, 194, 1112–1122, doi:10.1111/j.1469-8137.2012.04112.x.
- Lowry, D.B. Ecotypes and the controversy over stages in the formation of new species. Biol. J. Linn. Soc. 2012, 106, 241–257, doi:10.1111/j.1095-8312.2012.01867.x.
- Ågren, J.; Oakley, C.G.; McKay, J.K.; Lovell, J.T.; Schemske, D.W. Genetic mapping of adaptation reveals fitness tradeoffs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2013, 110, 21077–21082, doi:10.1073/pnas.1316773110.
- Ågren, J.; Oakley, C.G.; Lundemo, S.; Schemske, D.W. Adaptive divergence in flowering time among natural populations of Arabidopsis thaliana: estimates of selection and QTL mapping. Evolution 2017, 71, 550–564, doi:10.1111/evo.13126.
- Grillo, M.A.; Li, C.; Hammond, M.; Wang, L.; Schemske, D.W. Genetic architecture of flowering time differentiation between locally adapted populations of Arabidopsis thaliana. New Phytol. 2013, 197, 1321–1331, doi:10.1111/nph.12109.
- Postma, F.M.; Ågren, J. Maternal environment affects the genetic basis of seed dormancy in Arabidopsis thaliana. Mol. Ecol. 2015, 24, 785–797, doi:10.1111/mec.13061.
- Postma, F.M.; Ågren, J. Early life stages contribute strongly to local adaptation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 7590–7595, doi:10.1073/pnas.1606303113.
- Postma, F.M.; Ågren, J. Among-year variation in selection during early life stages and the genetic basis of fitness in Arabidop-sis thaliana. Mol. Ecol. 2018, 27, 2498– 2511, doi:10.1111/mec.14697.
- Postma, F.M.; Lundemo, S.; Ågren, J. Seed dormancy cycling and mortality differ between two locally adapted populations of Arabidopsis thaliana. Ann. Bot. 2016, 117, 249–256, doi:10.1093/aob/mcv171.
- Ellis, T.J.; Postma, F.M.; Oakley, C.G.; Ågren, J. Life‐history trade‐offs and the genetic basis of fitness in Arabidopsis thaliana. Mol. Ecol. 2021, 30, 2846–2858, doi:10.1111/mec.15941.
- Oakley, C.G.; Ågren, J.; Atchison, R.A.; Schemske, D.W. QTL mapping of freezing tolerance: Links to fitness and adaptive trade-offs. Mol. Ecol. 2014, 23, 4304–4315, doi:10.1111/mec.12862.
- Gehan, M.A.; Park, S.; Gilmour, S.J.; An, C.; Lee, C.; Thomashow, M.F. Natural variation in the C‐repeat binding factor cold response pathway correlates with local adaptation of Arabidopsis ecotypes. Plant. J. 2015, 84, 682–693, doi:10.1111/tpj.13027.
- Park, S.; Gilmour, S.J.; Grumet, R.; Thomashow, M.F. CBF-dependent and CBF-independent regulatory pathways contribute to the differences in freezing tolerance and cold-regulated gene expression of two Arabidopsis ecotypes locally adapted to sites in Sweden and Italy. PLoS ONE 2018, 13, e0207723, doi:10.1371/journal.pone.0207723.
- Oakley, C.G.; Savage, L.; Lotz, S.; Larson, G.R.; Thomashow, M.F.; Kramer, D.M.; Schemske, D.W. Genetic basis of photosyn-thetic responses to cold in two locally adapted populations of Arabidopsis thaliana. J. Exp. Bot. 2018, 69, 699–709, doi:10.1093/jxb/erx437.
- Sanderson, B.J.; Park, S.; Jameel, M.I.; Kraft, J.C.; Thomashow, M.F.; Schemske, D.W.; Oakley, C.G. Genetic and physiological mechanisms of freezing tolerance in locally adapted populations of a winter annual. Am. J. Bot. 2020, 107, 250–261, doi:10.1002/ajb2.1385.
- Baker, C.R.; Stewart, J.J.; Amstutz, C.L.; Ching, L.G.; Johnson, J.D.; Niyogi, K.K.; Adams, W.W., III; Demmig‐Adams, B. Geno-type‐dependent contribution of CBF transcription factors to long‐term acclimation to high light and cool temperature. Plant. Cell Environ. 2022, 45, 392–411, doi:10.1111/pce.14231.
- Savitch, L.V.; Allard, G.; Seki, M.; Robert, L.S.; Tinker, N.A.; Hüner, N.P.A.; Shinozaki, K.; Singh, J. The effect of overexpres-sion of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica na-pus. Plant. Cell Physiol. 2005, 46, 1525–1539, doi:10.1093/pcp/pci165.
- Hüner, N.P.A.; Dahal, K.; Bode, R.; Kurepin, L.V.; Ivanov, A.G. Photosynthetic acclimation, vernalization, crop productivity and ‘the grand design of photosynthesis’. J. Plant. Physiol. 2016, 203, 29–43, doi:10.1016/j.jplph.2016.04.006.
- Demmig-Adams, B.; Stewart, J.J.; Adams, W.W. III Environmental regulation of intrinsic photosynthetic capacity: An inte-grated view. Curr. Opin. Plant. Biol. 2017, 37, 34–41, doi:10.1016/j.pbi.2017.03.008.
- Stewart, J.J.; Adams, W.W., III; Cohu, C.M.; Polutchko, S.K.; Lombardi, E.M.; Demmig-Adams, B. Differences in light-harvesting, acclimation to growth-light environment, and leaf structural development between Swedish and Italian ecotypes of Arabidopsis thaliana. Planta 2015, 242, 1277–1290, doi:10.1007/s00425-015-2368-3.
- Adams, W.W., III; Stewart, J.J.; Cohu, C.M.; Muller, O.; Demmig-Adams, B. Habitat temperature and precipitation of Ara-bidopsis thaliana ecotypes determine the response of foliar vasculature, photosynthesis, and transpiration to growth temper-ature. Front. Plant. Sci. 2016, 7, 1026, doi:10.3389/fpls.2016.01026.
- Stewart, J.J.; Demmig‐Adams, B.; Cohu, C.M.; Wenzl, C.A.; Muller, O.; Adams, W.W. III Growth temperature impact on leaf form and function in Arabidopsis thaliana ecotypes from northern and southern Europe. Plant. Cell Environ. 2016, 39, 1549–1558, doi:10.1111/pce.12720.
- Stewart, J.J.; Polutchko, S.K.; Adams, W.W., III; Demmig‐Adams, B. Acclimation of Swedish and Italian ecotypes of Arabidop-sis thaliana to light intensity. Photosynth. Res. 2017, 134, 215–229, doi:10.1007/s11120-017-0436-1.
- Adams, W.W., III; Stewart, J.J.; Polutchko, S.K.; Demmig-Adams, B. Leaf vasculature and the upper limit of photosynthesis. In The Leaf: A Platform for Performing Photosynthesis, Advances in Photosynthesis and Respiration; Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 44, pp. 27–54, doi:10.1007/978-3-319-93594-2_2.
- Mason, C.M.; LaScaleia, M.C.; De La Pascua, D.R.; Monroe, J.G.; Goolsby, E.W. Learning from dynamic traits: Seasonal shifts yield insights into ecophysiological tradeoffs across scales from macroevolutionary to intra-individual. Int. J. Plant. Sci. 2020, 181, 88–102, doi:10.1086/706238.
- Westerband, A.C.; Funk, J.L.; Barton, K.E. Intraspecific trait variation in plants: a renewed focus on its role in ecological pro-cesses. Ann. Bot. 2021, 127, 397–410, doi:10.1093/aob/mcab011.
- Salguero-Gómez, R.; Jones, O.R.; Jongejans, E.; Blomberg, S.P.; Hodgson, D.J.; Mbeau-Ache, C.; Zuidema, P.A.; de Kroon, H.; Buckley, Y.M. Fast–slow continuum and reproductive strategies structure plant life-history variation worldwide. Proc. Natl. Acad. Sci. USA 2016, 113, 230–235, doi:10.1073/pnas.1506215112.
- Sulis, E.; Bacchetta, G., Cogoni, D.; Fenu, G. From global to local scale: Where is the best for conservation purpose? Biodivers. Conserv. 2021, 30, 183–200, doi:10.1007/s10531-020-02085-4.
- Stearns, S.C. Trade-offs in life-history evolution. Funct. Ecol. 1989, 3, 259, doi:10.2307/2389364.
- Sack, L.; Buckley, T.N. Trait multi-functionality in plant stress response. Integr. Comp. Biol. 2020, 60, 98–112, doi:10.1093/icb/icz152.
- Van Nuland, M.E.; Vincent, J.B.; Ware, I.M.; Mueller, L.O.; Bayliss, S.L.; Beals, K.K.; Schweitzer, J.A.; Bailey, J.K. Intraspecific trait variation across elevation predicts a widespread tree species' climate niche and range limits. Ecol. Evol. 2020, 10, 3856–3867, doi:10.1002/ece3.5969.
- Saltz, J.B.; Bell, A.M.; Flint, J.; Gomulkiewicz, R.; Hughes, K.A.; Keagy, J. Why does the magnitude of genotype‐by‐environment interaction vary? Ecol. Evol. 2018, 8, 6342–6353, doi:10.1002/ece3.4128.
- Krapp, A.; Stitt, M. An evaluation of direct and indirect mechanisms for the “sink-regulation” of photosynthesis in spinach: Changes in gas exchange, carbohydrates, metabolites, enzyme activities and steady-state transcript levels after cold-girdling source leaves. Planta 1995, 195, 313–323, doi:10.1007/BF00202587.
- Polutchko, S.K.; Stewart, J.J.; Demmig-Adams, B.; Adams, W.W. III. Evaluating the link between photosynthetic capacity and leaf vascular organization with principal component analysis. Photosynthetica 2018, 56, 392–403, doi:10.1007/s11099-017-0764-6.
- Adams, W.W., III; Stewart, J.J.; Polutchko, S.K.; Demmig-Adams, B. Foliar sieve elements: Nexus of the leaf. J. Plant. Physiol. 2022, 269, 153601, doi:10.1016/j.jplph.2021.153601.
- Demmig-Adams, B; Polutchko, S.K.; Baker, C.R.; Stewart, J.J.; Adams W.W., III; Distinct Cold Acclimation of Productivity Traits in Arabidopsis thaliana Ecotypes. International Journal of Molecular Sciences 2022, 23, 2129, 10.3390/ijms23042129.
- Eremina, M.; Rozhon, W.; Poppenberger, B. Hormonal control of cold stress responses in plants. Cell. Mol. Life Sci. 2016, 73, 797–810, doi:10.1007/s00018-015-2089-6.
- Alonso-Blanco, C.; Gomez-Mena, C.; Llorente, F.; Koornneef, M.; Salinas, J.; Martínez-Zapater, J.M. Genetic and molecular analyses of natural variation indicate CBF2 as a candidate gene for underlying a freezing tolerance quantitative trait locus in Arabidopsis. Plant. Physiol. 2005, 139, 1304–1312, doi:10.1104/pp.105.068510.
- Monroe, J.G.; McGovern, C.; Lasky, J.R.; Grogan, K.; Beck, J.; McKay, J.K. Adaptation to warmer climates by parallel functional evolution of CBF genes in Arabidopsis thaliana. Mol. Ecol. 2016, 25, 3632–3644, doi:10.1111/mec.13711.


