 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Md Abdul Hannan | + 4573 word(s) | 4573 | 2022-01-13 03:41:29 | | | |
| 2 | Peter Tang | + 1 word(s) | 4574 | 2022-01-26 02:35:46 | | |
Video Upload Options
Marine algae are considered to be a potential source of some unique metabolites with diverse health benefits. The pharmacological properties, such as antioxidant, anti-inflammatory, cholesterol homeostasis, protein clearance and anti-amyloidogenic potentials of algal metabolites endorse their protective efficacy against oxidative stress, neuroinflammation, mitochondrial dysfunction, and impaired proteostasis which are known to be implicated in the pathophysiology of neurodegenerative disorders and the associated complications after cerebral ischemia and brain injuries.
1. Introduction
2. Pathophysiology of Brain Disorders
2.1. Neurodegenerative Disorders (AD and PD)
2.2. Ischemic Stroke
2.3. Traumatic Brain Injury
3. Neuropharmacological Potentials of Marine Algae and Their Metabolites: Evidence from In Vitro Studies
|
Pharmacological Effects |
Compound (Class) |
Algal Source If Any) |
Effective Concentration |
Experimental Model (In Vivo/In Vitro) |
Cellular Effects/Significant Findings |
Signaling Pathways Involved |
Pharmacological Markers |
Reference |
|---|---|---|---|---|---|---|---|---|
|
Antioxidant activity |
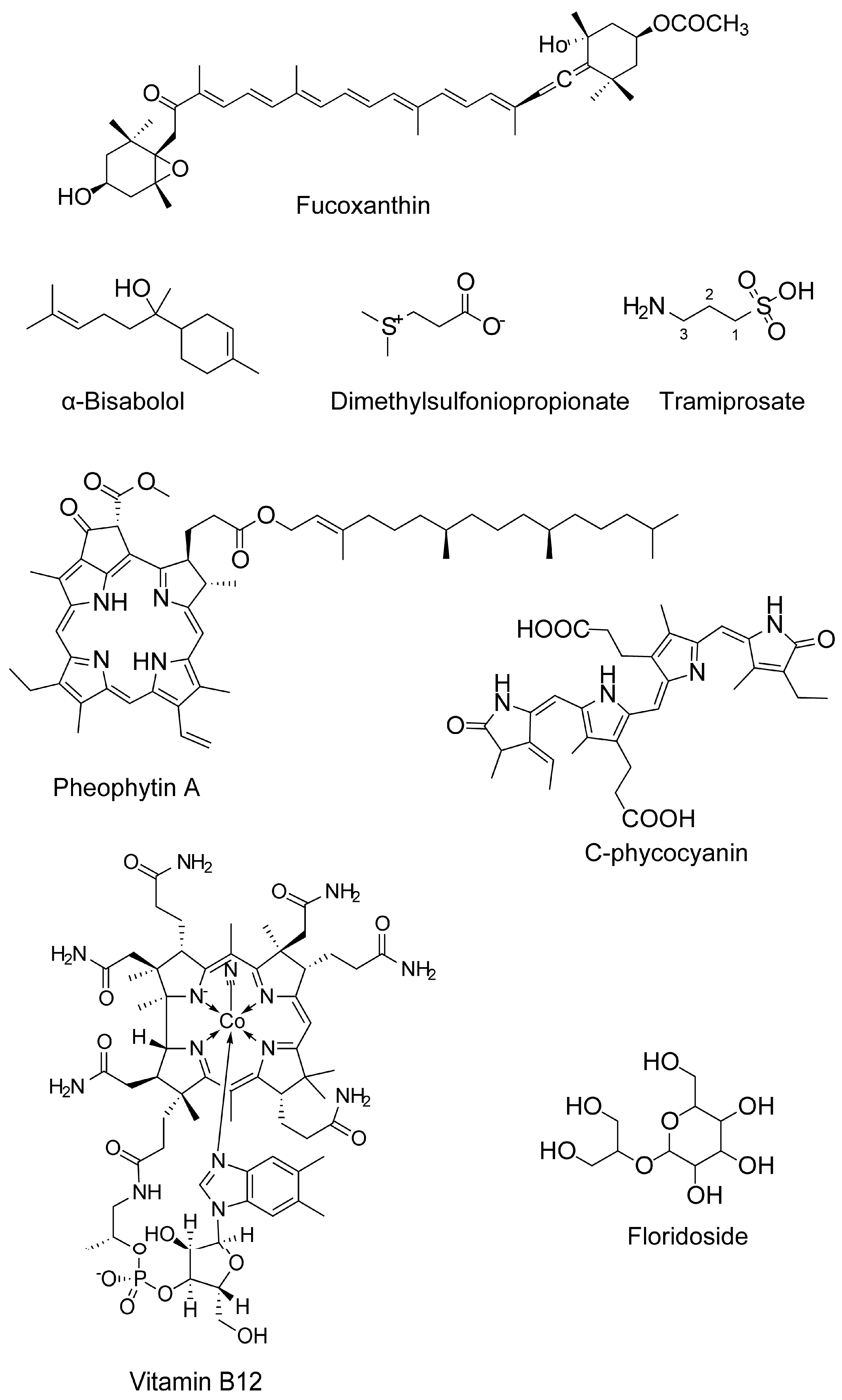
Fucoxanthin (carotenoids) |
Sargassum siliquastrum |
50 and 100 μM |
H2O2-induced cell damage in kidney fibroblast cells |
Attenuates oxidative stress |
n.d. |
↓ROS level |
[69] |
|
Fucoxanthin |
5, 10, and 50 μM |
H2O2 induced BV2 microglial cells |
Antioxidation |
Antioxidant pathway |
↓ROS ↑SOD and GSH |
[36] |
||
|
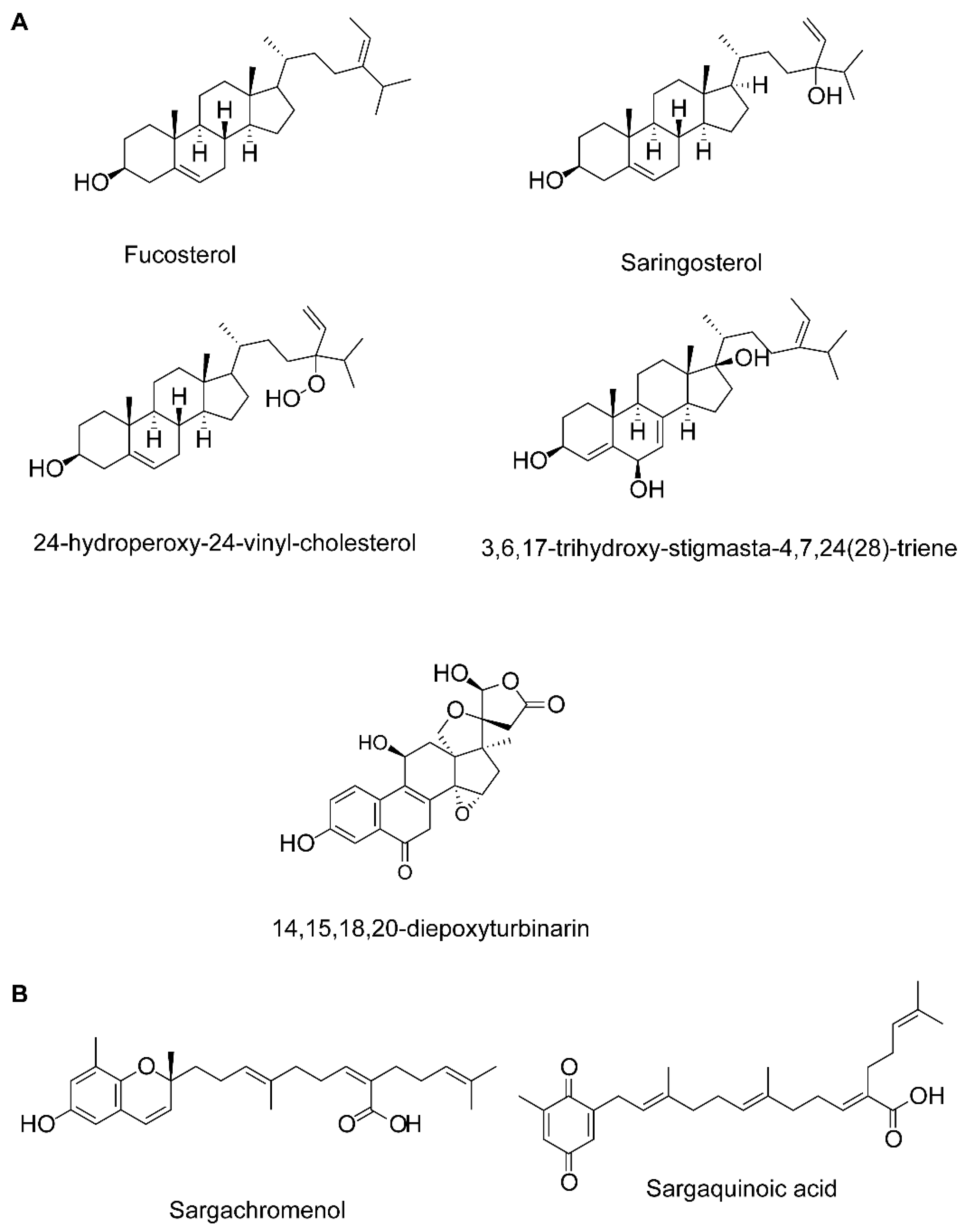
Fucosterol, 3,6,17-trihydroxy-stigmasta-4,7,24(28)-triene and 14,15,18,20-diepoxyturbinarin (sterols) |
Pelvetia siliquosa |
A seven day-dose regimen at 30 mg/kg/day before carbon tetrachloride (CCl4) administration |
Rat model |
Antioxidation |
n.d. |
↑SOD, CAT, and GPx |
[70] |
|
|
Fucosterol |
Eisenia bicyclis, brown alga |
25, 50, 100, 200, and 400 μM |
RAW 264.7 murine macrophages (t-BHP stimulated) |
Protects against oxidative stress |
n.d. |
↓ROS generation |
[71] |
|
|
Fucosterol |
Ecklonia stolonifera and Eisenia bicyclis; Brown algae |
25, 50, and 100 μM |
tert-Butyl hydroperoxide- and tacrine-induced HepG2cell injury model |
Antioxidation |
n.d. |
↓ROS generation ↑GSH level |
[72] |
|
|
Fucosterol |
Sargassum Binderi; brown alga |
3.125, 6.25, 12.5, 25, 50, and 100 μg /mL |
Particulate matter-induced injury and inflammation in A549 human lung epithelial cells |
Attenuates oxidative stress |
↓ROS level ↑SOD, CAT, and HO-1 in the cytosol, and NRF2 in the nucleus |
[73]. |
||
|
Glycoprotein |
U. pinnatifida |
SOD activity and Xox activity at a concentration of 5 mg/mL and 1 mg/mL, respectively |
In vitro enzyme assay |
↑SOD and↓Xox |
[74] |
|||
|
Sulfated oligosaccharides |
Ulva lactuca and Enteromorpha prolifera; green algae |
150 mg/kg·day |
Aging model (male senescence-accelerated prone (SAMP8) and male senescence resistant (SAMR1) mice) |
Antioxidantion |
n.d. |
↑GSH, SOD, CAT, telomerase levels, ↑Total antioxidant capacity, ↓MDA and AGEPs |
[75] |
|
|
Anti-inflammatory activity |
Fucoxanthin |
5, 10, and 50 μM |
Aβ42-induced BV2 microglia cells |
Anti-inflammation |
MAPK pathway |
↓iNOS, COX-2 ↓TNF-α, IL-6, IL-1β, PGE2 ↓JNK, ERK, and p38 MAPK phosphorylation |
[36] |
|
|
Fucoxanthin |
- |
LPS-activated BV-2 microglia |
Anti-inflammation and antioxidation |
Akt/NF-κB and MAPKs/AP-1 pathways; PKA/CREB pathway |
↓iNOS, COX-2, ↓TNF-α, IL-6, PGE2, NO, ROS ↓IL-6, TNF-α, iNOS, and COX-2 mRNA expression ↓Akt, NF-κB, ERK, p38 MAPK and AP-1 phosphorylation ↑Nrf2, HO-1 ↑PKA, CREB ↑BDNF |
[76] |
||
|
Fucosterol |
E. bicyclis; brown alga |
5–20 μM for NO |
RAW 264.7 murine macrophages (t-BHP 200 μM, LPS-1μM stimulated) |
↓Inflammatory response |
↓NF-κB pathway |
↓NO production ↓iNOS and COX-2 |
[71] |
|
|
Fucosterol |
U. pinnatifida |
10, 25, or 50 μM |
LPS-induced RAW 264.7 macrophages and THP-1 human monocyte cell line |
↓Inflammatory response |
↓NF-κB pathway |
↓iNOS, TNF-α, and IL-6 ↓DNA binding ↓phosphorylation of NF-κB, MKK3/6 and MK2 |
[77] |
|
|
Fucosterol |
Hizikia fusiformis |
1–10 μM |
CoCl2 induced hypoxia in keratinocytes |
↓Inflammatory response |
n.d. |
↓IL-6, IL-1β and TNF-α ↓pPI3K and pAkt and HIF1-α accumulation |
[78] |
|
|
Fucosterol |
Panida. australis |
0.004,0.2, and 10 μM |
LPS or Aβ-induced BV2 (microglial) cells |
Protects against LPS or Aβ-mediated neuroinflammation |
n.d. |
↓IL-6, IL-1β, TNF-α, NO, and PGE2 |
[79] |
|
|
Fucosterol |
S. Binderi; brown alga |
3.125, 6.25, 12.5, 25, 50, 100 μg/mL |
Particulate matter-induced injury and inflammation in A549 human lung epithelial cells |
↓Inflammatory response |
n.d. |
↓COX-2, PGE2, TNF-α and IL-6 |
[73] |
|
|
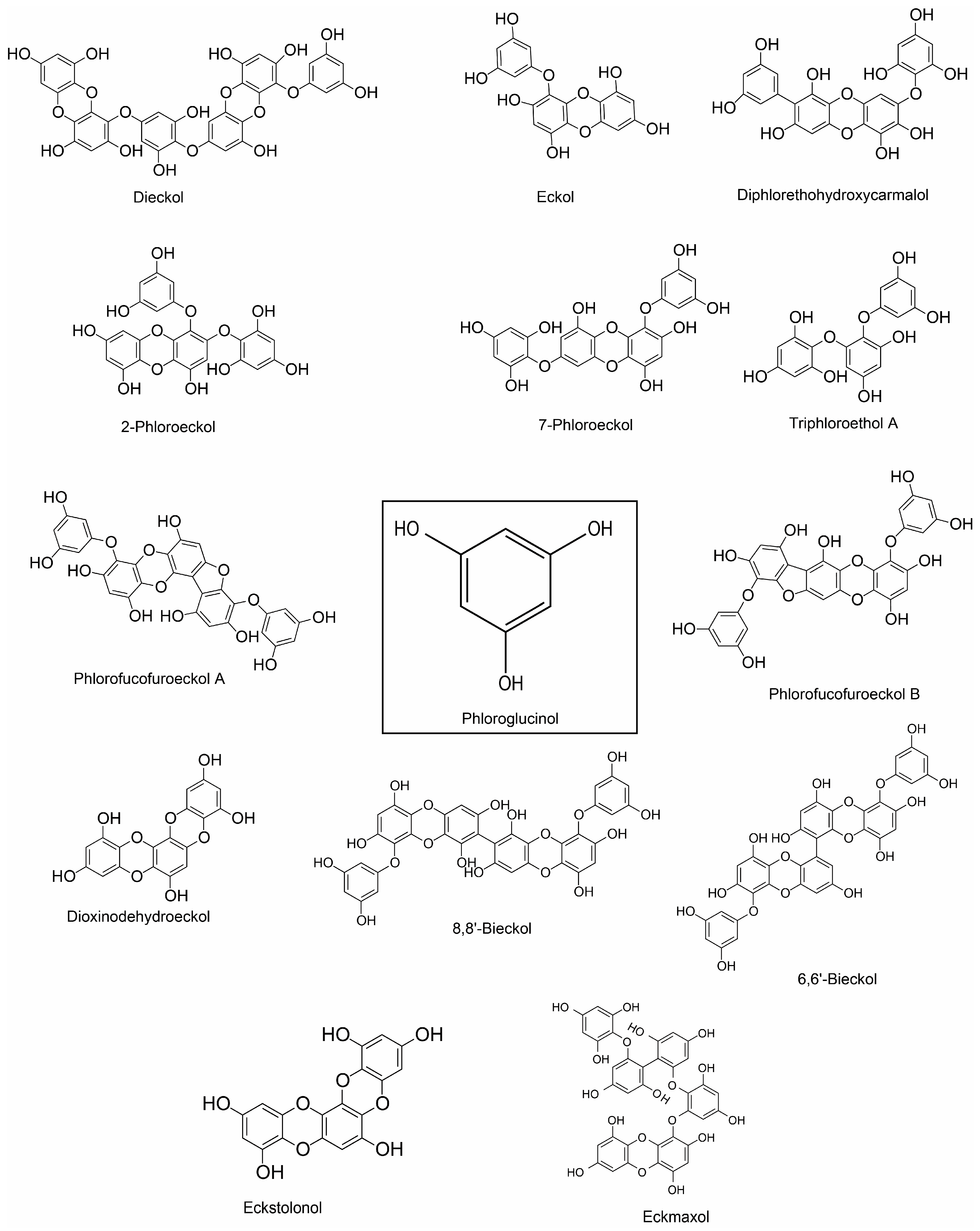
Dieckol (phlorotannin) |
E. cava |
50–300 µg/mL |
LPS-stimulated murine BV2 microglia |
Anti-inflammation and antioxidation |
p-38 MAPK/ NF-κB pathway |
↓NO and PGE2; ↓iNOS and COX-2; ↓IL-1β and TNF-α; ↓ROS |
[80] |
|
|
Phloroglucinol, eckol, dieckol, 7-phloroeckol, phlorofucofuroeckol A and dioxinodehydroeckol (phlorotannin) |
E. bicyclis; brown alga |
5–20 μM for NO |
LPS-stimulated RAW 264.7 murine macrophages |
↓Inflammatory response |
↓NF-κB pathway |
↓NO production |
[71] |
|
|
Phlorofucofuroeckol A |
E. stolonifera |
20 μM |
LPS-activated BV2 and primary microglial cells |
Anti-inflammation |
NF-κB, JNKs, p38 MAPK, and Akt pathways |
↓NO and PGE2; ↓iNOS and COX-2; ↓IL-1β, IL-6 and TNF-α; ↓NF-κB activation and IκB-α degradation ↓JNK, p38, and Akt |
[81] |
|
|
Phlorofucofuroeckol B (phlorotannin) |
E. stolonifera |
10–40 µM |
LPS-stimulated murine BV2 microglia |
Anti-inflammation |
IκB-α/NF-κB and Akt/ERK/JNK pathways |
↓TNF-α, IL-1β and IL-6; ↓COX-2 and iNOS ↓NF-κB activation and IκB-α degradation ↓Akt, ERK, and JNK phosphorylation |
[82] |
|
|
8,8’-bieckol (phlorotannin) |
E. cava |
LPS-stimulated primary macrophages and RAW 264.7 macrophages & LPS-induced septic mice |
Anti-inflammation; Protects mice from endotoxin shock |
NF-κB pathway |
↓NO and PGE2; ↓iNOS mRNA and protein expression; ↓IL-6; ↓Transactivation of NF-κB and nuclear translocation of the NF-κB p65 subunit ↓ROS |
[83] |
||
|
6,6′-bieckol (phlorotannin) |
E.stolonifera |
LPS-stimulated BV2 and murine primary microglial cells |
Anti-inflammation |
IκB-α/NF-κB and JNK/p38 MAPK/Akt pathways |
↓COX-2 and iNOS; ↓NO and PGE2, ↓IL-6 ↓Transactivation of NF-κB and nuclear translocation of the NF-κB p65 subunit ↓Akt, JNK and p38 MAPK phosphorylation |
[84] |
||
|
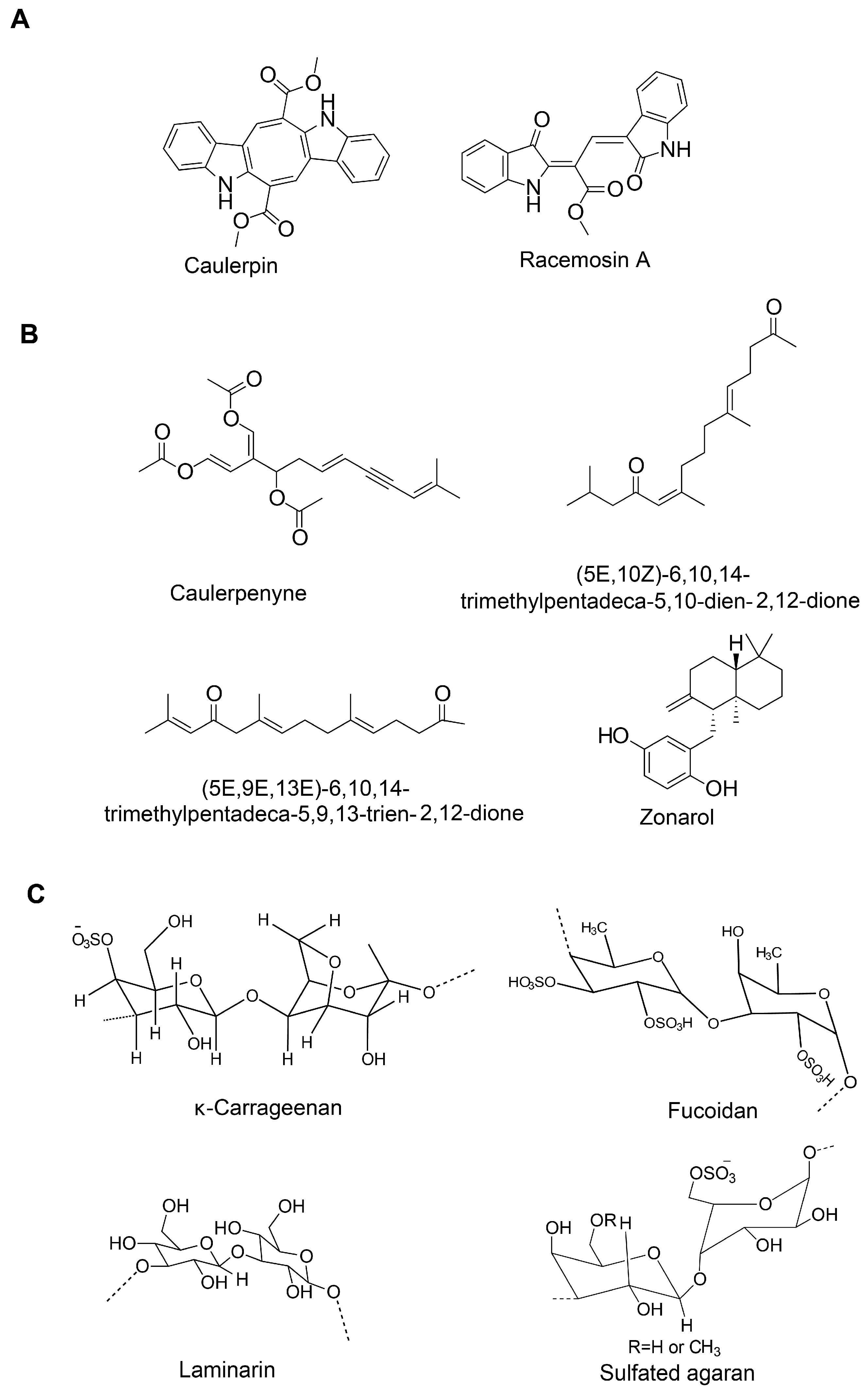
Fucoidan (sulfated polysaccharide) |
Brown seaweed |
25, 50, and 100 µg/mL |
LPS-stimulated murine BV2 microglia |
Anti-inflammation |
NF-κB and JNK/p38 MAPK/Akt pathways |
↓NO and PGE2; ↓COX-2, iNOS and MCP-1; ↓TNF-α and IL-1β; ↓NF-κB activation; ↓Akt, ERK, p38 MAPK and JNK phosphorylation |
[85] |
|
|
Fucoidan |
- |
125 µg/mL |
LPS-activated primary microglia |
Anti-inflammation |
n.d. |
↓TNF-α and ROS |
[86] |
|
|
κ-carrageenan oligosaccharides and desulfated derivatives |
Red algae |
LPS-activated microglia |
Anti-inflammation |
n.d. |
↓TNF-α |
[87] |
||
|
Sulfated oligosaccharides |
U. lactuca and E. prolifera; green algae |
150 mg/kg·day |
Aging model (male senescence-accelerated prone (SAMP8) and male senescence resistant (SAMR1) mice) |
↓Inflammatory response |
n.d. |
↓IFN-γ, TNF-α, and IL-6 |
[75] |
|
|
Alginate-derived oligosaccharide |
Brown algae |
50–500 µg/mL |
LPS/Aβ-stimulated BV2 microglia |
Anti-inflammation |
TLR4/NF-κB signaling pathway |
↓NO and PGE2; ↓COX-2 and iNOS; ↓TNF-α, IL-6 and IL-12; ↓TLR4; ↑NF-κB/p65 subunit translocation |
[88] |
|
|
Seleno-polymannuronate |
Brown algae |
0.8 mg/mL |
LPS-activated primary microglia and astrocytes; mouse model of acute inflammation |
Anti-inflammation |
NF-κB and MAPK signaling |
↓NO and PGE2; ↓COX-2 and iNOS; ↓TNF-α, IL-1β and IL-6; ↑IκB-α, p65, p38, ERK and JNK phosphorylation |
[89] |
|
|
Sargachromenol (plastoquinone) |
Sargassum micracanthum |
30.2 μM (IC50) |
LPS-stimulated RAW 264.7 macrophages |
Anti-inflammation |
NF-κB signaling |
↓NO and PGE2; ↓COX-2 and iNOS; ↑IκB-α |
[90] |
|
|
Sargaquinoic acid (plastoquinone) |
Sargassum siliquastrum |
LPS-stimulated RAW 264.7 macrophages |
Anti-inflammation |
NF-κB signaling |
↓NO; ↓iNOS; ↑IκB-α; ↓nuclear translocation of NF-κB; ↓JNK1/2 MAPK |
[91] |
||
|
Floridoside (glycerol glycosides) |
Laurencia undulate; red alga |
50 μM |
LPS-stimulated murine BV2 microglia |
Anti-inflammation |
MAPK Signaling |
↓NO, ROS; ↓iNOS and COX-2; ↓p38 MAPK and ERK phosphorylation |
[92] |
|
|
Glycoprotein |
U. pinnatifida |
COX-1 and COX-2 inhibition with IC50 values of 53.03 ± 1.03 μg/mL and 193.35 ± 3.08 μg/mL, respectively |
LPS-stimulated RAW 264.7 macrophages |
Anti-inflammation |
n.d. |
↓COX-1 and COX-2 ↓NO |
[74] |
|
|
Caulerpin (bisindole alkaloid) |
Caulerpa racemosa |
100 µM/kg body wt |
Capsaicin-induced ear edema and carrageenan-induced peritonitis |
Inhibition of nociception |
n.d. |
n.d. |
[93] |
|
|
Caulerpenyne (sesquiterpene) |
C. prolifera and C. racemosa |
5.1 μM |
Lipoxygenase (LOX) enzyme activity assay |
Inhibitory activity against LOX |
- |
Un-competitive type of inhibition |
[94] |
|
|
Aquamin (multi-mineral complex) |
Lithothamnion corallioides; red alga |
LPS-stimulated, glial-enriched primary cultures of rat cortex |
Anti-inflammation |
n.d. |
↓TNF-α and IL-1β |
[95] |
||
|
Anticholinesterase activity |
Fucosterol and 24-hydroperoxy 24-vinylcholesterol |
E. stolonifera |
IC50 values of 421.72 ± 1.43, 176.46 ± 2.51 µM, respectively |
In vitro enzymatic assay |
↓BChE activity |
- |
Selective inhibition of BChE |
[96] |
|
Fucosterol |
Panida australis |
inhibition against AChE (10.99–20.71%) and BChE (4.53–17.53%) with concentrations ≤ 56 μM, |
In vitro enzymatic assay |
↓AChE and BChE activities |
- |
Nonselective cholinesterase inhibition |
[79] |
|
|
Fucosterol |
Sargassum horridum |
- |
In vitro enzymatic assay |
↓AChE activity |
- |
Non-competitive inhibition |
[97] |
|
|
Fucoxanthin |
- |
IC50 value 1.97 mM |
In vitro BChE activity assay |
↓BChE activity |
Mixed inhibition type |
[98]. |
||
|
Fucoxanthin |
Brown seaweed |
IC50 value of 81.2 μM |
In vitro AChE activity assay; Molecular docking analysis |
↓AChE activity |
Fucoxanthin likely interacts with the peripheral anionic site within AChE |
Non-competitive manner |
[99] |
|
|
α-Bisabolol |
Padina gymnospora |
IC50 value < 10 μg/mL |
In vitro enzymatic assay |
↓AChE and BChE activity |
- |
- |
[100] |
|
|
Glycoprotein |
U. pinnatifida |
AChE and BChE inhibitory activities with IC50 values of 63.56 ± 1.86 and 99.03 ± 4.64, respectively |
In vitro enzymatic assay |
↓AChE and BChE activity |
- |
- |
[74] |
|
|
Phloroglucinol, |
Ecklonia maxima; Brown alga |
IC50 value: 76.70 to 579.32 μM |
In vitro AChE activity assay |
↓AChE activity |
- |
- |
[101] |
|
|
Dieckol and phlorofucofuroeckol |
E. cava |
Ethanol-intoxicated memory impairment in mice |
↓AChE activity |
n.d. |
↑Acetylcholine |
[102] |
||
|
Sargaquinoic acid and sargachromenol (plastoquinones) |
Sargassum sagamianum |
IC50 value for anti-AChE: 23.2 and 32.7 μM, respectively; IC50 value for anti-BChE of sargaquinoic acid 26 nm |
In vitro ChE activity assay |
Sargaquinoic acid shows potent inhibitory activity against BuChE and moderate inhibitory activity against AChE |
-. |
- |
[103] |
|
|
(5E,10Z)-6,10,14-trimethylpentadeca-5,10-dien-2,12-dione and (5E,9E,13E)-6,10,14-trimethylpentadeca-5,9,13-trien-2,12-dione (Sesquiterpenes) |
S. sagamianum |
IC50 values of 65.0 and 48.0, and 34.0 and 23.0 μM, respectively |
In vitro ChE activity assay |
Moderate inhibitory activity against AChE and BuChE |
- |
- |
[104] |
|
|
Anti-amyloidogenic and aggregation inhibition activity |
Fucoxanthin |
E. stolonifera and U. pinnatifida |
↓β-secretase activity; Binding energy (-7.0 kcal/mol) |
- |
mixed-type inhibition |
[105] |
||
|
Fucoxanthin |
- |
0.1–30 μM |
Suppresses the formation of Aβ1-42 fibrils and Aβ1–42 oligomers, and inhibits Aβ aggregation |
- |
- |
[106] |
||
|
Fucoxanthin |
- |
2 μM |
ThT assay |
Inhibits Aβ1-42 fibril and aggregate formation |
- |
- |
[107] |
|
|
Fucosterol |
E. stolonifera and U. pinnatifida |
10–100 μM (IC50 value of 64.12 ± 1.0 μM) |
In vitro enzyme assay; In silico analysis |
↓β-secretase activity; Binding energy (−10.1 kcal/mol) |
- |
Noncompetitive inhibition |
[105] |
|
|
α-Bisabolol |
Padina gymnospora |
5 μg/mL |
Thioflavin T (ThT), Confocal laser scanning microscopy (CLSM) analysis, Transmission electron microscopy (TEM), Fourier transform infrared (FTIR) spectroscopic analysis and molecular dynamics simulation |
Prevents oligomers formation as well as disaggregates the matured fibrils |
- |
- |
[108] |
|
|
Glycoprotein |
U. pinnatifida |
IC50 values of 73.35 ± 2.54 μg/mL |
In vitro enzymatic assay |
↓BACE1 activity |
- |
- |
[74] |
|
|
Cholesterol homeostasis and Aβ clearance activity |
Fucosterol |
- |
100 and 200 μM (HEK293 cell cultures); 100 or 200 μM (macrophages and HepG2, H4IIE, and Caco2 cells) |
HEK293 cell cultures (Reporter system); THP-1-derived macrophages; Caco-2 cells HepG2 cells |
Reverses cholesterol transport. No accumulation of triglyceride in HepG2 |
n.d. |
Dual-LXR agonist (LXR-α and LXR-β) ↑ABCA1, ABCG1, and ApoE; ↑Intestinal NPC1L1 and ABCA1; ↑Insig-2a, that delays nuclear translocation of SREBP-1c |
[109] |
|
Saringosterol |
Sargassum fusiforme |
30 μM |
Luciferase reporter assay system; HEK293T, THP-1 monocytes, HepG2, RAW264.7, THP-1 macrophages and Caco-2 cells |
n.d. |
n.d. |
Selective LXRβ agonist; ↑ABCA1, ABCG1, and SREBP-1c |
[110] |
|
|
Alginate-derived oligosaccharide |
Marine brown algae |
BV2 microglial cells |
Microglial phagocytosis of Aβ |
Toll-like receptor signaling |
↑TLR4 |
[88]. |
||
|
Monoamine oxidase inhibition and affinity to dopaminergic receptors |
Phlorofucofuroeckol-A and dieckol (phlorotannin) |
- |
In vitro enzyme assay and functional assay for GPCR screening; Docking analysis |
↓hMAO activity; D3R and D4R stimulation |
- |
- |
[111]. |
|
|
Antiaging |
Sulfated oligosaccharides |
U. lactuca and E. prolifera; green algae |
150 mg/kg/day |
Aging model (male senescence-accelerated prone (SAMP8) and male senescence resistant (SAMR1) mice) |
Antioxidant and anti-inflammation |
n.d. |
↑GSH, SOD, CAT, telomerase levels, ↑Total antioxidant capacity, ↓MDA and AGEPs ↓IFN-γ, TNF-α, and IL-6 ↑BDNF and ChAT; ↑Sirt1, ↑p53 and FOXO1 |
[75] |
|
Fucosterol |
Hizikia fusiformis |
50 µg/mL |
Culture model of C. elegans |
Extends lifespan |
↑Antioxidant mechanism |
n.d. |
[112] |
n.d.: not defined; -: information not available.
4. Neuropharmacological Potentials of Marine Algae and Their Metabolites: Evidence from In Vivo Studies
5. Conclusion and Future perspectives
References
- Chi, H.; Chang, H.Y.; Sang, T.K. Neuronal Cell Death Mechanisms in Major Neurodegenerative Diseases. Int. J. Mol. Sci. 2018, 19, 3082.
- Vasili, E.; Dominguez-Meijide, A.; Outeiro, T.F. Spreading of α-Synuclein and Tau: A Systematic Comparison of the Mechanisms Involved. Front. Mol. Neurosci. 2019, 12, 107.
- Zhong, X.; Wang, J.; Carlsson, C.; Okonkwo, O.; Zetterberg, H.; Li, L. A Strategy for Discovery and Verification of Candidate Biomarkers in Cerebrospinal Fluid of Preclinical Alzheimer’s Disease. Front. Mol. Neurosci. 2018, 11, 483.
- Ganguly, G.; Chakrabarti, S.; Chatterjee, U.; Saso, L. Proteinopathy, oxidative stress and mitochondrial dysfunction: Cross talk in Alzheimer’s disease and Parkinson’s disease. Drug Des. Dev. Ther. 2017, 11, 797–810.
- Verri, M.; Pastoris, O.; Dossena, M.; Aquilani, R.; Guerriero, F.; Cuzzoni, G.; Venturini, L.; Ricevuti, G.; Bongiorno, A.I. Mitochondrial Alterations, Oxidative Stress and Neuroinflammation in Alzheimer’s Disease. Int. J. Immunopathol. Pharmacol. 2012, 25, 345–353.
- Jayaraj, R.L.; Azimullah, S.; Beiram, R.; Jalal, F.Y.; Rosenberg, G.A. Neuroinflammation: Friend and foe for ischemic stroke. J. Neuroinflamm. 2019, 16, 142.
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82.
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618.
- Leon, R.; Garcia, A.G.; Marco-Contelles, J. Recent advances in the multitarget-directed ligands approach for the treatment of Alzheimer’s disease. Med. Res. Rev. 2013, 33, 139–189.
- Angeloni, C.; Vauzour, D. Natural Products and Neuroprotection. Int. J. Mol. Sci. 2019, 20, 5570.
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An update. Curr. Neuropharmacol. 2019, 17, 247–267.
- Kim, J.; Lee, H.J.; Lee, K.W. Naturally occurring phytochemicals for the prevention of Alzheimer’s disease. J. Neurochem. 2010, 112, 1415–1430.
- Turner, R.S.; Thomas, R.G.; Craft, S.; van Dyck, C.H.; Mintzer, J.; Reynolds, B.A.; Brewer, J.B.; Rissman, R.A.; Raman, R.; Aisen, P.S. A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology 2015, 85, 1383–1391.
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharifi-Rad, M.; Anil Kumar, N.V.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545.
- Leandro, A.; Pereira, L.; Gonçalves, A.M.M. Diverse Applications of Marine Macroalgae. Mar. Drugs 2020, 18, 17.
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982.
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597.
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828.
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules 2019, 24, 4182.
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266.
- Alghazwi, M.; Kan, Y.Q.; Zhang, W.; Gai, W.P.; Garson, M.J.; Smid, S. Neuroprotective activities of natural products from marine macroalgae during 1999–2015. J. Appl. Phycol. 2016, 28, 3599–3616.
- Barbalace, M.C.; Malaguti, M.; Giusti, L.; Lucacchini, A.; Hrelia, S.; Angeloni, C. Anti-Inflammatory Activities of Marine Algae in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3061.
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive compounds from macroalgae in the new millennium: Implications for neurodegenerative diseases. Mar. Drugs 2014, 12, 4934–4972.
- Cornish, M.L.; Critchley, A.T.; Mouritsen, O.G. Consumption of seaweeds and the human brain. J. Appl. Phycol. 2017, 29, 2377–2398.
- Rengasamy, K.R.R.; Mahomoodally, M.F.; Aumeeruddy, M.Z.; Zengin, G.; Xiao, J.; Kim, D.H. Bioactive compounds in seaweeds: An overview of their biological properties and safety. Food Chem. Toxicol. 2020, 135, 111013.
- Rajeswari, R.; Jeyaprakash, K. Biopotential effects of seaweeds for neurological disorders mini review. J. Pharm. Pharm. Sci. 2017, 6, 427–436.
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6.
- Caltagirone, C.; Ferrannini, L.; Marchionni, N.; Nappi, G.; Scapagnini, G.; Trabucchi, M. The potential protective effect of tramiprosate (homotaurine) against Alzheimer’s disease: A review. Aging Clin. Exp. Res. 2012, 24, 580–587.
- Schepers, M.; Martens, N.; Tiane, A.; Vanbrabant, K.; Liu, H.B.; Lütjohann, D.; Mulder, M.; Vanmierlo, T. Edible seaweed-derived constituents: An undisclosed source of neuroprotective compounds. Neural Regen. Res. 2020, 15, 790–795.
- Williams, L.L. Marine algae as a source of prevention and relief in those with depression and dementia. World J. Pharm. Pharm. Sci. 2017, 6, 26–38.
- Kim, S.K.; Pangestuti, R. Biological activities and potential health benefits of fucoxanthin derived from marine brown algae. Adv. Food Nutr. Res. 2011, 64, 111–128.
- Syed, Y.Y. Sodium Oligomannate: First Approval. Drugs 2020, 80, 441–444.
- Pangestuti, R.; Kim, S.K. Neuroprotective effects of marine algae. Mar. Drugs 2011, 9, 803–818.
- Huang, C.; Zhang, Z.; Cui, W. Marine-Derived Natural Compounds for the Treatment of Parkinson’s Disease. Mar. Drugs 2019, 17, 221.
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609.
- Pangestuti, R.; Vo, T.S.; Ngo, D.H.; Kim, S.K. Fucoxanthin Ameliorates Inflammation and Oxidative Reponses in Microglia. J. Agric. Food Chem. 2013, 61, 3876–3883.
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581.
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583.
- Uddin, M.S.; Kabir, M.T.; Mamun, A.A.; Barreto, G.E.; Rashid, M.; Perveen, A.; Ashraf, G.M. Pharmacological approaches to mitigate neuroinflammation in Alzheimer’s disease. Int. Immunopharmacol. 2020, 84, 106479.
- Hannan, M.A.; Dash, R.; Sohag, A.A.M.; Haque, M.N.; Moon, I.S. Neuroprotection against oxidative stress: Phytochemicals targeting TrkB signaling and the Nrf2-ARE antioxidant system. Front. Mol. Neurosci. 2020, 13, 116.
- Sivandzade, F.; Prasad, S.; Bhalerao, A.; Cucullo, L. NRF2 and NF-κB interplay in cerebrovascular and neurodegenerative disorders: Molecular mechanisms and possible therapeutic approaches. Redox Biol. 2019, 21, 101059.
- Castelli, V.; Benedetti, E.; Antonosante, A.; Catanesi, M.; Pitari, G.; Ippoliti, R.; Cimini, A.; d’Angelo, M. Neuronal Cells Rearrangement During Aging and Neurodegenerative Disease: Metabolism, Oxidative Stress and Organelles Dynamic. Front. Mol. Neurosci. 2019, 12, 132.
- Kandlur, A.; Satyamoorthy, K.; Gangadharan, G. Oxidative Stress in Cognitive and Epigenetic Aging: A Retrospective Glance. Front. Mol. Neurosci. 2020, 13, 41.
- Chaudhari, N.; Talwar, P.; Parimisetty, A.; Lefebvre d’Hellencourt, C.; Ravanan, P. A Molecular Web: Endoplasmic Reticulum Stress, Inflammation, and Oxidative Stress. Front. Mol. Neurosci. 2014, 8, 213.
- Gerakis, Y.; Hetz, C. Emerging roles of ER stress in the etiology and pathogenesis of Alzheimer’s disease. FEBS J. 2018, 285, 995–1011.
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 2019, 10, 1008.
- Yanuck, S.F. Microglial Phagocytosis of Neurons: Diminishing Neuronal Loss in Traumatic, Infectious, Inflammatory, and Autoimmune CNS Disorders. Front. Psychiatry 2019, 10, 712.
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263.
- Dash, R.; Mitra, S.; Ali, M.C.; Oktaviani, D.F.O.; Hannan, M.A.; Choi, S.M.; Moon, I.S. Phytosterols: Targeting Neuroinflammation in Neurodegeneration. Curr. Pharm. Des. 2020, 26, 1–23.
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid. Med. Cell. Longev. 2017, 2017, 2525967.
- Mouzat, K.; Chudinova, A.; Polge, A.; Kantar, J.; Camu, W.; Raoul, C.; Lumbroso, S. Regulation of Brain Cholesterol: What Role Do Liver X Receptors Play in Neurodegenerative Diseases? Int. J. Mol. Sci. 2019, 20, 3858.
- Ito, A.; Hong, C.; Rong, X.; Zhu, X.; Tarling, E.J.; Hedde, P.N.; Gratton, E.; Parks, J.; Tontonoz, P. LXRs link metabolism to inflammation through Abca1-dependent regulation of membrane composition and TLR signaling. eLife 2015, 4, e08009.
- Xu, P.; Li, D.; Tang, X.; Bao, X.; Huang, J.; Tang, Y.; Yang, Y.; Xu, H.; Fan, X. LXR agonists: New potential therapeutic drug for neurodegenerative diseases. Mol. Neurobiol. 2013, 48, 715–728.
- Dai, Y.B.; Tan, X.J.; Wu, W.F.; Warner, M.; Gustafsson, J.A. Liver X receptor beta protects dopaminergic neurons in a mouse model of Parkinson disease. Proc. Natl. Acad. Sci. USA 2012, 109, 13112–13117.
- Futter, M.; Diekmann, H.; Schoenmakers, E.; Sadiq, O.; Chatterjee, K.; Rubinsztein, D.C. Wild-type but not mutant huntingtin modulates the transcriptional activity of liver X receptors. J. Med. Genet. 2009, 46, 438–446.
- Wolf, A.; Bauer, B.; Hartz, A.M. ABC Transporters and the Alzheimer’s Disease Enigma. Front. Psychiatry 2012, 3, 54.
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic stroke. Nat. Rev. Dis. Primers 2019, 5, 70.
- Soares, R.O.S.; Losada, D.M.; Jordani, M.C.; Évora, P.; Castroesilva, O. Ischemia/Reperfusion Injury Revisited: An Overview of the Latest Pharmacological Strategies. Int. J. Mol. Sci. 2019, 20, 5034.
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting Oxidative Stress and Inflammation to Prevent Ischemia-Reperfusion Injury. Front. Mol. Neurosci. 2020, 13, 28.
- Tschoe, C.; Bushnell, C.D.; Duncan, P.W.; Alexander-Miller, M.A.; Wolfe, S.Q. Neuroinflammation after Intracerebral Hemorrhage and Potential Therapeutic Targets. J. Stroke 2020, 22, 29–46.
- Liu, F.; Lu, J.; Manaenko, A.; Tang, J.; Hu, Q. Mitochondria in Ischemic Stroke: New Insight and Implications. Aging Dis. 2018, 9, 924–937.
- Yang, J.L.; Mukda, S.; Chen, S.D. Diverse roles of mitochondria in ischemic stroke. Redox Biol. 2018, 16, 263–275.
- Gadani, S.P.; Walsh, J.T.; Lukens, J.R.; Kipnis, J. Dealing with Danger in the CNS: The Response of the Immune System to Injury. Neuron 2015, 87, 47–62.
- Mracsko, E.; Veltkamp, R. Neuroinflammation after intracerebral hemorrhage. Front. Cell. Neurosci. 2014, 8, 388.
- Malekahmadi, M.; Moradi Moghaddam, O.; Islam, S.M.S.; Tanha, K.; Nematy, M.; Pahlavani, N.; Firouzi, S.; Zali, M.R.; Norouzy, A. Evaluation of the effects of pycnogenol (French maritime pine bark extract) supplementation on inflammatory biomarkers and nutritional and clinical status in traumatic brain injury patients in an intensive care unit: A randomized clinical trial protocol. Trials 2020, 21, 162.
- Beauchamp, K.; Mutlak, H.; Smith, W.R.; Shohami, E.; Stahel, P.F. Pharmacology of traumatic brain injury: Where is the “golden bullet”? Mol. Med. 2008, 14, 731–740.
- Khatri, N.; Thakur, M.; Pareek, V.; Kumar, S.; Sharma, S.; Datusalia, A.K.J.C.; Targets, N.D.-D. Oxidative stress: Major threat in traumatic brain injury. CNS Neurol. Disord. Drug Targets 2018, 17, 689–695.
- Rodriguez-Rodriguez, A.; Jose Egea-Guerrero, J.; Murillo-Cabezas, F.; Carrillo-Vico, A. Oxidative stress in traumatic brain injury. Curr. Med. Chem. 2014, 21, 1201–1211.
- Heo, S.J.; Ko, S.C.; Kang, S.M.; Kang, H.S.; Kim, J.P.; Kim, S.H.; Lee, K.W.; Cho, M.G.; Jeon, Y.J. Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage. Eur. Food Res. Technol. 2008, 228, 145–151.
- Lee, S.; Lee, Y.S.; Jung, S.H.; Kang, S.S.; Shin, K.H. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722.
- Jung, H.A.; Jin, S.E.; Ahn, B.R.; Lee, C.M.; Choi, J.S. Anti-inflammatory activity of edible brown alga Eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem. Toxicol. 2013, 59, 199–206.
- Choi, J.S.; Han, Y.R.; Byeon, J.S.; Choung, S.Y.; Sohn, H.S.; Jung, H.A. Protective effect of fucosterol isolated from the edible brown algae, Ecklonia stolonifera and Eisenia bicyclis, on tert-butyl hydroperoxide- and tacrine-induced HepG2 cell injury. J. Pharm. Pharmacol. 2015, 67, 1170–1178.
- Fernando, I.P.S.; Jayawardena, T.U.; Kim, H.S.; Lee, W.W.; Vaas, A.P.J.P.; De Silva, H.I.C.; Abayaweera, G.S.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, D.S.; et al. Beijing urban particulate matter-induced injury and inflammation in human lung epithelial cells and the protective effects of fucosterol from Sargassum binderi (Sonder ex J. Agardh). Environ. Res. 2019, 172, 150–158.
- Rafiquzzaman, S.M.; Kim, E.Y.; Lee, J.M.; Mohibbullah, M.; Alam, M.B.; Soo Moon, I.; Kim, J.M.; Kong, I.S. Anti-Alzheimers and anti-inflammatory activities of a glycoprotein purified from the edible brown alga Undaria pinnatifida. Food Res. Int. 2015, 77, 118–124.
- Liu, X.Y.; Liu, D.; Lin, G.P.; Wu, Y.J.; Gao, L.Y.; Ai, C.; Huang, Y.F.; Wang, M.F.; El-Seedi, H.R.; Chen, X.H.; et al. Anti-ageing and antioxidant effects of sulfate oligosaccharides from green algae Ulva lactuca and Enteromorpha prolifera in SAMP8 mice. Int. J. Biol. Macromol. 2019, 139, 342–351.
- Zhao, D.; Kwon, S.H.; Chun, Y.S.; Gu, M.Y.; Yang, H.O. Anti-Neuroinflammatory Effects of Fucoxanthin via Inhibition of Akt/NF-κB and MAPKs/AP-1 Pathways and Activation of PKA/CREB Pathway in Lipopolysaccharide-Activated BV-2 Microglial Cells. Neurochem. Res. 2017, 42, 667–677.
- Yoo, M.S.; Shin, J.S.; Choi, H.E.; Cho, Y.W.; Bang, M.H.; Baek, N.I.; Lee, K.T. Fucosterol isolated from Undaria pinnatifida inhibits lipopolysaccharide-induced production of nitric oxide and pro-inflammatory cytokines via the inactivation of nuclear factor-kappaB and p38 mitogen-activated protein kinase in RAW264.7 macrophages. Food Chem. 2012, 135, 967–975.
- Sun, Z.; Mohamed, M.A.A.; Park, S.Y.; Yi, T.H. Fucosterol protects cobalt chloride induced inflammation by the inhibition of hypoxia-inducible factor through PI3K/Akt pathway. Int. Immunopharmacol. 2015, 29, 642–647.
- Wong, C.H.; Gan, S.Y.; Tan, S.C.; Gany, S.A.; Ying, T.; Gray, A.I.; Igoli, J.; Chan, E.W.L.; Phang, S.M. Fucosterol inhibits the cholinesterase activities and reduces the release of pro-inflammatory mediators in lipopolysaccharide and amyloid-induced microglial cells. J. Appl. Phycol. 2018, 30, 3261–3270.
- Jung, W.K.; Heo, S.J.; Jeon, Y.J.; Lee, C.M.; Park, Y.M.; Byun, H.G.; Choi, Y.H.; Park, S.G.; Choi, I.W. Inhibitory effects and molecular mechanism of dieckol isolated from marine brown alga on COX-2 and iNOS in microglial cells. J. Agric. Food Chem. 2009, 57, 4439–4446.
- Kim, A.R.; Lee, M.S.; Choi, J.W.; Utsuki, T.; Kim, J.I.; Jang, B.C.; Kim, H.R. Phlorofucofuroeckol A suppresses expression of inducible nitric oxide synthase, cyclooxygenase-2, and pro-inflammatory cytokines via inhibition of nuclear factor-κB, c-Jun NH2-terminal kinases, and Akt in microglial cells. Inflammation 2013, 36, 259–271.
- Yu, D.K.; Lee, B.; Kwon, M.; Yoon, N.; Shin, T.; Kim, N.G.; Choi, J.S.; Kim, H.R. Phlorofucofuroeckol B suppresses inflammatory responses by down-regulating nuclear factor κB activation via Akt, ERK, and JNK in LPS-stimulated microglial cells. Int. Immunopharmacol. 2015, 28, 1068–1075.
- Yang, Y.I.; Jung, S.H.; Lee, K.T.; Choi, J.H. 8,8′-Bieckol, isolated from edible brown algae, exerts its anti-inflammatory effects through inhibition of NF-κB signaling and ROS production in LPS-stimulated macrophages. Int. Immunopharmacol. 2014, 23, 460–468.
- Kim, A.R.; Lee, B.; Joung, E.J.; Gwon, W.G.; Utsuki, T.; Kim, N.G.; Kim, H.R. 6,6′-Bieckol suppresses inflammatory responses by down-regulating nuclear factor-κB activation via Akt, JNK, and p38 MAPK in LPS-stimulated microglial cells. Immunopharmacol. Immunotoxicol. 2016, 38, 244–252.
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.Y.; Kim, G.Y.; Choi, I.W.; Kim, N.D.; Nam, T.J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752.
- Cui, Y.Q.; Jia, Y.J.; Zhang, T.; Zhang, Q.B.; Wang, X.M. Fucoidan Protects against Lipopolysaccharide-Induced Rat Neuronal Damage and Inhibits the Production of Proinflammatory Mediators in Primary Microglia. CNS Neurosci. Ther. 2012, 18, 827–833.
- Yao, Z.A.; Xu, L.; Wu, H.G. Immunomodulatory function of κ-carrageenan oligosaccharides acting on LPS-activated microglial cells. Neurochem. Res. 2014, 39, 333–343.
- Zhou, R.; Shi, X.Y.; Bi, D.C.; Fang, W.S.; Wei, G.B.; Xu, X. Alginate-Derived Oligosaccharide Inhibits Neuroinflammation and Promotes Microglial Phagocytosis of β-Amyloid. Mar. Drugs 2015, 13, 5828–5846.
- Bi, D.; Lai, Q.; Han, Q.; Cai, N.; He, H.; Fang, W.; Yi, J.; Li, X.; Xu, H.; Li, X.; et al. Seleno-polymannuronate attenuates neuroinflammation by suppressing microglial and astrocytic activation. J. Funct. Foods 2018, 51, 113–120.
- Yang, E.J.; Ham, Y.M.; Yang, K.W.; Lee, N.H.; Hyun, C.G. Sargachromenol from Sargassum micracanthum inhibits the lipopolysaccharide-induced production of inflammatory mediators in RAW 264.7 macrophages. Sci. World J. 2013, 2013, 712303.
- Kang, G.J.; Han, S.C.; Yoon, W.J.; Koh, Y.S.; Hyun, J.W.; Kang, H.K.; Youl Cho, J.; Yoo, E.S. Sargaquinoic acid isolated from Sargassum siliquastrum inhibits lipopolysaccharide-induced nitric oxide production in macrophages via modulation of nuclear factor-κB and c-Jun N-terminal kinase pathways. Immunopharm. Immunot. 2013, 35, 80–87.
- Kim, M.; Li, Y.X.; Dewapriya, P.; Ryu, B.; Kim, S.K. Floridoside suppresses pro-inflammatory responses by blocking MAPK signaling in activated microglia. BMB Rep. 2013, 46, 398–403.
- De Souza, E.T.; de Lira, D.P.; de Queiroz, A.C.; da Silva, D.J.C.; de Aquino, A.B.; Mella, E.A.C.; Lorenzo, V.P.; de Miranda, G.E.C.; de Araújo-Júnior, J.X.; de Oliveira Chaves, M.C.; et al. The antinociceptive and anti-inflammatory activities of caulerpin, a bisindole alkaloid isolated from seaweeds of the genus Caulerpa. Mar. Drugs 2009, 7, 689–704.
- Cengiz, S.; Cavas, L.; Yurdakoc, K.; Pohnert, G. The Sesquiterpene Caulerpenyne from Caulerpa spp. is a Lipoxygenase Inhibitor. Mar. Biotechnol. 2011, 13, 321–326.
- Ryan, S.; O’Gorman, D.M.; Nolan, Y.M. Evidence that the marine-derived multi-mineral Aquamin has anti-inflammatory effects on cortical glial-enriched cultures. Phytother. Res. 2011, 25, 765–767.
- Yoon, N.Y.; Chung, H.Y.; Kim, H.R.; Choi, J.S. Acetyl- and butyrylcholinesterase inhibitory activities of sterols and phlorotannins from Ecklonia stolonifera. Fish. Res. 2008, 74, 200–207.
- Castro-Silva, E.S.; Bello, M.; Hernandez-Rodriguez, M.; Correa-Basurto, J.; Murillo-Alvarez, J.I.; Rosales-Hernandez, M.C.; Munoz-Ochoa, M. In vitro and in silico evaluation of fucosterol from Sargassum horridum as potential human acetylcholinesterase inhibitor. J. Biomol. Struct. Dyn. 2019, 37, 3259–3268.
- Kawee-ai, A.; Kuntiya, A.; Kim, S.M. Anticholinesterase and antioxidant activities of fucoxanthin purified from the microalga Phaeodactylum tricornutum. Nat. Prod. Commun. 2013, 8, 1381–1386.
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase in vitro. Mar. Drugs 2016, 14, 67.
- Shanmuganathan, B.; Sheeja Malar, D.; Sathya, S.; Pandima Devi, K. Antiaggregation Potential of Padina gymnospora against the Toxic Alzheimer’s Beta-Amyloid Peptide 25-35 and Cholinesterase Inhibitory Property of Its Bioactive Compounds. PLoS ONE 2015, 10, e0141708.
- Kannan, R.R.R.; Aderogba, M.A.; Ndhlala, A.R.; Stirk, W.A.; Van Staden, J. Acetylcholinesterase inhibitory activity of phlorotannins isolated from the brown alga, Ecklonia maxima (Osbeck) Papenfuss. Food Res. Int. 2013, 54, 1250–1254.
- Myung, C.S.; Shin, H.C.; Bao, H.Y.; Yeo, S.J.; Lee, B.H.; Kang, J.S. Improvement of memory by dieckol and phlorofucofuroeckol in ethanol-treated mice: Possible involvement of the inhibition of acetylcholinesterase. Arch. Pharm. Res. 2005, 28, 691–698.
- Choi, B.W.; Ryu, G.; Park, S.H.; Kim, E.S.; Shin, J.; Roh, S.S.; Shin, H.C.; Lee, B.H. Anticholinesterase activity of plastoquinones from Sargassum sagamianum: Lead compounds for Alzheimer’s disease therapy. Phytother. Res. 2007, 21, 423–426.
- Ryu, G.; Park, S.H.; Kim, E.S.; Choi, B.W.; Ryu, S.Y.; Lee, B.H. Cholinesterase inhibitory activity of two farnesylacetone derivatives from the brown alga Sargassum sagamianum. Arch. Pharm. Res. 2003, 26, 796–799.
- Jung, H.A.; Ali, M.Y.; Choi, R.J.; Jeong, H.O.; Chung, H.Y.; Choi, J.S. Kinetics and molecular docking studies of fucosterol and fucoxanthin, BACE1 inhibitors from brown algae Undaria pinnatifida and Ecklonia stolonifera. Food Chem. Toxicol. 2016, 89, 104–111.
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J. Agric. Food Chem. 2017, 65, 4092–4102.
- Alghazwi, M.; Smid, S.; Musgrave, I.; Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta (Aβ(1-42)) toxicity and aggregation. Neurochem. Int. 2019, 124, 215–224.
- Shanmuganathan, B.; Suryanarayanan, V.; Sathya, S.; Narenkumar, M.; Singh, S.K.; Ruckmani, K.; Pandima Devi, K. Anti-amyloidogenic and anti-apoptotic effect of α-bisabolol against Aβ induced neurotoxicity in PC12 cells. Eur. J. Med. Chem. 2018, 143, 1196–1207.
- Hoang, M.H.; Jia, Y.; Jun, H.J.; Lee, J.H.; Lee, B.Y.; Lee, S.J. Fucosterol Is a Selective Liver X Receptor Modulator That Regulates the Expression of Key Genes in Cholesterol Homeostasis in Macrophages, Hepatocytes, and Intestinal Cells. J. Agric. Food Chem. 2012, 60, 11567–11575.
- Chen, Z.; Liu, J.; Fu, Z.; Ye, C.; Zhang, R.; Song, Y.; Zhang, Y.; Li, H.; Ying, H.; Liu, H. 24(S)-Saringosterol from edible marine seaweed Sargassum fusiforme is a novel selective LXRbeta agonist. J. Agric. Food Chem. 2014, 62, 6130–6137.
- Seong, S.H.; Paudel, P.; Choi, J.W.; Ahn, D.H.; Nam, T.J.; Jung, H.A.; Choi, J.S. Probing multi-target action of phlorotannins as new monoamine oxidase inhibitors and dopaminergic receptor modulators with the potential for treatment of neuronal disorders. Mar. Drugs 2019, 17, 377.
- Oktaviani, D.F.; Bae, Y.S.; Meinita, M.D.N.; Moon, I.S.; Hong, Y.K. An ethanol extract of the brown seaweed Hizikia fusiformis and its active constituent, fucosterol, extend the lifespan of the nematode Caenorhabditis elegans. J. Life Sci. 2019, 29, 1120–1125.
- Luo, D.; Zhang, Q.; Wang, H.; Cui, Y.; Sun, Z.; Yang, J.; Zheng, Y.; Jia, J.; Yu, F.; Wang, X.; et al. Fucoidan protects against dopaminergic neuron death in vivo and in vitro. Eur. J. Pharmacol. 2009, 617, 33–40.
- Gao, Y.; Li, C.; Yin, J.; Shen, J.; Wang, H.; Wu, Y.; Jin, H. Fucoidan, a sulfated polysaccharide from brown algae, improves cognitive impairment induced by infusion of Aβ peptide in rats. Environ. Toxicol. Pharmacol. 2012, 33, 304–311.
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective effects of fucoidan on Aβ25-35 and d-Gal-induced neurotoxicity in PC12 cells and d-Gal-induced cognitive dysfunction in mice. Mar. Drugs 2017, 15, 77.
- Wang, X.; Yi, K.; Zhao, Y. Fucoidan inhibits amyloid-β-induced toxicity in transgenic Caenorhabditis elegans by reducing the accumulation of amyloid-β and decreasing the production of reactive oxygen species. Food Funct. 2018, 9, 552–560.
- Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoo, S.K.; Han, H.J.; Chung, D.H.; Kim, D.O.; Kim, G.H.; Heo, H.J. Fucoidan-rich substances from ecklonia cava improve trimethyltin-induced cognitive dysfunction via down-regulation of amyloid β production/Tau hyperphosphorylation. Mar. Drugs 2019, 17, 591.
- Kim, H.; Ahn, J.H.; Song, M.; Kim, D.W.; Lee, T.K.; Lee, J.C.; Kim, Y.M.; Kim, J.D.; Cho, J.H.; Hwang, I.K.; et al. Pretreated fucoidan confers neuroprotection against transient global cerebral ischemic injury in the gerbil hippocampal CA1 area via reducing of glial cell activation and oxidative stress. Biomed. Pharmacother. 2019, 109, 1718–1727.
- Lee, T.K.; Ahn, J.H.; Park, C.W.; Kim, B.; Park, Y.E.; Lee, J.C.; Park, J.H.; Yang, G.E.; Shin, M.C.; Cho, J.H.; et al. Pre-treatment with laminarin protects hippocampal ca1 pyramidal neurons and attenuates reactive gliosis following transient forebrain ischemia in gerbils. Mar. Drugs 2020, 18, 52.
- Park, J.H.; Ahn, J.H.; Lee, T.K.; Park, C.W.; Kim, B.; Lee, J.C.; Kim, D.W.; Shin, M.C.; Cho, J.H.; Lee, C.H.; et al. Laminarin pretreatment provides neuroprotection against forebrain ischemia/reperfusion injury by reducing oxidative stress and neuroinflammation in aged gerbils. Mar. Drugs 2020, 18, 213.
- Liu, Y.; Geng, L.; Zhang, J.; Wang, J.; Zhang, Q.; Duan, D.; Zhang, Q. Oligo-Porphyran ameliorates neurobehavioral deficits in parkinsonian mice by regulating the PI3K/Akt/Bcl-2 pathway. Mar. Drugs 2018, 16, 82.
- Zhang, Z.; Wang, X.; Pan, Y.; Wang, G.; Mao, G. The degraded polysaccharide from Pyropia haitanensis represses amyloid beta peptide-induced neurotoxicity and memory in vivo. Int. J. Biol. Macromol. 2020, 146, 725–729.
- Hu, L.; Chen, W.; Tian, F.; Yuan, C.; Wang, H.; Yue, H. Neuroprotective role of fucoxanthin against cerebral ischemic/reperfusion injury through activation of Nrf2/HO-1 signaling. Biomed. Pharmacother. 2018, 106, 1484–1489.
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin provides neuroprotection in models of traumatic brain injury via the Nrf2-ARE and Nrf2-autophagy pathways. Sci. Rep. 2017, 7, 46763.
- Oh, J.H.; Choi, J.S.; Nam, T.J. Fucosterol from an edible brown alga Ecklonia stolonifera prevents soluble amyloid beta-induced cognitive dysfunction in aging rats. Mar. Drugs 2018, 16, 368.
- Yang, E.J.; Ahn, S.; Ryu, J.; Choi, M.S.; Choi, S.; Chong, Y.H.; Hyun, J.W.; Chang, M.J.; Kim, H.S. Phloroglucinol Attenuates the Cognitive Deficits of the 5XFAD Mouse Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0135686.
- Yang, Y.; Yang, I.; Cao, M.; Su, Z.Y.; Wu, R.; Guo, Y.; Fang, M.; Kong, A.N. Fucoxanthin elicits epigenetic modifications, nrf2 activation and blocking transformation in mouse skin JB6 P+ Cells. AAPS J. 2018, 20, 32.
- Kim, J.J.; Kang, Y.J.; Shin, S.A.; Bak, D.H.; Lee, J.W.; Lee, K.B.; Yoo, Y.C.; Kim, D.K.; Lee, B.H.; Kim, D.W.; et al. Phlorofucofuroeckol improves glutamate-induced neurotoxicity through modulation of oxidative stress-mediated mitochondrial dysfunction in PC12 cells. PLoS ONE 2016, 11, e0163433.
- Pentón-Rol, G.; Marín-Prida, J.; Pardo-Andreu, G.; Martínez-Sánchez, G.; Acosta-Medina, E.F.; Valdivia-Acosta, A.; Lagumersindez-Denis, N.; Rodríguez-Jiménez, E.; Llópiz-Arzuaga, A.; López-Saura, P.A.; et al. C-Phycocyanin is neuroprotective against global cerebral ischemia/reperfusion injury in gerbils. Brain. Res. Bull. 2011, 86, 42–52.
- Wu, S.; Yue, Y.; Tian, H.; Tao, L.; Wang, Y.; Xiang, J.; Wang, S.; Ding, H. Tramiprosate protects neurons against ischemic stroke by disrupting the interaction between PSD95 and nNOS. Neuropharmacology 2014, 83, 107–117.
- Souza, R.B.; Frota, A.F.; Sousa, R.S.; Cezario, N.A.; Santos, T.B.; Souza, L.M.; Coura, C.O.; Monteiro, V.S.; Cristino Filho, G.; Vasconcelos, S.M.; et al. Neuroprotective effects of sulphated agaran from marine alga Gracilaria cornea in Rat 6-Hydroxydopamine parkinson’s disease model: Behavioural, neurochemical and transcriptional alterations. Basic Clin. Pharmacol. Toxicol. 2017, 120, 159–170.
- Oh, J.H.; Nam, T.J. Hydrophilic glycoproteins of an edible green alga Capsosiphon fulvescens prevent aging-induced spatial memory impairment by suppressing gsk-3beta-mediated er stress in dorsal hippocampus. Mar. Drugs 2019, 17, 168.
- Oh, J.H.; Nam, T.J.; Choi, Y.H. Capsosiphon fulvescens Glycoproteins Enhance Probiotics-Induced Cognitive Improvement in Aged Rats. Nutrients 2020, 12, 837.
- Mohibbullah, M.; Choi, J.S.; Bhuiyan, M.M.H.; Haque, M.N.; Rahman, M.K.; Moon, I.S.; Hong, Y.K. The red alga Gracilariopsis chorda and its active constituent arachidonic acid promote spine dynamics via dendritic filopodia and potentiate functional synaptic plasticity in hippocampal neurons. J. Med. Food 2018, 21, 481–488.
- Nisha, S.A.; Devi, K.P. Gelidiella acerosa protects against Aβ 25-35-induced toxicity and memory impairment in Swiss Albino mice: An in vivo report. Pharm. Biol. 2017, 55, 1423–1435.
- Siddiqui, P.J.A.; Khan, A.; Uddin, N.; Khaliq, S.; Rasheed, M.; Nawaz, S.; Dar, A.; Hanif, M. Sargassum swartzii extracts ameliorate memory functions by neurochemical modulation in a rat model. Food Sci. Biotechnol. 2017, 26, 1055–1062.
- Kim, T.E.; Son, H.J.; Lim, D.W.; Yoon, M.; Lee, J.; Kim, Y.T.; Han, D.; Lee, C.; Um, M.Y. Memory-enhancing effects of Ishige foliacea extract: In vitro and in vivo study. J. Food Biochem. 2020, 44, e13162.
- Choi, J.Y.; Mohibbullah, M.; Park, I.S.; Moon, I.S.; Hong, Y.K. An ethanol extract from the phaeophyte Undaria pinnatifida improves learning and memory impairment and dendritic spine morphology in hippocampal neurons. J. Appl. Phycol. 2018, 30, 129–136.
- El-Baz, F.K.; Aly, H.F.; Ali, G.H. Haematococcus pluvialis modulating effect on neurotransmitters, hormones and oxidative damage-associated with alzheimers disease in experimental rats model. Int. J. Pharm. Pharm. Sci. 2017, 9, 198–206.
- Choi, W.Y.; Kang, D.H.; Heo, S.J.; Lee, H.Y. Enhancement of the neuroprotective effect of fermented Spirulina maxima associated with antioxidant activities by ultrasonic extraction. Appl. Sci. 2018, 8, 2469.
- Kim, J.H.; Lee, N.S.; Jeong, Y.G.; Lee, J.H.; Kim, E.J.; Han, S.Y. Protective efficacy of an Ecklonia cava extract used to treat transient focal ischemia of the rat brain. Anat. Cell. Biol. 2012, 45, 103–113.

