 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Mara Fernandes | + 3654 word(s) | 3654 | 2021-12-21 04:25:30 | | | |
| 2 | Lindsay Dong | Meta information modification | 3654 | 2022-01-20 04:31:06 | | |
Video Upload Options
Non-Hodgkin lymphoma (NHL) is a heterogeneous malignancy with variable patient outcomes. There is still a lack of understanding about the different players involved in lymphomagenesis, and the identification of new diagnostic and prognostic biomarkers is urgent. MicroRNAs and long non-coding RNAs emerged as master regulators of B-cell development, and their deregulation has been associated with the initiation and progression of lymphomagenesis. They can function by acting alone or, as recently proposed, by creating competing endogenous RNA (ceRNA) networks. The study of miRNAs’ and lncRNAs’ deregulation in NHL, either alone or as ceRNAs networks, offers new insights into the molecular mechanisms underlying lymphoma pathogenesis and opens a window of opportunity to identify potential diagnostic and prognostic biomarkers.
1. Introduction
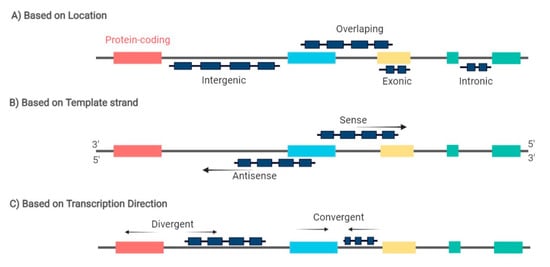
2. MiRNAs and lncRNAs Deregulation in Lymphomagenesis
2.1. The Role of miRNAs in B-Cell Lymphomagenesis
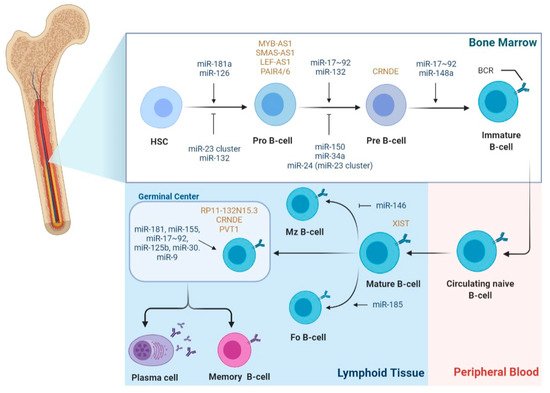
2.2. The Role of LncRNAs in B-Cell Lymphomagenesis
2.3. The Role of LncRNAs as ceRNAs in B-Cell Lymphomagenesis
Karreth et al. demonstrated that lncRNA BRAFP1, which is aberrantly expressed in B-cell lymphomas, acts as a ceRNA with BRAF mRNA, increasing its stability and BRAF levels by sequestering specific BRAF-targeting miRNAs, such as miR-134, miR-543, and miR-653. Consequently, BRAF activates MAPK signaling, resulting in DLBCL cells’ proliferation [34]. In fact, NEAT1 was identified as an MYC-regulated transcript promoting DLBCL cells proliferation and lymphomagenesis by regulating the miR-34b-5p-GLI1 pathway [35]. Interestingly, NEAT1, along with LincRNA-p21, were also identified as p53-dependent DNA damage response machinery in lymphoma and CLL [29].
Another reported upregulated lncRNA in DLBCL is MALAT1, whose ceRNA function is through sponging miR-195, resulting in the activation of the immune checkpoint molecule PD-L1 and consequently promoting cell proliferation, migration, and immune escape. Moreover, MALAT1 can induce CD8+ T cell apoptosis and epithelial–mesenchymal transition (EMT)-like processes by regulating the Ras/ERK signaling pathway [36]. In MCL, the knockdown of MALAT1 resulted in cell-cycle arrest and impaired proliferation due to the upregulation of p21 and p27 by EZH2 [37]. MiR-423-5p was reported to be involved in a ceRNA network with lncRNA FOXP4-AS1 in MCL cells. Mechanistically, FOXP4-AS1 acts as a sponge to miR-423-5p, upregulating the expression of NACC1, which results in MCL cell proliferation, migration, and invasion [38].
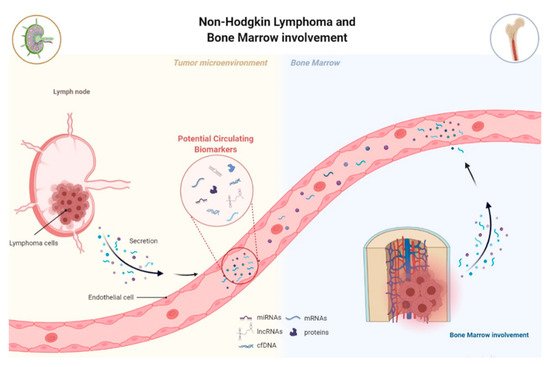
3. MiRNAs and lncRNAs as Potential Biomarkers for NHL
| NHL | miRNA | Expression | Biomarker Utility | Source Material |
Refs. |
|---|---|---|---|---|---|
| DLBCL | miR-155 | Upregulated | Diagnostic | Serum | [42][43] |
| Subclassification | Serum | [44] | |||
| Prognostic of OS, PFS and RFS | Plasma, serum |
[45][46] | |||
| miR-210 | Upregulated | Diagnostic | Serum | [42][47] | |
| let-7b/c | Upregulated | Diagnostic | Serum | [48] | |
| miR-15a | Upregulated | Diagnostic | Serum | [43][47][48] | |
| miR-16-1 | Upregulated | Diagnostic | Serum | [43] | |
| miR-18a | Upregulated | Diagnostic | Serum | [48] | |
| miR-20a/b | Upregulated | Prognostic of OS | Serum | [49] | |
| miR-21 | Upregulated | Diagnostic | Serum | [42][47] | |
| Subclassification | Serum | [50] | |||
| Monitoring | Plasma | [51] | |||
| Prognostic of OS, PFS and RFS | Serum | [42][46][50][52] | |||
| miR-22 | Upregulated | Prognostic of PFS | Serum | [53] | |
| miR-24 | Upregulated | Diagnostic | Serum | [48] | |
| miR-28 | Downregulated | Prognostic of OS, PFS and RFS | Serum | [46] | |
| miR-29c | Upregulated | Diagnostic | Serum | [43] | |
| miR-33a | Downregulated | Prognostic of RFS | Serum | [54] | |
| miR-34 | Downregulated | Diagnostic | Serum | [43] | |
| miR-92a | Downregulated | Diagnostic | Plasma | [55] | |
| Monitoring | |||||
| miR-93 | Upregulated | Prognostic of OS | Serum | [49] | |
| miR-106a/b | Upregulated | Prognostic of OS | Serum | [49] | |
| miR-125b | Upregulation | Prognostic of OS | Serum | [56] | |
| miR-130a | Upregulated | Monitoring | Serum | [56] | |
| miR-130b | upregulation | Prognostic of OS, PFS and RFS | Serum | [46] | |
| miR-181-5p | Downregulated | Subclassification | Serum | [44] | |
| miR-199-5p | Upregulated | Prognostic of OS | Plasma | [45] | |
| miR-224 | Upregulated | Prognostic of RFS | Serum | [54] | |
| miR-323b | Downregulated | Diagnostic | Serum | [57] | |
| miR-326 | Upregulated | Diagnostic | Serum | [58] | |
| miR-375 | Downregulated | Diagnostic | Serum | [58] | |
| miR-431 | Downregulated | Diagnostic | Serum | [57] | |
| miR-455-3p | downregulated | Prognostic of RFS | Serum | [54] | |
| miR-494 | upregulated | Monitoring | Plasma | [51] | |
| miR-520d-3p | Upregulated | Prognostic of RFS | Serum | [54] | |
| miR-1236 | Upregulated | Prognostic of RFS | Serum | [54] | |
| CLL | miR-34a | Upregulated | Diagnostic | Serum | [59] |
| miR-31-5p | Upregulated | Diagnostic | Serum | [59] | |
| miR-150-5p | Upregulated | Diagnostic | Serum | [59] | |
| miR-155-5p | Upregulated | Diagnostic | Serum | [59] | |
| miR-15a-3p | Upregulated | Diagnostic | Serum | [59] | |
| miR-29a-3p | Upregulated | Diagnostic | Serum | [59] |
| NHL | LncRNA | Expression | Biomarker Utility | Source Material |
Refs. |
|---|---|---|---|---|---|
| DLBCL | PEG10 | Upregulated | Diagnostic | Tissue Cell lines |
[60] |
| Prognostic of OS | |||||
| LUNAR1 | Upregulated | Diagnostic | Tissue Cell lines |
[61] | |
| Prognostic of OS and PFS | |||||
| FIRRE | Upregulated | Diagnostic | Tissue Cell lines |
[32] | |
| Prognostic of OS | |||||
| HULC | Upregulated | Diagnostic | Tissue Cell lines |
[31] | |
| Prognostic of OS and PFS | |||||
| LINC01857 | Upregulated | Diagnostic | Tissue Cell lines |
[62] | |
| OR3A4 | Upregulated | Diagnostic | Tissue Cell lines |
[63] | |
| Prognostic of OS | |||||
| ENST00000424690 | Upregulated | Diagnostic | Tissue Cell lines |
[64] | |
| ENST00000425358 | |||||
| NR_026892 | |||||
| ENST00000464929 | Downregulated | ||||
| ENST00000475089 | |||||
| SubSigLnc-17 | - | Diagnostic | Tissue Cell lines |
[65] | |
| Subclassification | |||||
| Prognostic of OS and PFS | |||||
| NONHSAG026900 | Upregulated | Diagnostic | Tissue Cell lines |
[66] | |
| Prognostic of OS and PFS | |||||
| NEAT1_1 | Upregulated | Diagnostic | Tissue Cell lines |
[67] | |
| Prognostic of OS | |||||
| GAS5 | Upregulated | Diagnostic | Tissue Cell lines |
[68] | |
| MIR17HG | Upregulated | Diagnostic | Tissue Cell lines |
||
| HULC | Upregulation | Diagnostic | Tissue Cell lines |
||
| PCA3 | Upregulated | Diagnostic | Tissue Cell lines |
||
| PANDA | Downregulation | Diagnostic | Plasma Tissue |
[30] | |
| Prognostic of OS and RFS | |||||
| TUG1 | Upregulated | Diagnostic | Plasma | [30] | |
| HOTAIR | Upregulated | Diagnostic | Plasma Tissue |
[69][70] | |
| Predictive of Treatment response |
|||||
| Prognostic of OS |
|||||
| XIST | Upregulated | Diagnostic | Plasma | [70] | |
| GAS5 | Downregulated | Diagnostic | Plasma | ||
| Predictive of Treatment response |
|||||
| 6-lncRNA signature | - | Prognostic of OS | Tissue | [71] | |
| FL | RP11-625 L16.3 | Upregulated | Diagnostic | Tissue | [72] |
| RP4-694A7.2 | Upregulated | Diagnostic and subclassification | Tissue | [73] | |
| MCL | LINK-A | Upregulated | Diagnostic | Plasma | [74] |
| GATA6-AS | Downregulated | Diagnostic | Plasma | ||
| MALAT1 | Upregulated | Prognostic of OS and DFS | Tissue Cell lines |
[37] | |
| FOXP4-AS1 | Upregulated | Prognostic of OS and DFS | Plasma | [38] | |
| MORT | Downregulated | Diagnostic | Plasma | [75] | |
| CLL | lincRNA-p21 | Downregulated | Diagnostic | Plasma | [76] |
| MM | TUG1 | Upregulated | Diagnostic | Plasma | |
| MALAT1 | Downregulated | Diagnostic | Plasma | ||
| HOTAIR | Diagnostic | Plasma | |||
| GAS5 | Diagnostic | Plasma |
3.1. MiRNAs and lncRNAs as Non-Invasive Diagnostic Biomarkers
3.2. MiRNAs and lncRNAs as Prognostic Biomarkers
Several studies have been exploring the value of circulating miRNAs as prognostic markers for NHL. In 2008, Lawrie et al. were the first to report that high serum levels of miR-21 were associated with relapse-free survival (RFS) in DLBCL patients, which were later on supported in other studies [42][50]. Similarly, high serum levels of miR-22 at diagnosis in DLBCL were associated with a worse progression-free survival (PFS), independently of the currently used clinical prognostic index [53]. The upregulation of circulating miR-155 and miR-125b was associated with shorter overall survival (OS) of DLBCL patients, while miR-20a/b, miR-93, and miR-106a/b plasma profiles were associated with higher mortality in DLBCL [45][49][56]. Song et al.’s study reported that elevated levels of miR-224, miR-520d-3p, and miR-1236 and lower levels of miR-33a and miR-455-3p were associated with lower medium remission time, and consequently, higher probability of remission, independently of IPI score [54]. A 4-miRNA expression profile (higher levels of miR-21, miR-130b, miR-155; lower levels of miR-28) was shown to be associated with relapse, as well as inferior PFS and OS after R-CHOP, independently of IPI score [46].
he 6-lncRNA signature defined by Sun et al. was shown to be associated with patients OS, independently of standard clinical factors, and permitted to stratify DLBCL patients in high and low-risk groups, improving survival prediction [71]. Moreover, the SubSigLnc-17 profile was not only able to discriminate clinically molecular DLBCL subtypes but also was shown to be significantly associated with patients’ OS and PFS [65]. The expression levels of lncRNA HOTAIR were not only associated with tumor size and clinical stage but also with the presence of B symptoms and IPI score. In fact, higher levels of HOTAIR were associated with better patients’ prognoses, being characterized as an independent predictive biomarker for DLBCL [69][70].
The assessment of patients’ prognosis and decisions on treatment alteration are mostly based on imaging PET-CT and clinical evaluations [81]. However, due to the insufficient sensitivity and specificity of these tools, there is an impending need to identify new predictors that permit the early identification of patients with inherent or acquired the refractory disease during treatment. Detection of miRNAs or lncRNAs can lower the detection limit of disease beyond the capabilities of current methods and create a “window of opportunity” for intervention prior to clinical relapse. Earlier initiation of second-line therapy at a point of minimal tumor burden may improve patients’ outcomes. Song et al. identified a 5-miRNA profile (miR-224, miR-455-3p, miR-1236, miR-33a, and miR-520d-3p) associated with R-CHOP response in DLBCL patients, being a significant predictor of response, independent from the IPI score [54].
Regarding lncRNAs as treatment response biomarkers, Senousy et al. observed that pretreatment circulating levels of HOTAIR were higher, whereas GAS5 were lower in non-responders compared to responders to R-CHOP. Moreover, when performing multivariate analysis, HOTAIR appeared as an independent predictor of R-CHOP failure [70]. DLBCL patients with higher expression levels of NONHSAG026900 were shown to have a better response to chemotherapy compared to patients with lower levels [66].
3.3. Clinical Trials for Potential miRNA and lncRNA Biomarkers
References
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310.
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529.
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402.
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297.
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276.
- Fernandes, M.; Teixeira, A.L.; Medeiros, R. The opportunistic effect of exosomes on Non-Hodgkin Lymphoma microenvironment modulation. Crit. Rev. Oncol. Hematol. 2019, 144, 102825.
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17.
- Schmitt, A.M.; Chang, H.Y. Long noncoding RNAs in cancer pathways. Cancer Cell 2016, 29, 452–463.
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407.
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789.
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA–LncRNA Interactions. In Long Non-Coding RNAs: Methods and Protocols; Feng, Y., Zhang, L., Eds.; Springer: New York, NY, USA, 2016; pp. 271–286.
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015.
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358.
- Si, Y.; Yang, Z.; Ge, Q.; Yu, L.; Yao, M.; Sun, X.; Ren, Z.; Ding, C. Long non-coding RNA Malat1 activated autophagy, hence promoting cell proliferation and inhibiting apoptosis by sponging miR-101 in colorectal cancer. Cell. Mol. Biol. Lett. 2019, 24, 50.
- Fu, Z.; Luo, W.; Wang, J.; Peng, T.; Sun, G.; Shi, J.; Li, Z.; Zhang, B. Malat1 activates autophagy and promotes cell proliferation by sponging miR-101 and upregulating STMN1, RAB5A and ATG4D expression in glioma. Biochem. Biophys. Res. Commun. 2017, 492, 480–486.
- Xiong, Z.; Wang, L.; Wang, Q.; Yuan, Y. LncRNA MALAT1/miR-129 axis promotes glioma tumorigenesis by targeting SOX2. J. Cell. Mol. Med. 2018, 22, 3929–3940.
- Liao, K.; Lin, Y.; Gao, W.; Xiao, Z.; Medina, R.; Dmitriev, P.; Cui, J.; Zhuang, Z.; Zhao, X.; Qiu, Y.; et al. Blocking lncRNA MALAT1/miR-199a/ZHX1 Axis Inhibits Glioblastoma Proliferation and Progression. Mol. Ther. Nucleic Acids 2019, 18, 388–399.
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21.
- Greene, J.; Baird, A.-M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38.
- Spierings, D.C.; McGoldrick, D.; Hamilton-Easton, A.M.; Neale, G.; Murchison, E.P.; Hannon, G.J.; Green, D.R.; Withoff, S. Ordered progression of stage-specific miRNA profiles in the mouse B2 B-cell lineage. Blood J. Am. Soc. Hematol. 2011, 117, 5340–5349.
- Tayari, M.M.; Winkle, M.; Kortman, G.; Sietzema, J.; de Jong, D.; Terpstra, M.; Mestdagh, P.; Kroese, F.G.; Visser, L.; Diepstra, A. Long noncoding RNA expression profiling in normal B-cell subsets and Hodgkin lymphoma reveals hodgkin and reed-sternberg cell–specific long noncoding RNAs. Am. J. Pathol. 2016, 186, 2462–2472.
- Brazão, T.F.; Johnson, J.S.; Müller, J.; Heger, A.; Ponting, C.P.; Tybulewicz, V.L. Long noncoding RNAs in B-cell development and activation. Blood J. Am. Soc. Hematol. 2016, 128, e10–e19.
- Petri, A.; Dybkaer, K.; Bøgsted, M.; Thrue, C.A.; Hagedorn, P.H.; Schmitz, A.; Bødker, J.S.; Johnsen, H.E.; Kauppinen, S. Long noncoding RNA expression during human B-cell development. PLoS ONE 2015, 10, e0138236.
- Casero, D.; Sandoval, S.; Seet, C.S.; Scholes, J.; Zhu, Y.; Ha, V.L.; Luong, A.; Parekh, C.; Crooks, G.M. Long non-coding RNA profiling of human lymphoid progenitor cells reveals transcriptional divergence of B cell and T cell lineages. Nat. Immunol. 2015, 16, 1282–1291.
- Meng, Y.B.; He, X.; Huang, Y.F.; Wu, Q.N.; Zhou, Y.C.; Hao, D.J. Long Noncoding RNA CRNDE Promotes Multiple Myeloma Cell Growth by Suppressing miR-451. Oncol. Res. 2017, 25, 1207–1214.
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a long non-coding RNA responsive to insulin/IGF signaling, regulates genes involved in central metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 372–386.
- Ebralidze, A.K.; Guibal, F.C.; Steidl, U.; Zhang, P.; Lee, S.; Bartholdy, B.; Jorda, M.A.; Petkova, V.; Rosenbauer, F.; Huang, G. PU.1 expression is modulated by the balance of functional sense and antisense RNAs regulated by a shared cis-regulatory element. Genes Dev. 2008, 22, 2085–2092.
- DeKoter, R.P.; Singh, H. Regulation of B lymphocyte and macrophage development by graded expression of PU. 1. Science 2000, 288, 1439–1441.
- Blume, C.J.; Hotz-Wagenblatt, A.; Hüllein, J.; Sellner, L.; Jethwa, A.; Stolz, T.; Slabicki, M.; Lee, K.; Sharathchandra, A.; Benner, A.; et al. p53-dependent non-coding RNA networks in chronic lymphocytic leukemia. Leukemia 2015, 29, 2015–2023.
- Wang, Y.; Zhang, M.; Xu, H.; Wang, Y.; Li, Z.; Chang, Y.; Wang, X.; Fu, X.; Zhou, Z.; Yang, S.; et al. Discovery and validation of the tumor-suppressive function of long noncoding RNA PANDA in human diffuse large B-cell lymphoma through the inactivation of MAPK/ERK signaling pathway. Oncotarget 2017, 8, 72182–72196.
- Peng, W.; Wu, J.; Feng, J. Long noncoding RNA HULC predicts poor clinical outcome and represents pro-oncogenic activity in diffuse large B-cell lymphoma. Biomed. Pharmacother. 2016, 79, 188–193.
- Shi, X.; Cui, Z.; Liu, X.; Wu, S.; Wu, Y.; Fang, F.; Zhao, H. LncRNA FIRRE is activated by MYC and promotes the development of diffuse large B-cell lymphoma via Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2019, 510, 594–600.
- Song, Y.; Gao, F.; Peng, Y.; Yang, X. Long non-coding RNA DBH-AS1 promotes cancer progression in diffuse large B-cell lymphoma by targeting FN1 via RNA-binding protein BUD13. Cell Biol. Int. 2020, 44, 1331–1340.
- Florian, A.K.; Reschke, M.; Ruocco, A.; Ng, C.; Chapuy, B.; Léopold, V.; Sjoberg, M.; Thomas, M.K.; Verma, A.; Ala, U.; et al. The BRAF Pseudogene Functions as a Competitive Endogenous RNA and Induces Lymphoma In Vivo. Cell 2015, 161, 319–332.
- Qian, C.-S.; Li, L.-J.; Huang, H.-W.; Yang, H.-F.; Wu, D.-P. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. 2020, 20, 87.
- Wang, Q.M.; Lian, G.Y.; Song, Y.; Huang, Y.F.; Gong, Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. 2019, 231, 116335.
- Wang, X.; Sehgal, L.; Jain, N.; Khashab, T.; Mathur, R.; Samaniego, F. LncRNA MALAT1 promotes development of mantle cell lymphoma by associating with EZH2. J. Transl. Med. 2016, 14, 346.
- Tao, H.F.; Shen, J.X.; Hou, Z.W.; Chen, S.Y.; Su, Y.Z.; Fang, J.L. lncRNA FOXP4-AS1 predicts poor prognosis and accelerates the progression of mantle cell lymphoma through the miR-423-5p/NACC1 pathway. Oncol. Rep. 2021, 45, 469–480.
- De Rubis, G.; Rajeev Krishnan, S.; Bebawy, M. Liquid Biopsies in Cancer Diagnosis, Monitoring, and Prognosis. Trends Pharmacol. Sci. 2019, 40, 172–186.
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008.
- Shi, T.; Gao, G.; Cao, Y. Long Noncoding RNAs as Novel Biomarkers Have a Promising Future in Cancer Diagnostics. Dis. Markers 2016, 2016, 9085195.
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675.
- Fang, C.; Zhu, D.-X.; Dong, H.-J.; Zhou, Z.-J.; Wang, Y.-H.; Liu, L.; Fan, L.; Miao, K.-R.; Liu, P.; Xu, W.; et al. Serum microRNAs are promising novel biomarkers for diffuse large B cell lymphoma. Ann. Hematol. 2012, 91, 553–559.
- Bedewy, A.M.L.; Elmaghraby, S.M.; Shehata, A.A.; Kandil, N.S. Prognostic Value of miRNA-155 Expression in B-Cell Non-Hodgkin Lymphoma. Turk. J. Haematol. 2017, 34, 207–212.
- Ahmadvand, M.; Eskandari, M.; Pashaiefar, H.; Yaghmaie, M.; Manoochehrabadi, S.; Khakpour, G.; Sheikhsaran, F.; Montazer Zohour, M. Over expression of circulating miR-155 predicts prognosis in diffuse large B-cell lymphoma. Leuk. Res. 2018, 70, 45–48.
- Sun, R.; Zheng, Z.; Wang, L.; Cheng, S.; Shi, Q.; Qu, B.; Fu, D.; Leboeuf, C.; Zhao, Y.; Ye, J.; et al. A novel prognostic model based on four circulating miRNA in diffuse large B-cell lymphoma: Implications for the roles of MDSC and Th17 cells in lymphoma progression. Mol. Oncol. 2021, 15, 246–261.
- Inada, K.; Okoshi, Y.; Cho, Y.; Saito, H.; Iijima, T.; Hori, M.; Kojima, H. Availability of Circulating microRNAs as a Biomarker for the Early Diagnosis of Diffuse Large B-Cell Lymphoma. Blood 2014, 124, 2988.
- Beheshti, A.; Stevenson, K.; Vanderburg, C.; Ravi, D.; McDonald, J.T.; Christie, A.L.; Shigemori, K.; Jester, H.; Weinstock, D.M.; Evens, A.M. Identification of Circulating Serum Multi-MicroRNA Signatures in Human DLBCL Models. Sci. Rep. 2019, 9, 17161.
- Khare, D.; Goldschmidt, N.; Bardugo, A.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Avni, B. Plasma microRNA profiling: Exploring better biomarkers for lymphoma surveillance. PLoS ONE 2017, 12, e0187722.
- Chen, W.; Wang, H.; Chen, H.; Liu, S.; Lu, H.; Kong, D.; Huang, X.; Kong, Q.; Lu, Z. Clinical significance and detection of microRNA-21 in serum of patients with diffuse large B-cell lymphoma in Chinese population. Eur. J. Haematol. 2014, 92, 407–412.
- Cui, Q.; Vari, F.; Cristino, A.S.; Salomon, C.; Rice, G.E.; Sabdia, M.B.; Guanzon, D.; Palma, C.; Mathew, M.; Talaulikar, D.; et al. Circulating cell-free miR-494 and miR-21 are disease response biomarkers associated with interim-positron emission tomography response in patients with diffuse large B-cell lymphoma. Oncotarget 2018, 9, 34644–34657.
- Li, J.; Fu, R.; Yang, L.; Tu, W. miR-21 expression predicts prognosis in diffuse large B-cell lymphoma. Int. J. Clin. Exp. Pathol. 2015, 8, 15019–15024.
- Marchesi, F.; Regazzo, G.; Palombi, F.; Terrenato, I.; Sacconi, A.; Spagnuolo, M.; Donzelli, S.; Marino, M.; Ercolani, C.; Di Benedetto, A.; et al. Serum miR-22 as potential non-invasive predictor of poor clinical outcome in newly diagnosed, uniformly treated patients with diffuse large B-cell lymphoma: An explorative pilot study. J. Exp. Clin. Cancer Res. 2018, 37, 95.
- Song, G.; Gu, L.; Li, J.; Tang, Z.; Liu, H.; Chen, B.; Sun, X.; He, B.; Pan, Y.; Wang, S.; et al. Serum microRNA expression profiling predict response to R-CHOP treatment in diffuse large B cell lymphoma patients. Ann. Hematol. 2014, 93, 1735–1743.
- Ohyashiki, K.; Umezu, T.; Yoshizawa, S.-i.; Ito, Y.; Ohyashiki, M.; Kawashima, H.; Tanaka, M.; Kuroda, M.; Ohyashiki, J.H. Clinical Impact of Down-Regulated Plasma miR-92a Levels in Non-Hodgkin’s Lymphoma. PLoS ONE 2011, 6, e16408.
- Yuan, W.X.; Gui, Y.X.; Na, W.N.; Chao, J.; Yang, X. Circulating microRNA-125b and microRNA-130a expression profiles predict chemoresistance to R-CHOP in diffuse large B-cell lymphoma patients. Oncol. Lett. 2016, 11, 423–432.
- Meng, Y.; Quan, L.; Liu, A. Identification of key microRNAs associated with diffuse large B-cell lymphoma by analyzing serum microRNA expressions. Gene 2018, 642, 205–211.
- Jørgensen, S.; Paulsen, I.W.; Hansen, J.W.; Tholstrup, D.; Hother, C.; Sørensen, E.; Petersen, M.S.; Nielsen, K.R.; Rostgaard, K.; Larsen, M.A.H.; et al. The value of circulating microRNAs for early diagnosis of B-cell lymphoma: A case-control study on historical samples. Sci. Rep. 2020, 10, 9637.
- Filip, A.A.; Grenda, A.; Popek, S.; Koczkodaj, D.; Michalak-Wojnowska, M.; Budzyński, M.; Wąsik-Szczepanek, E.; Zmorzyński, S.; Karczmarczyk, A.; Giannopoulos, K. Expression of circulating miRNAs associated with lymphocyte differentiation and activation in CLL—another piece in the puzzle. Ann. Hematol. 2017, 96, 33–50.
- Peng, W.; Fan, H.; Wu, G.; Wu, J.; Feng, J. Upregulation of long noncoding RNA PEG10 associates with poor prognosis in diffuse large B cell lymphoma with facilitating tumorigenicity. Clin. Exp. Med. 2016, 16, 177–182.
- Peng, W.; Feng, J. Long noncoding RNA LUNAR1 associates with cell proliferation and predicts a poor prognosis in diffuse large B-cell lymphoma. Biomed. Pharmacother. 2016, 77, 65–71.
- Li, Q.; Li, B.; Lu, C.-L.; Wang, J.-Y.; Gao, M.; Gao, W. LncRNA LINC01857 promotes cell growth and diminishes apoptosis via PI3K/mTOR pathway and EMT process by regulating miR-141-3p/MAP4K4 axis in diffuse large B-cell lymphoma. Cancer Gene Ther. 2020.
- Meng, H.; Zhao, B.; Wang, Y. FOXM1-induced upregulation of lncRNA OR3A4 promotes the progression of diffuse large B-cell lymphoma via Wnt/β-catenin signaling pathway. Exp. Mol. Pathol. 2020, 115, 104451.
- Gao, H.-Y.; Wu, B.; Yan, W.; Gong, Z.-M.; Sun, Q.; Wang, H.-H.; Yang, W. Microarray expression profiles of long non-coding RNAs in germinal center-like diffuse large B-cell lymphoma. Oncol. Rep. 2017, 38, 1363–1372.
- Zhou, M.; Zhao, H.; Xu, W.; Bao, S.; Cheng, L.; Sun, J. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol. Cancer 2017, 16, 16.
- Zhao, S.; Fang, S.; Liu, Y.; Li, X.; Liao, S.; Chen, J.; Liu, J.; Zhao, L.; Li, H.; Zhou, W.; et al. The long non-coding RNA NONHSAG026900 predicts prognosis as a favorable biomarker in patients with diffuse large B-cell lymphoma. Oncotarget 2017, 8, 34374.
- Deng, L.; Jiang, L.; Tseng, K.F.; Liu, Y.; Zhang, X.; Dong, R.; Lu, Z.; Wang, X. Aberrant NEAT1_1 expression may be a predictive marker of poor prognosis in diffuse large B cell lymphoma. Cancer Biomark. 2018, 23, 157–164.
- Dousti, F.; Shahrisa, A.; Ansari, H.; Hajjari, M.; Tahmasebi Birgani, Y.; Mohammadiasl, J.; Tahmasebi Birgani, M. Long non-coding RNAs expression levels in diffuse large B-cell lymphoma: An in silico analysis. Pathol. Res. Pract. 2018, 214, 1462–1466.
- Yan, Y.; Han, J.; Li, Z.; Yang, H.; Sui, Y.; Wang, M. Elevated RNA expression of long non-coding HOTAIR promotes cell proliferation and predicts a poor prognosis in patients with diffuse large B cell lymphoma. Mol. Med. Rep. 2016, 13, 5125–5131.
- Senousy, M.A.; El-Abd, A.M.; Abdel-Malek, R.R.; Rizk, S.M. Circulating long non-coding RNAs HOTAIR, Linc-p21, GAS5 and XIST expression profiles in diffuse large B-cell lymphoma: Association with R-CHOP responsiveness. Sci. Rep. 2021, 11, 2095.
- Sun, J.; Cheng, L.; Shi, H.; Zhang, Z.; Zhao, H.; Wang, Z.; Zhou, M. A potential panel of six-long non-coding RNA signature to improve survival prediction of diffuse large-B-cell lymphoma. Sci. Rep. 2016, 6, 27842.
- Pan, Y.; Li, H.; Guo, Y.; Luo, Y.; Li, H.; Xu, Y.; Deng, J.; Sun, B. A pilot study of long noncoding RNA expression profiling by microarray in follicular lymphoma. Gene 2016, 577, 132–139.
- Roisman, A.; Castellano, G.; Navarro, A.; Gonzalez-Farre, B.; Pérez-Galan, P.; Esteve-Codina, A.; Dabad, M.; Heath, S.; Gut, M.; Bosio, M.; et al. Differential expression of long non-coding RNAs are related to proliferation and histological diversity in follicular lymphomas. Br. J. Haematol. 2019, 184, 373–383.
- Zhang, Y.; Lu, P.; Du, H.; Zhang, L. LINK-A lncRNA Promotes Proliferation and Inhibits Apoptosis of Mantle Cell Lymphoma Cell by Upregulating Survivin. Med. Sci. Monit. 2019, 25, 365–370.
- Tang, X.; Long, Y.; Xu, L.; Yan, X. LncRNA MORT Inhibits Cancer Cell Proliferation and Promotes Apoptosis in Mantle Cell Lymphoma by Upregulating miRNA-16. Cancer Manag. Res. 2020, 12, 2119–2125.
- Isin, M.; Ozgur, E.; Cetin, G.; Erten, N.; Aktan, M.; Gezer, U.; Dalay, N. Investigation of circulating lncRNAs in B-cell neoplasms. Clin. Chim. Acta 2014, 431, 255–259.
- Beheshti, A.; Vanderburg, C.; McDonald, J.T.; Ramkumar, C.; Kadungure, T.; Zhang, H.; Gartenhaus, R.B.; Evens, A.M. A Circulating microRNA Signature Predicts Age-Based Development of Lymphoma. PLoS ONE 2017, 12, e0170521.
- Verma, A.; Jiang, Y.; Du, W.; Fairchild, L.; Melnick, A.; Elemento, O. Transcriptome sequencing reveals thousands of novel long non-coding RNAs in B cell lymphoma. Genome Med. 2015, 7, 110.
- Qi, P.; Zhou, X.-y.; Du, X. Circulating long non-coding RNAs in cancer: Current status and future perspectives. Mol. Cancer 2016, 15, 39.
- Diez-Fraile, A.; Ceulaer, J.D.; Derpoorter, C.; Spaas, C.; Backer, T.D.; Lamoral, P.; Abeloos, J.; Lammens, T. Circulating Non-Coding RNAs in Head and Neck Cancer: Roles in Diagnosis, Prognosis, and Therapy Monitoring. Cells 2021, 10, 48.
- Adams, H.J.A.; Nievelstein, R.A.J.; Kwee, T.C. Prognostic value of interim and end-of-treatment FDG-PET in follicular lymphoma: A systematic review. Ann. Hematol. 2016, 95, 11–18.
- Study Biomarkers in Samples From Patients With B-Cell Acute Lymphoblastic Leukemia. Available online: https://ClinicalTrials.gov/show/NCT01505699 (accessed on 3 June 2021).
- Studying Biomarkers in Cell Samples From Patients With Acute Myeloid Leukemia. Available online: https://ClinicalTrials.gov/show/NCT01057199 (accessed on 3 June 2021).
- Circulating microRNAs as Disease Markers in Pediatric Cancers. Available online: https://ClinicalTrials.gov/show/NCT01541800 (accessed on 3 June 2021).
- Qian, Y.; Shi, L.; Luo, Z. Long Non-coding RNAs in Cancer: Implications for Diagnosis, Prognosis, and Therapy. Front. Med. 2020, 7, 612393.
- ClinicalTrials.gov. 2021. Available online: https://clinicaltrials.gov/ct2/about-site/new (accessed on 3 June 2021).
- Pal, R.P.; Maitra, N.U.; Mellon, J.K.; Khan, M.A. Defining prostate cancer risk before prostate biopsy. Urol. Oncol. 2013, 31, 1408–1418.
- Immunophenotyping and Xist Gene in AML. Available online: https://ClinicalTrials.gov/show/NCT04288739 (accessed on 3 June 2021).

