Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Indra Prasad Subedi | + 1917 word(s) | 1917 | 2021-12-17 07:18:37 | | | |
| 2 | Beatrix Zheng | + 130 word(s) | 2047 | 2021-12-23 02:13:16 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Subedi, I. Forest Ant (Hymenoptera: Formicidae) Diversity in Nepal. Encyclopedia. Available online: https://encyclopedia.pub/entry/17463 (accessed on 08 August 2026).
Subedi I. Forest Ant (Hymenoptera: Formicidae) Diversity in Nepal. Encyclopedia. Available at: https://encyclopedia.pub/entry/17463. Accessed August 08, 2026.
Subedi, Indra. "Forest Ant (Hymenoptera: Formicidae) Diversity in Nepal" Encyclopedia, https://encyclopedia.pub/entry/17463 (accessed August 08, 2026).
Subedi, I. (2021, December 22). Forest Ant (Hymenoptera: Formicidae) Diversity in Nepal. In Encyclopedia. https://encyclopedia.pub/entry/17463
Subedi, Indra. "Forest Ant (Hymenoptera: Formicidae) Diversity in Nepal." Encyclopedia. Web. 22 December, 2021.
Copy Citation
Seventy ant species from thirty-six genera and six subfamilies were recorded from eastern, central, and western regions of Nepal using vegetation beating, sweeping, and hand collection methods. The research also discovered five genera and nine species new for the country, as well as eight tramp species, four of which are major ecological, agricultural, and/or household pests. Ant diversity was found to decrease with increasing elevation. The assessment of ant diversity using multiple sampling methods that cover all seasons and forest types may be useful in obtaining complete ant diversity data. Early intervention through sustainable forest management initiatives would aid in preventing invasive ant incursions in the forests of Nepal.
Himalaya
invasive species
Nepalese ants
new record
species richness
1. Introduction
Ants (family: Formicidae) occur in almost all terrestrial ecosystems, while tropical regions harbor peaks of their diversity [1]. Some ants are key predators [2][3], ecosystem engineers [4][5][6], seed dispersers [7][8], and biocontrol agents [9][10]. However, some ant species are notorious pests of households, agriculture, and forests. Five ant species (Anoplolepis gracilipes, Linepithema humile, Pheidole megacephala, Wasmannia auropunctata, and Solenopsis invicta) are highly damaging invaders and listed amongst the 100 worst invasive alien species of the world [11]. Ant diversity is noted to be higher in undisturbed primary forests than in secondary forests [12] and is often considered as an indicator of biodiversity and forest health [13][14]. There are two major ant species distribution patterns: decreasing with increasing elevations [15] and mid-elevation peaks [16].
Nepal has been home to eight subfamilies, 48 genera, and 133 species of ants [17][18]. Considering the unique geographical and ecological diversity, many ant species are yet to be discovered, and their ecology (diversity and distribution) is yet to be studied in the country. Since the first description of two ant species from Nepal, Aphaenogaster pachei and Myrmica pachei, by Forel [19], Nepalese ants have received occasional attention from scientists. Nepalese myrmecology is still in its early stages. There have previously been no publications based on a focused systematic survey of Nepalese ants. Collingwood [20] published the first Nepalese ant list, which included 34 species, while Subedi et al. [17] published the recent updated checklist, which included 128 species. Twenty-one ant species have Nepal as type locality, and nine of them are endemic to the country [17]. The majority of the current information about Nepalese ants is based on hand-collected specimens. This implies the need to conduct systematic surveys using multiple collection methods. However, a dearth of taxonomic expertise and equipped infrastructure in the country pose a challenge in the study of ants.
Forest biodiversity conservation is an important objective of forest management [21][22][23]. Old-growth forests support greater biodiversity than plantation forests [24]. Plantation forests can also play a role in conserving biodiversity [25]. An assessment of forest biodiversity is essential to ensure ecosystem integrity and the sustainability of ecosystem functions [26]. Inventories of entire forest biota are almost impossible, and, thus, bioindicators have been used in monitoring. Ants are ideal candidates in monitoring ecosystem conditions because they occur everywhere and are numerically abundant in both intact and disturbed habitats [27][28], can be easily sampled [29], and are sensitive and rapid responders to environmental variables [27][30]. Ants have been widely used as bioindicators for ecosystem management and biodiversity restoration [31], and, further, they are a useful tool in predicting human impacts on forests [32]. Detecting the presence or absence of invasive or native ‘weedy’ ants can provide valuable information for monitoring [31]. It is extremely difficult to eradicate invasive ants once they have been introduced into natural ecosystems, where they can have a variety of direct and indirect effects on native ant and non-ant taxa [33]. As a result, it is critical to plan ahead of time and take the necessary precautions to avoid possible incursions.
With ants being indicators of forest health, the interaction of ants and forest health must be studied closely; the assessment of the association of ants with different forest regimes is imperative. The present study focuses on the inventory of ants in various forest types along the longitudinal (east, central, and west) and latitudinal (north to south in Tarai, Siwalik, middle mountains, and high mountains) gradients. It aims to provide a baseline for further research on forest–ant interaction and sustainable forest management in Nepal.
2. Current Insights
2.1. Ant Species Diversity
This study represents the first nationwide survey of forest ants in Nepal. Species lists are important tools for species conservation because they provide a solid understanding of the current state of the biota, which can serve as a foundation for conservation actions [34]. The species–accumulation curve revealed that the curve approached the asymptote but was not completely leveled off (Figure 1). Despite the fact that the majority of the species were captured in the study areas, additional sampling efforts involving multiple collection methods would result in the capture of a few more species. Crematogaster (208 individuals, 42 occurrences, 6 species) and Camponotus (181 individuals, 40 occurrences, 6 species) were the most diverse genera. Of all the species, Oecophylla smaragdina was recorded most often (38 occurrences) during the study, while the genus Pheidole was recorded from Tarai and Siwalik and the hills of the western, central, and eastern regions. Seventeen genera were represented by only one species in our collection, and eight genera were collected from a single site. In Nepal, the richness of Oriental genera (such as Oecophylla, Polyrhachis, Prenolepis, Carebara, Lophomyrmex, Leptogenys, Meranoplus, and Tetraponera) outnumbered the Palaearctic genera (such as Myrmica, Lasius, and Temnothorax) as in southern China [35]. Several genera were found to be common throughout Tarai and Siwalik and the hills of western, central, and eastern regions, while others were found to be peculiar to a specific location (Figure 2 and Figure 3), with the Palearctic genera being reported at high elevations. Although it is difficult to make an exact comparison of this study with other studies because objectives, sampling methods, the area covered, and identification levels vary, our findings are consistent with ant inventories in our neighboring countries, particularly in dominant subfamilies and commonly occurring genera, such as southern China [35], Yunnan, China [36], northwestern Siwalik, India [37], southwest China [38], Jammu and Kashmir, India [39]. Furthermore, our findings agree with those of a previous study that recognized Camponotus, Pheidole, and Crematogaster as the three most species-rich genera on a global scale [40]. Weaver ants (O. smaragdina) are conspicuous arboreal ants with more than 2700 site records from 21 countries [10]. They share beneficial traits with other ant species and cause cascading effects to lower trophic levels, reducing pest number and damage in orchards and forests [41].
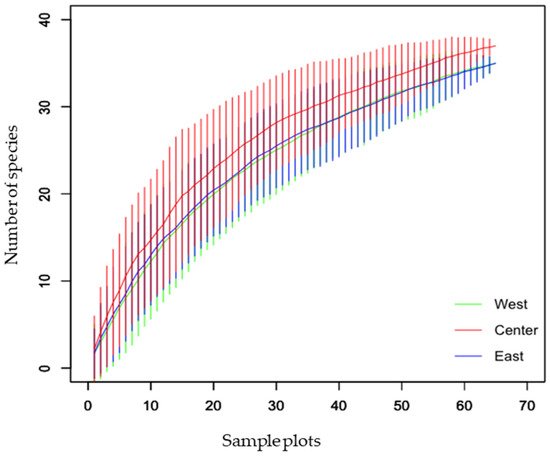
Figure 1. Sample-based species–accumulation curve of ant species in western, central, and eastern regions of Nepal.
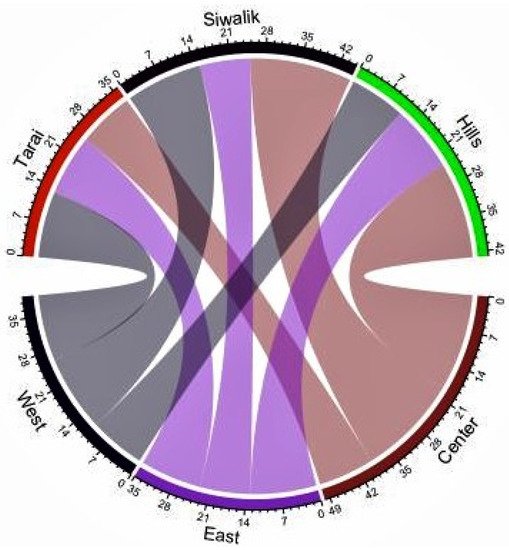
Figure 2. Chord diagram showing shared ant genera in Tarai and Siwalik and the hills of western, central, and eastern regions of Nepal.
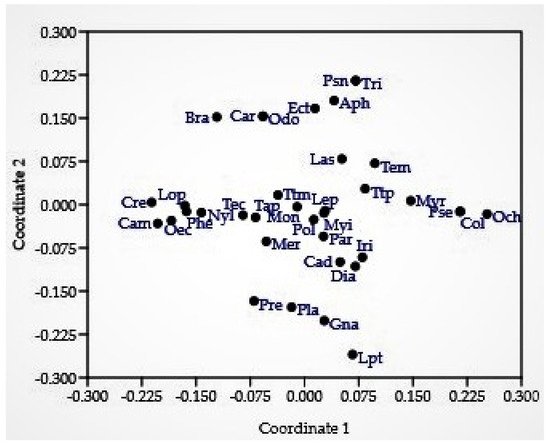
Figure 3. Non-metric dimensional scaling (Bray–Curtis similarity index). Legend genera: Aph: Aphaenogaster, Bra: Brachyponera, Cam: Camponotus, Cad: Cardiocondyla, Car: Carebara, Col: Colobopsis, Cre: Crematogaster, Dia: Diacamma, Ect: Ectomomyrmex, Gna: Gnamptogenys, Iri: Iridomyrmex, Las: Lasius, Lep: Lepisiota, Lpt: Leptogenys, Lop: Lophomyrmex, Mer: Meranoplus, Mon: Monomorium, Myr: Myrmica, Myi: Myrmicaria, Nyl: Nylanderia, Och: Ochetellus, Odo: Odontoponera, Oec: Oecophylla, Par: Paratrechina, Phe: Pheidole, Pla: Plagiolepis, Pol: Polyrhachis, Pre: Prenolepis, Pse: Pseudolasius, Psn: Pseudoneoponera, Tap: Tapinoma, Tec: Technomyrmex, Tem: Temnothorax, Ttm: Tetramorium, Ttp: Tetraponera, Tri: Trichomyrmex.
2.2. Distribution of Ants in Nepal
The variation in the distribution of the forest ants of Nepal with elevation is in line with other studies. The highest number of ant genera in our study was recorded below 1000 m asl, and there was a decreasing pattern with the increase in elevation (Figure 4) as reported by Subedi and Budha [42] when they extrapolated available elevation-related records of ants of Nepal from 200 m to 4550 m. A decreasing pattern is one of the most common patterns of ant species richness [42][43]. A number of studies from different parts of the world also reported a decreasing pattern of species richness with elevation, such as those from Mount Kinabalu [44], the Mediterranean and the oro-Mediterranean parts of Montenegro [45], rainforest in subtropical Queensland [46], wet forest on Costa Rica’s Atlantic slope [47], Imbak Canyon [48], Hengduan Mountains [49], and eastern Himalaya [15]. The most common pattern of species richness along the elevation gradient in the Nepal Himalaya seems to be a unimodal pattern [50][51][52]. However, the elevation with maximum richness is not similar for different taxa [53][54].
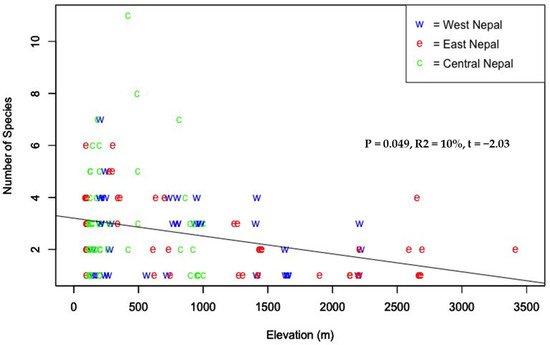
Figure 4. Species richness of ants along elevation gradient in Nepal.
2.3. Ants and Forest Health
In Nepal, we recorded eight common tramp species, including M. pharaonis, P. longicornis, T. melanocephalum, and T. destructor. These four species are amongst twelve cosmopolitan ants that have become significant ecological, agricultural, and/or household pests [55]. Wetterer [56] considered P. longicornis to be the most widely dispersed ant species, with records from both the Old and New Worlds in both the northern and southern hemispheres. Tramp and invasive species are geographically widespread, and observations of functional groups and biogeography may help to better understand the factors that contribute to their spread [57]. Most of the widespread ants belong to the subfamily Myrmicinae and, more specifically, to the following functional groups: cryptic, opportunist, and generalized myrmicine [57].
Ants are indispensable components for the maintenance and appropriate functioning of most terrestrial ecosystems and resulting ecosystem services and disservices [58]. Ant species composition strongly influences seed dispersal by ants [59], and ant communities are very useful for the rapid assessment of terrestrial ecosystem health [60]. Ants play a crucial ecological role by helping in soil formation and by increasing its fertility [61][62]. Three ant species, including O. smaragdina, have been identified as predators of the sal heartwood borer (Hoplocerambyx spinicornis) in nature [63]. In our study, we also frequently observed O. smaragdina in Sal forests.
Of the 42 cosmopolitan ants, only 12 species are major ecological, agricultural, and/or household pests [55], and 4 of them were recorded in Tarai and Siwalik and the mid-mountains of the western, central, and eastern regions of the country. Invasive ant characteristics, such as greater abundance, aggressive behavior, and attraction to high-carbohydrate resources, may result in a high and low risk of deleterious effects on plants, resulting in a negative conservation impact [64]. Bharti and Sharma [16] revealed that the most invasive ant species occurred at lower altitudes, indicating their correlation with the disturbance of the ecosystem.
When used as bioindicators, ants can help detect early signs of habitat disturbance and can help develop management strategies [65]. Invasive ant eradication is critical for biodiversity conservation; however, more than half of the global eradication efforts were not successful [66]. The initiatives aimed to eradicate 11 species around the world, namely Anoplolepis gracilipes, Linepithema humile, Pheidole megacephala, Wasmannia auropunctata, Solenopsis invicta, S. geminata, T. melanocephalum, Lepisiota frauenfeldi, Myrmecia brevinoda, Monomorium indicum, and P. longicornis [66]. Two of the eleven species causing problems around the world, namely P. longicornis, and T. melanocephalum, were also recorded in our study. Other invasive ants, such as A. gracilipes and P. megacephala that are common in neighboring countries are not yet known in Nepal. We believe that studies focusing on the tramp and invasive species of ants in urban areas should be conducted in order to complete their inventory in the country.
MoFE [67] identified invasive species expansion as one of the direct drivers of deforestation and forest degradation, posing a very high effect in Tarai and the Siwaliks, a low effect in the mid-mountains, and a very low effect in the high mountains. Using ants as bioindicators to assess ecosystem health, Bharti and Sharma [16] discovered that the invasive species are seriously threatening the native species of the primary and secondary forests of the Himalaya. They also stated that while ant invasions are currently limited to lower mountain ranges, as global temperatures rise, they may spread to highlands.
The vast majority of insect species are herbivores, but only nine insect orders, including Hymenoptera (ants, wasps, bees, hornets, and sawflies), have species that feed on live plants [68]. Larger carpenter ants (Camponotus sp.) may excavate solid wood in living tree trunks, making them major forest pests [69]. In Nepal, pests and pathogens have invaded all types of forests, resulting in serious consequences, such as forest degradation and biodiversity loss [70].
The early detection of invasive species, pests, and pathogens is critical for preventing damage because early control activities are more feasible and effective [70][71]. Thorough monitoring of invasive ant species encroachment in forest areas is critical for protecting fragile ecosystems and maintaining sound forest health. With increased global travel and trade, several other undocumented tramp/invasive ants may occur or be introduced into the country, particularly in urban areas. A more concentrated collection of tramp/invasive ants in urban areas is necessary to identify them and prevent their possible incursion into natural forests.
References
- Guénard, B. An overview of the species and ecological diversity of ants. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2013.
- Thurman, J.H.; Northfield, T.D.; Snyder, W.E. Weaver ants provide ecosystem services to tropical tree crops. Front. Ecol. Evol. 2019, 7, 120.
- Ohyama, L.; King, J.R.; Jenkins, D.G. Are tiny subterranean ants top predators affecting aboveground ant communities? Ecology 2020, 101, e03084.
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244.
- Frouz, J.; Jilková, V. The effect of ants on soil properties and processes (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 191–199.
- Kovář, P.; Vojtíšek, P.; Zentsová, I. Ants as ecosystem engineers in natural restoration of human made habitats. J. Landsc. Ecol. 2013, 6, 18–31.
- Warren, R.J.; Giladi, I. Ant-mediated seed dispersal: A few ant species (Hymenoptera: Formicidae) benefit many plants. Myrmecol. News 2014, 20, 129–140.
- Anjos, D.V.; Andersen, A.N.; Carvalho, R.L.; Sousa, R.M.; Del-Claro, K. Switching roles from antagonist to mutualist: A harvester ant as a key seed disperser of a myrmecochorous plant. Ecol. Entomol. 2020, 45, 1063–1070.
- Offenberg, J.; Cuc, N.T.T.; Wiwatwitaya, D. The effectiveness of weaver ant (Oecophylla smaragdina) biocontrol in Southeast Asian citrus and mango. Asian Myrmecol. 2013, 5, 139–149.
- Wetterer, J.K. Geographic distribution of the weaver ant Oecophylla smaragdina. Asian Myrmecol. 2017, 9, e009004.
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12.
- Wilkie, K.T.R.; Mertl, A.L.; Traniello, J.F. Diversity of ground-dwelling ants (Hymenoptera: Formicidae) in primary and secondary forests in Amazonian Ecuador. Myrmecol. News 2009, 12, 139–147.
- Stephens, S.S.; Wagner, M.R. Using ground foraging ant (Hymenoptera: Formicidae) functional groups as bioindicators of forest health in northern Arizona ponderosa pine forests. Environ. Entomol. 2006, 35, 937–949.
- Majer, J.D.; Orabi, G.; Bisevac, L. Ants (Hymenoptera: Formicidae) pass the bioindicator scorecard. Myrmecol. News 2007, 10, 69–76.
- Marathe, A.; Priyadarsanan, D.R.; Krishnaswamy, J.; Shanker, K. Spatial and climatic variables independently drive elevational gradients in ant species richness in the Eastern Himalaya. PLoS ONE 2020, 15, e0227628.
- Bharti, H.; Sharma, Y.P.; Bharti, M.; Pfeiffer, M. Ant species richness, endemicity and functional groups, along an elevational gradient in the Himalayas. Asian Myrmecol. 2013, 5, 79–101.
- Subedi, I.P.; Budha, P.B.; Bharti, H.; Alonso, L. An updated checklist of Nepalese ants (Hymenoptera, Formicidae). ZooKeys 2020, 1006, 99–136.
- Subedi, I.P.; Budha, P.B.; Bharti, H.; Alonso, L.; Yamane, S. First Record of the Ant Subgenus Orthonotomyrmex of the Genus Camponotus from Nepal (Hymenoptera, Formicidae). Zoodiversity 2021, 55, 279–284.
- Forel, A. Les fourmis de l’Himalaya. Bull. De La Société Vaud. Des. Sci. Nat. 1906, 42, 79–94.
- Collingwood, C. Formicidae (Hymenoptera: Aculeata) from Nepal. Khumbu Himal 1970, 3, 371–387.
- Lindenmayer, D.; Franklin, J.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445.
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950.
- Junninen, K.; Penttilä, R.; Martikainen, P. Fallen retention aspen trees on clear-cuts can be important habitats for red-listed polypores: A case study in Finland. Biodivers. Conserv. 2007, 16, 475–490.
- Ohsawa, M. Species richness and composition of Curculionidae (Coleoptera) in a conifer plantation, secondary forest, and old-growth forest in the central mountainous region of Japan. Ecol. Res. 2005, 20, 632–645.
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95.
- Ferris, R.; Humphrey, J. A review of potential biodiversity indicators for application in British forests. Forestry 1999, 72, 313–328.
- Majer, J. Ants: Bio-indicators of minesite rehabilitation, land-use, and land conservation. Environ. Manage 1983, 7, 375–383.
- Hoffmann, B.D.; Griffiths, A.D.; Andersen, A.N. Responses of ant communities to dry sulfur deposition from mining emissions in semi-arid tropical Australia, with implications for the use of functional groups. Austral Ecol. 2000, 25, 653–663.
- Agosti, D.; Alonso, L.E. The ALL protocol: A standard protocol for the collection of ground-dwelling ants. In Ants. Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 204–206.
- Alonso, L.E. Ants as indicators of diversity. In Ants. Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 80–88.
- Underwood, E.C.; Fisher, B.L. The role of ants in conservation monitoring: If, when, and how. Biol. Conserv. 2006, 132, 166–182.
- Ribas, C.R.; Schoereder, J.H. Ant communities, environmental characteristics and their implications for conservation in the Brazilian Pantanal. Biodivers.Conserv. 2007, 16, 1511–1520.
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233.
- Pfeiffer, M.; Mezger, D.; Hosoishi, S.; Bakhtiar, E.; Kohout, R.J. The Formicidae of Borneo (Insecta: Hymenoptera): A preliminary species list. Asian Myrmecol. 2011, 4, 9–58.
- Fellowes, J.R. Ant (Hymenoptera: Formicidae) genera in southern China: Observations on the Oriental-Palaearctic boundary. Myrmecol. Nachr. 2006, 8, 239–249.
- Liu, C.; Guénard, B.; Hita Garcia, F.; Yamane, S.; Blanchard, B.; Yang, D.; Economo, E. New records of ant species from Yunnan, China. ZooKeys 2015, 477, 17–78.
- Bharti, H.; Wachkoo, A.A.; Kumar, R. First inventory of ants (Hymenoptera: Formicidae) in northwestern Shivalik, India. Halteres 2017, 8, 33–68.
- Fontanilla, A.M.; Nakamura, A.; Xu, Z.; Cao, M.; Kitching, R.L.; Tang, Y.; Burwell, C.J. Taxonomic and functional ant diversity along tropical, subtropical, and subalpine elevational transects in Southwest China. Insects 2019, 10, 128.
- Wachkoo, A.A.; Akbar, S.A.; Jan, U.; Shah, G.M. Taxonomic inventory of ants (Hymenoptera: Formicidae) in Jammu and Kashmir state. In Biodiversity of the Himalaya: Jammu and Kashmir State; Springer Nature: Basingstoke, UK, 2020; pp. 733–747.
- Wilson, E.O. Which are the most prevalent ant genera. Studia Entomol. 1976, 19, 187–200.
- Offenberg, J. Ants as tools in sustainable agriculture. J. Appl. Ecol. 2015, 52, 1197–1205.
- Subedi, I.P.; Budha, P.B. Diversity and distribution patterns of ants along elevational gradients. Nepal. J. Zool. 2020, 4, 44–49.
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239.
- Brühl, C.A.; Mohamed, M.; Linsenmair, K.E. Altitudinal distribution of leaf litter ants along a transect in primary forests on Mount Kinabalu, Sabah, Malaysia. J. Trop. Ecol. 1999, 15, 265–277.
- Karaman, M.G. Zoogeography, diversity and altitudinal distribution of ants (Hymenoptera: Formicidae) in the Mediterranean and the oro-Mediterranean parts of Montenegro. North West J. Zool. 2011, 7, 26–34.
- Burwell, C.J.; Nakamura, A. Distribution of ant species along an altitudinal transect in continuous rainforest in subtropical Queensland, Australia. Mem. Qld. Mus. 2011, 55, 391–411.
- Longino, J.T.; Colwell, R.K. Density compensation, species composition, and richness of ants on a neotropical elevational gradient. Ecosphere 2011, 2, 1–20.
- Yusah, K.M.; Turner, E.C.; Yahya, B.E.; Fayle, T.M. An elevational gradient in litter-dwelling ant communities in Imbak Canyon, Sabah, Malaysia. J. Trop. Biol. Conserv. JTBC 2012, 9, 192–199.
- Liu, C.; Dudley, K.L.; Xu, Z.H.; Economo, E.P. Mountain metacommunities: Climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 2018, 41, 101–112.
- Bhattarai, K.R.; Vetaas, O.R. Can Rapoport’s rule explain tree species richness along the Himalayan elevation gradient, Nepal? Diver. Distrib. 2006, 12, 373–378.
- Acharya, K.P.; Vetaas, O.R.; Birks, H. Orchid species richness along Himalayan elevational gradients. J. Biogeogr. 2011, 38, 1821–1833.
- Manish, K.; Pandit, M.K. Phylogenetic diversity, structure and diversification patterns of endemic plants along the elevational gradient in the Eastern Himalaya. Plant Ecol. Divers. 2018, 11, 501–513.
- Grau, O.; Grytnes, J.A.; Birks, H. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915.
- Baniya, C.B.; Solhøy, T.; Gauslaa, Y.; Palmer, M.W. The elevation gradient of lichen species richness in Nepal. Lichenologist 2010, 42, 83–96.
- Wetterer, J.K. Geographic origin and spread of cosmopolitan ants (Hymenoptera: Formicidae). Halteres 2015, 6, 66–78.
- Wetterer, J.K. Worldwide spread of the longhorn crazy ant, Paratrechina longicornis (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 137–149.
- McGlynn, T.P. The worldwide transfer of ants: Geographical distribution and ecological invasions. J. Biogeogr. 1999, 26, 535–548.
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The little things that run the world revisited: A review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol. News 2012, 17, 133–146.
- Gorb, S.N.; Gorb, E.V. Effects of ant species composition on seed removal in deciduous forest in eastern Europe. Oikos 1999, 84, 110–118.
- Wike, L.D.; Martin, F.D.; Paller, M.H.; Nelson, E.A. Impact of forest seral stage on use of ant communities for rapid assessment of terrestrial ecosystem health. J. Insect Sci. 2010, 10, 77.
- Brener, A.G.F.; Silva, J.F. Leaf-cutting ant nests and soil fertility in a well-drained savanna in western Venezuela. Biotropica 1995, 17, 250–254.
- Knoepp, J.D.; Coleman, D.C.; Crossley, D., Jr.; Clark, J.S. Biological indices of soil quality: An ecosystem case study of their use. For. Ecol. Manag. 2000, 138, 357–368.
- Roychoudhury, N. New record of natural enemies of sal heartwood borer, Hoplocerambyx spinicornis newman (Coleoptera: Cerambycidae). Pestology 2017, 41, 33–38.
- Lach, L. Invasive ants: Unwanted partners in ant-plant interactions? Ann. Mo. Bot. Gard. 2003, 90, 91–108.
- Verdinelli, M.; Yakhlef, S.E.B.; Cossu, C.S.; Pilia, O.; Mannu, R. Variability of ant community composition in cork oak woodlands across the Mediterranean region: Implications for forest management. iForest 2017, 10, 707.
- Hoffmann, B.D.; Luque, G.M.; Bellard, C.; Holmes, N.D.; Donlan, C.J. Improving invasive ant eradication as a conservation tool: A review. Biol. Conserv. 2016, 198, 37–49.
- MoFE Nepal National REDD+ Strategy (2018–2022); Ministry of Forests and Environment, Government of Nepal, Singh Durbar: Kathmandu, Nepal, 2018.
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities; Cambridge University Press: Cambridge, UK, 2011; p. 828.
- Rossi, N.; Feldhaar, H. Carpenter ants. In Encyclopedia of Social Insects; Springer: Cham, Switzerland, 2020; pp. 973–978.
- Malla, R.; Pokharel, K.K. Forest Pests and Pathogens Problem in Different Forest Types of Nepal; Department of Forest Research and Survey: Kathmandu, Nepal, 2018.
- Finley, K.; Chhin, S. Forest health management and detection of invasive forest insects. Resources 2016, 5, 18.
More
Information
Subjects:
Entomology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Entry Collection:
Environmental Sciences
Revisions:
2 times
(View History)
Update Date:
23 Dec 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No