 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | khadija amin | + 8855 word(s) | 8855 | 2021-10-26 08:11:10 | | | |
| 2 | Lindsay Dong | Meta information modification | 8855 | 2021-10-29 08:53:18 | | |
Video Upload Options
Hyperthermophilic Archaea colonizing unnatural habitats of extremes conditions such as volcanoes and deep-sea hydrothermal vents represent an unmeasurable bioresource for enzymes used in various industrial applications. Their enzymes show distinct structural and functional properties and are resistant to extreme conditions of temperature and pressure where their mesophilic homologs fail. In this review, we will outline carbohydrate-active enzymes (CAZymes) from hyperthermophilic Archaea with specific focus on the two largest families, glycoside hydrolases (GHs) and glycosyltransferases (GTs). We will present the latest advances on these enzymes particularly in the light of novel accumulating data from genomics and metagenomics sequencing technologies. We will discuss the contribution of these enzymes from hyperthermophilic Archaea to industrial applications and put the emphasis on newly identifed enzymes. We will highlight their common biochemical and distinct features. Finally, we will overview the areas that remain to be explored to identify novel promising hyperthermozymes.
1. Introduction
Archaea are prokaryotic microorganisms with distinct biochemical and physiological traits and have important ecological significance due to the ability of many archaeal members to thrive in extreme habitats. Due to some of their unusual characteristics, Archaea have largely contributed to the development of novel biotechnological processes to produce highly valuable bioproducts [1]. In particular, hyperthermophilic Archaea that colonize extreme environments of temperature have recently received great interest and constitute promising bioresource of biocatalysts for high-temperature industrial processes [2].
2. Hyperthermophilic GHs from Archaea
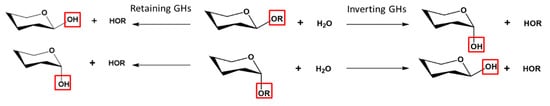
2.1. General Features and Reaction Mechanisms
2.2. Glycoside Hydrolases (GHs) in the Genomes of Hyperthermophilic Archaea
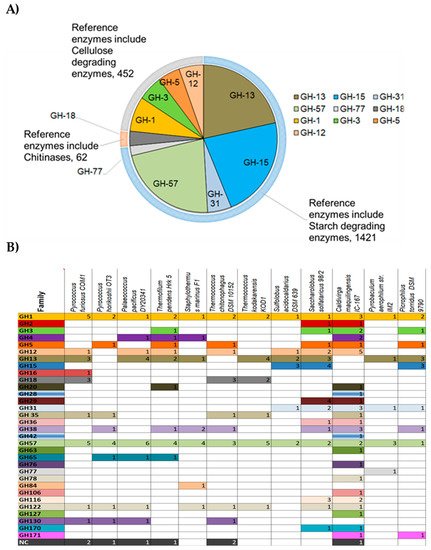
2.3. Biochemical Features and Biotechnological Applications
2.3.1. Starch Degrading Enzymes
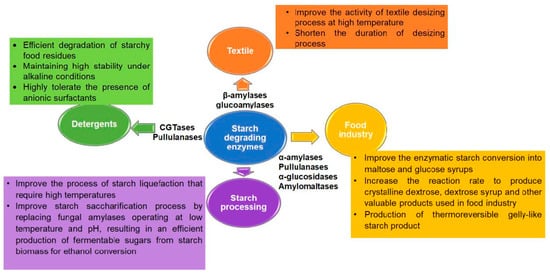
α-Amylases
β-Amylases and Glucoamylases
Pullulanases
Isoamylases
α-Glucosidases
Cyclodextrin Glucanotransferases (CGTases)
Amylomaltases
2.3.2. Cellulose Hydrolyzing Enzymes

Endoglucanases
β-Glucosidases
2.3.3. Chitinases

2.4. Metagenomics of Extreme Environments for the Discovery of GHs from Hyperthermophilic Archaea
3. Glycosyltransferases (GTs)
3.1. General Features and Reaction Mechanism
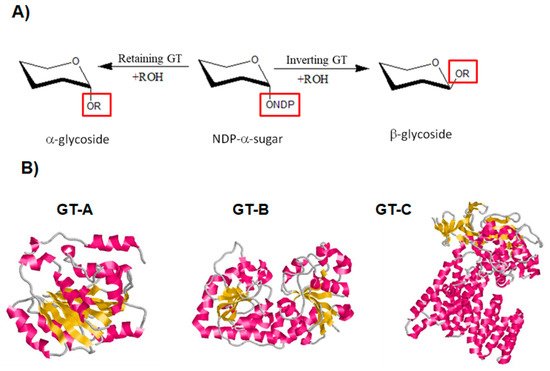
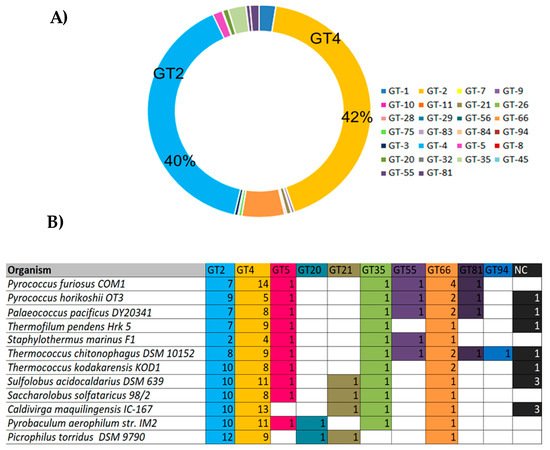
3.2. Hyperthermophilic GTs in Archaea: Biochemical Features and Biotechnological Applications
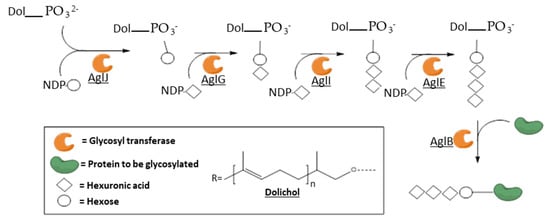
3.3. Membrane-Associated Archaeal GTs: The Protein N-Glycosylation Pathway
3.4. Soluble GTs
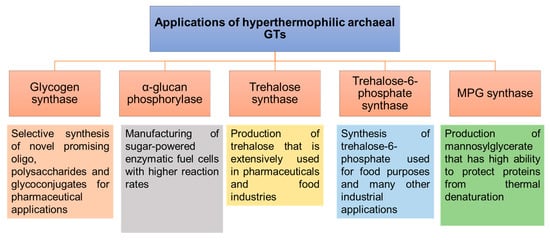
3.4.1. Glycogen Synthase (GS)
3.4.2. α-Glucan Phosphorylase/Maltodextrin Phosphorylases
3.4.3. Trehalose Synthases
Trehalose 6-Phosphate Synthases
3.4.4. Mannosyl-3-Phosphoglycerate (MPG) Synthases
4. Conclusions
References
- Pfeifer, K.; Ergal, İ.; Koller, M.; Basen, M.; Schuster, B.; Rittmann, S.K.R. Archaea Biotechnology. Biotechnol. Adv. 2021, 47, 107668.
- Zeldes, B.M.; Keller, M.W.; Loder, A.J.; Straub, C.T.; Adams, M.W.; Kelly, R.M. Extremely thermophilic microorganisms as metabolic engineering platforms for production of fuels and industrial chemicals. Front. Microbiol. 2015, 6, 1209.
- Park, S.; Lee, B.; Park, K. Extremophilic Carbohydrate Active Enzymes (CAZymes). J. Nutr. Health Food Eng. 2017, 7, 230–237.
- Dumorné, K.; Córdova, D.C.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A Potential Source for Industrial Applications. J. Microbiol. Biotechnol. 2017, 27, 649–659.
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic. Acids Res. 2009, 37, D233–D238.
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic. Acids Res. 2014, 42, D490–D495.
- Gloster, T.M. Advances in understanding glycosyltransferases from a structural perspective. Curr. Opin. Struct. Biol. 2014, 28, 131–141.
- Cobucci-Ponzano, B.; Rossi, M.; Moracci, M. Extremophiles Handbook; Springer: Tokyo, Japan, 2011; pp. 427–441.
- Cabrera, M.; Blamey, J.M. Biotechnological applications of archaeal enzymes from extreme environments. Biol. Res. 2018, 51, 37.
- Ebaid, R.; Wang, H.; Sha, C.; Abomohra, A.E.-F.; Shao, W. Recent trends in hyperthermophilic enzymes production and future perspectives for biofuel industry: A critical review. J. Clean. Prod. 2019, 238, 117925.
- Straub, C.T.; Counts, J.A.; Nguyen, D.M.N.; Wu, C.H.; Zeldes, B.M.; Crosby, J.R.; Conway, J.M.; Otten, J.K.; Lipscomb, G.L.; Schut, G.J.; et al. Biotechnology of extremely thermophilic archaea. FEMS Microbiol. Rev. 2018, 42, 543–578.
- Davies, G.; Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 1995, 3, 853–859.
- Naumoff, D. Hierarchical classification of glycoside hydrolases. Biochemistry (Moscow) 2011, 76, 622–635.
- Ati, J.; Lafite, P.; Daniellou, R. Enzymatic synthesis of glycosides: From natural O- and N-glycosides to rare C- and S-glycosides. Beilstein. J. Org. Chem. 2017, 13, 1857–1865.
- Sjogren, J.; Collin, M. Bacterial glycosidases in pathogenesis and glycoengineering. Future Microbiol. 2014, 9, 1039–1051.
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell Wall Hydrolases in Bacteria: Insight on the Diversity of Cell Wall Amidases, Glycosidases and Peptidases Toward Peptidoglycan. Front. Microbiol. 2019, 10, 331.
- Henrissat, B.; Romeu, A. Families, superfamilies and subfamilies of glycosyl hydrolases. Biochem. J. 1995, 311, 350–351.
- Vuong, T.V.; Wilson, D.B. Glycoside hydrolases: Catalytic base/nucleophile diversity. Biotechnol. Bioeng. 2010, 107, 195–205.
- McCarter, J.D.; Withers, S.G. Mechanisms of enzymatic glycoside hydrolysis. Curr. Opin. Struct. Biol. 1994, 4, 885–892.
- Carbohydrate Active Enzymes Database. Available online: http://www.cazy.org/ (accessed on 1 July 2021).
- Strazzulli, A.; Cobucci-Ponzano, B.; Iacono, R.; Giglio, R.; Maurelli, L.; Curci, N.; Schiano-di-Cola, C.; Santangelo, A.; Contursi, P.; Lombard, V.; et al. Discovery of hyperstable carbohydrate-active enzymes through metagenomics of extreme environments. FEBS J. 2020, 287, 1116–1137.
- Iacono, R.; Cobucci-Ponzano, B. Spatial Metagenomics of Three Geothermal Sites in Pisciarelli Hot Spring Focusing on the Biochemical Resources of the Microbial Consortia. Molecules 2020, 25, 4023.
- Sharma, N.; Kumar, J.; Abedin, M.; Sahoo, D.; Pandey, A.; Rai, A.K.; Singh, S.P. Metagenomics revealing molecular profiling of community structure and metabolic pathways in natural hot springs of the Sikkim Himalaya. BMC Microbiol. 2020, 20, 1–17.
- Dadwal, A.; Sharma, S.; Satyanarayana, T. Thermostable cellulose saccharifying microbial enzymes: Characteristics, recent advances and biotechnological applications. Int. J. Biol. Macromol. 2021, 188, 226–244.
- Suleiman, M.; Krüger, A.; Antranikian, G. Biomass-degrading glycoside hydrolases of archaeal origin. Biotechnol. Biofuels 2020, 13, 1–14.
- Koch, R.; Spreinat, A.; Lemke, K.; Antranikian, G. Purification and properties of a hyperthermoactive α-amylase from the archaeobacterium Pyrococcus woesei. Arch. Microbiol. 1991, 155, 572–578.
- Andrade, C.M.M.C.; Pereira, N., Jr.; Antranikian, G. Extremely thermophilic microorganisms and their polymer-hidrolytic enzymes. Rev. Microbiol. 1999, 30, 287–298.
- Schiraldi, C.; Giuliano, M.; De Rosa, M. Perspectives on biotechnological applications of archaea. Archaea 2002, 1, 75–86.
- Burg, B.V.D. Extremophiles as a source for novel enzymes. Curr. Opin. Microbiol. 2003, 6, 213–218.
- Desmet, T.; Soetaert, W. Enzymatic glycosyl transfer: Mechanisms and applications. Biocatal. Biotransformation 2011, 29, 1–18.
- Yennamalli, R.M.; Rader, A.J.; Kenny, A.J.; Wolt, J.D.; Sen, T.Z. Endoglucanases: Insights into thermostability for biofuel applications. Biotechnol. Biofuels 2013, 6, 136.
- Leemhuis, H.; Dijkhuizen, L. Hydrolysis and Transglycosylation Reaction Specificity of Cyclodextrin Glycosyltransferases. J. Appl. Glycosci. 2003, 50, 263–271.
- Torres, D.P.; Gonçalves, M.D.P.F.; Teixeira, J.A.; Rodrigues, L.R. Galacto-Oligosaccharides: Production, Properties, Applications, and Significance as Prebiotics. Compr. Rev. Food Sci. Food Saf. 2010, 9, 438–454.
- Velázquez, J.B.; Villa, T.G. Industrial Applications of Hyperthermophilic Enzymes: A Review. Protein Pept. Lett. 2006, 13, 645–651.
- Egorova, K.; Antranikian, G. Industrial relevance of thermophilic Archaea. Curr. Opin. Microbiol. 2005, 8, 649–655.
- Henrissat, B.; Bairoch, A. Updating the sequence-based classification of glycosyl hydrolases. Biochem. J. 1996, 316, 695–696.
- Li, X.; Zhao, J.; Fu, J.; Pan, Y.; Li, D. Sequence analysis and biochemical properties of an acidophilic and hyperthermophilic amylopullulanase from Thermofilum pendens. Int. J. Biol. Macromol. 2018, 114, 235–243.
- Kim, M.-S.; Park, J.-T.; Kim, Y.-W.; Lee, H.-S.; Nyawira, R.; Shin, H.-S.; Park, C.-S.; Yoo, S.-H.; Kim, Y.-R.; Moon, T.-W.; et al. Properties of a Novel Thermostable Glucoamylase from the Hyperthermophilic Archaeon Sulfolobus solfataricus in Relation to Starch Processing. Appl. Environ. Microbiol. 2004, 70, 3933–3940.
- Jung, J.-H.; Seo, D.-H.; Holden, J.F.; Park, C.-S. Maltose-forming α-amylase from the hyperthermophilic archaeon Pyrococcus sp. ST04. Appl. Microbiol. Biotechnol. 2014, 98, 2121–2131.
- Ghasemi, A.; Ghafourian, S.; Vafaei, S.; Mohebi, R.; Farzi, M.; Taherikalani, M.; Sadeghifard, N. Cloning, Expression, and Purification of Hyperthermophile α-Amylase from Pyrococcus woesei. Osong. Public. Health Res. Perspect. 2015, 6, 336–340.
- Wang, S.; Lu, Z.; Lu, M.; Qin, S.; Liu, H.; Deng, X.; Lin, Q.; Chen, J. Identification of archaeon-producing hyperthermophilic α-amylase and characterization of the α-amylase. Appl. Microbiol. Biotechnol. 2008, 80, 605–614.
- Ying, Q.; Zhang, C.; Guo, F.; Wang, S.; Bie, X.; Lu, F.; Lu, Z. Secreted Expression of a Hyperthermophilic α-Amylase Gene from Thermococcus sp. HJ21 in Bacillus subtilis. J. Mol. Microbiol. Biotechnol. 2012, 22, 392–398.
- Kwak, Y.S.; Akiba, T.; Kudo, T. Purification and characterization of α-amylase from hyperthermophilic archaeon Thermococcus profundus, which hydrolyzes both α-1,4 and α-1,6 glucosidic linkages. J. Ferment. Bioeng. 1998, 86, 363–367.
- Chung, Y.C.; Kobayashi, T.; Kanai, H.; Akiba, T.; Kudo, T. Purification and Properties of Extracellular Amylase from the Hyperthermophilic Archaeon Thermococcus profundus DT5432. Appl. Environ. Microbiol. 1995, 61, 1502–1506.
- Haseltine, C.; Rolfsmeier, M.; Blum, P. The glucose effect and regulation of alpha-amylase synthesis in the hyperthermophilic archaeon Sulfolobus solfataricus. J. Bacteriol. 1996, 178, 945–950.
- Kim, J.W.; Flowers, L.O.; Whiteley, M.; Peeples, T.L. Biochemical confirmation and characterization of the family-57-like alpha-amylase of Methanococcus jannaschii. Folia Microbiol. (Praha) 2001, 46, 467–473.
- Savchenko, A.; Vieille, C.; Kang, S.; Zeikus, J.G. Pyrococcus furiosus alpha-amylase is stabilized by calcium and zinc. Biochemistry 2002, 41, 6193–6201.
- Jung, T.; Li, D.; Park, J.-T.; Yoon, S.-M.; Tran, P.L.; Oh, B.-H.; Janeček, S.; Park, S.G.; Woo, E.-J.; Park, K.-H. Association of Novel Domain in Active Site of Archaic Hyperthermophilic Maltogenic Amylase from Staphylothermus marinus. J. Biol. Chem. 2012, 287, 7979–7989.
- Li, X.; Wang, Y.; Park, J.T.; Gu, L.; Li, D. An extremely thermostable maltogenic amylase from Staphylothermus marinus: Bacillus expression of the gene and its application in genistin glycosylation. Int. J. Biol. Macromol. 2018, 107, 413–417.
- Li, D.; Park, J.-T.; Li, X.; Kim, S.; Lee, S.; Shim, J.-H.; Park, S.-H.; Cha, J.; Lee, B.; Kim, J.-W.; et al. Overexpression and characterization of an extremely thermostable maltogenic amylase, with an optimal temperature of 100°C, from the hyperthermophilic archaeon Staphylothermus marinus. New Biotechnol. 2010, 27, 300–307.
- Kim, J.-W.; Kim, Y.-H.; Lee, H.-S.; Yang, S.-J.; Kim, Y.-W.; Lee, M.-H.; Seo, N.-S.; Park, C.-S.; Park, K.-H. Molecular cloning and biochemical characterization of the first archaeal maltogenic amylase from the hyperthermophilic archaeon Thermoplasma volcanium GSS1. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2007, 1774, 661–669.
- Satyanarayana, T.; Noorwez, S.; Kumar, S.; Rao, J.; Ezhilvannan, M.; Kaur, P. Development of an ideal starch saccharification process using amylolytic enzymes from thermophiles. Biochem. Soc. Trans. 2004, 32, 276–278.
- Comfort, D.A.; Chou, C.J.; Conners, S.B.; VanFossen, A.L.; Kelly, R.M. Functional-genomics-based identification and characterization of open reading frames encoding alpha-glucoside-processing enzymes in the hyperthermophilic archaeon Pyrococcus furiosus. Appl. Environ. Microbiol. 2008, 74, 1281–1283.
- Serour, E.; Antranikian, G. Novel thermoactive glucoamylases from the thermoacidophilic Archaea Thermoplasma acidophilum, Picrophilus torridus and Picrophilus oshimae. Antonie. Van. Leeuwenhoek. 2002, 81, 73–83.
- Dock, C.; Hess, M.; Antranikian, G. A thermoactive glucoamylase with biotechnological relevance from the thermoacidophilic Euryarchaeon Thermoplasma acidophilum. Appl. Microbiol. Biotechnol. 2007, 78, 105–114.
- Schepers, B.; Thiemann, V.; Antranikian, G. Characterization of a Novel Glucoamylase from the Thermoacidophilic ArchaeonPicrophilus torridus Heterologously Expressed inE. coli. Eng. Life Sci. 2006, 6, 311–317.
- Satyanarayana, T.; Nisha, M. Archaeal and bacterial thermostable amylopullulanases: Characteristic features and biotechnological applications. Amylase 2018, 2, 44.
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Bin Ariff, A. Pullulanase: Role in Starch Hydrolysis and Potential Industrial Applications. Enzym. Res. 2012, 2012, 1–14.
- Saha, B.C.; Zeikus, J.G. Novel highly thermostable pullulanase from thermophiles. Trends Biotechnol. 1989, 7, 234–239.
- Brown, S.H.; Costantino, H.R.; Kelly, R.M. Characterization of Amylolytic Enzyme Activities Associated with the Hyperthermophilic Archaebacterium Pyrococcus furiosus. Appl. Environ. Microbiol. 1990, 56, 1985–1991.
- Brown, S.H.; Kelly, R.M. Characterization of Amylolytic Enzymes, Having Both alpha-1,4 and alpha-1,6 Hydrolytic Activity, from the Thermophilic Archaea Pyrococcus furiosus and Thermococcus litoralis. Appl. Environ. Microbiol. 1993, 59, 2614–2621.
- Pan, S.; Ding, N.; Ren, J.; Gu, Z.; Li, C.; Hong, Y.; Cheng, L.; Holler, T.P.; Li, Z. Maltooligosaccharide-forming amylase: Characteristics, preparation, and application. Biotechnol. Adv. 2017, 35, 619–632.
- Ben Ali, M.; Khemakhem, B.; Robert, X.; Haser, R.; Bejar, S. Thermostability enhancement and change in starch hydrolysis profile of the maltohexaose-forming amylase of Bacillus stearothermophilus US100 strain. Biochem. J. 2006, 394, 51–56.
- Cheng, K.; Zhang, F.; Sun, F.; Chen, H.; Zhang, Y.-H.P. Doubling Power Output of Starch Biobattery Treated by the Most Thermostable Isoamylase from an Archaeon Sulfolobus tokodaii. Sci. Rep. 2015, 5, 13184.
- Van der Maarel, M.J.; van der Veen, B.; Uitdehaag, J.C.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol. 2002, 94, 137–155.
- Li, X.; Li, D.; Park, K.-H. An extremely thermostable amylopullulanase from Staphylothermus marinus displays both pullulan- and cyclodextrin-degrading activities. Appl. Microbiol. Biotechnol. 2012, 97, 5359–5369.
- Li, Y.; Xu, J.; Zhang, L.; Ding, Z.; Gu, Z.; Shi, G. Investigation of debranching pattern of a thermostable isoamylase and its application for the production of resistant starch. Carbohydr. Res. 2017, 446–447, 93–100.
- Costantino, H.; Brown, S.; Kelly, R. Purification and characterization of an α-glucosidase from a hyperthermophilic archaebacterium, Pyrococcus furiosus, exhibiting a temperature optimum of 105 to 115°C. J. Bacteriol. 1990, 172, 3654–3660.
- Rolfsmeier, M.; Blum, P. Purification and characterization of a maltase from the extremely thermophilic crenarchaeote Sulfolobus solfataricus. J. Bacteriol. 1995, 177, 482–485.
- Rolfsmeier, M.; Haseltine, C.; Bini, E.; Clark, A.; Blum, P. Molecular Characterization of the α-Glucosidase Gene from the Hyperthermophilic Archaeon Sulfolobus solfataricus. J. Bacteriol. 1998, 180, 1287–1295.
- Chang, S.T.; Parker, K.N.; Bauer, M.W.; Kelly, R.M. Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2001; pp. 260–269.
- Lee, M.-H.; Yang, S.-J.; Kim, J.-W.; Lee, H.-S.; Kim, J.-W.; Park, K.-H. Characterization of a thermostable cyclodextrin glucanotransferase from Pyrococcus furiosus DSM3638. Extremophiles 2007, 11, 537–541.
- Rashid, N.; Cornista, J.; Ezaki, S.; Fukui, T.; Atomi, H.; Imanaka, T. Characterization of an archaeal cyclodextrin glucanotransferase with a novel C-terminal domain. J. Bacteriol. 2002, 184, 777–784.
- Park, H.-S.; Park, J.-T.; Kang, H.-K.; Cha, H.; Kim, D.-S.; Kim, J.-W.; Park, K.-H. TreX fromSulfolobus solfataricusATCC 35092 Displays Isoamylase and 4-α-Glucanotransferase Activities. Biosci. Biotechnol. Biochem. 2007, 71, 1348–1352.
- Xavier, K.B.; Peist, R.; Kossmann, M.; Boos, W.; Santos, H. Maltose metabolism in the hyperthermophilic archaeon Thermococcus litoralis: Purification and characterization of key enzymes. J. Bacteriol. 1999, 181, 3358–3367.
- Kaper, T.; Talik, B.; Ettema, T.J.; Bos, H.; van der Maarel, M.J.E.C.; Dijkhuizen, L. Amylomaltase of Pyrobaculum aerophilum IM2 Produces Thermoreversible Starch Gels. Appl. Environ. Microbiol. 2005, 71, 5098–5106.
- Mehboob, S.; Ahmad, N.; Rashid, N.; Imanaka, T.; Akhtar, M. Pcal_0768, a hyperactive 4-α-glucanotransferase from Pyrobacculum calidifontis. Extremophiles 2016, 20, 559–566.
- Mehboob, S.; Ahmad, N.; Munir, S.; Ali, R.; Younas, H.; Rashid, N. Gene cloning, expression enhancement in Escherichia coli and biochemical characterization of a highly thermostable amylomaltase from Pyrobaculum calidifontis. Int. J. Biol. Macromol. 2020, 165, 645–653.
- Patel, A.K.; Singhania, R.R.; Sim, S.J.; Pandey, A. Thermostable cellulases: Current status and perspectives. Bioresour. Technol. 2019, 279, 385–392.
- Pisani, F.M.; Rella, R.; Raia, C.A.; Rozzo, C.; Nucci, R.; Gambacorta, A.; de Rosa, M.; Rossi, M. Thermostable beta-galactosidase from the archaebacterium Sulfolobus solfataricus Purification and properties. JBIC J. Biol. Inorg. Chem. 1990, 187, 321–328.
- Vieille, C.; Zeikus, G.J. Hyperthermophilic Enzymes: Sources, Uses, and Molecular Mechanisms for Thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43.
- Rigoldi, F.; Donini, S.; Redaelli, A.; Parisini, E.; Gautieri, A. Review: Engineering of thermostable enzymes for industrial applications. Appl. Bioeng. 2018, 2, 011501.
- Sharma, S.; Vaid, S.; Bhat, B.; Singh, S.; Bajaj, B.K. Biomass, Biofuels and Biochemicals: Advances in Enzyme Technology; Elsevier: Jammu, India, 2019; Volume 17, pp. 469–495.
- Turner, P.; Mamo, G.; Karlsson, E.N. Potential and utilization of thermophiles and thermostable enzymes in biorefining. Microb. Cell Factories 2007, 6, 9.
- Singhal, G.; Bhagyawant, S.S.; Srivastava, N. Current Status and Future Scope of Microbial Cellulases; Elsevier: Jammu, India, 2021; Volume 3, pp. 39–57.
- Kashima, Y.; Mori, K.; Fukada, H.; Ishikawa, K. Analysis of the function of a hyperthermophilic endoglucanase from Pyrococcus horikoshii that hydrolyzes crystalline cellulose. Extremophiles 2005, 9, 37–43.
- Bauer, M.W.; Driskill, L.E.; Callen, W.; Snead, M.A.; Mathur, E.J.; Kelly, R.M. An Endoglucanase, EglA, from the Hyperthermophilic Archaeon Pyrococcus furiosus Hydrolyzes β-1,4 Bonds in Mixed-Linkage (1→3),(1→4)-β-D-Glucans and Cellulose. J. Bacteriol. 1999, 181, 284–290.
- Neelamegam, A.; Rajeswari, M.; Thangavel, B.; Gupta, V. Microbial Enzymes in Bioconversions of Biomass; Springer: Cham, Switzerland, 2016; pp. 37–45.
- Girfoglio, M.; Rossi, M.; Cannio, R. Cellulose Degradation by Sulfolobus solfataricus Requires a Cell-Anchored Endo-β-1-4-Glucanase. J. Bacteriol. 2012, 194, 5091–5100.
- Klose, H.; Röder, J.; Girfoglio, M.; Fischer, R.; Commandeur, U. Hyperthermophilic endoglucanase for in planta lignocellulose conversion. Biotechnol. Biofuels 2012, 5, 63.
- Zhang, X.-Z.; Zhang, Y.-H.P. Bioprocessing Technologies in Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 131–146.
- Béguin, P. Molecular biology of cellulose degradation. Annu. Rev. Microbiol. 1990, 44, 219–248.
- Tipparat, H. Hyperthermostable cellulolytic and hemicellulolytic enzymes and their biotechnological applications. SJST 2002, 24, 481–491.
- Olajuyigbe, F.M.; Nlekerem, C.M.; Ogunyewo, O.A. Production and Characterization of Highly Thermostable β-Glucosidase during the Biodegradation of Methyl Cellulose by Fusarium oxysporum. Biochem. Res. Int. 2016, 2016, 3978124.
- Li, D.; Li, X.; Dang, W.; Tran, P.L.; Park, S.H.; Oh, B.C.; Hong, W.S.; Lee, J.S.; Park, K.H. Characterization and application of an acidophilic and thermostable beta-glucosidase from Thermofilum pendens. J. Biosci. Bioeng. 2013, 115, 490–496.
- Hansson, T.; Kaper, T.; van Der Oost, J.; de Vos, W.M.; Adlercreutz, P. Improved oligosaccharide synthesis by protein engineering of beta-glucosidase CelB from hyperthermophilic Pyrococcus furiosus. Biotechnol. Bioeng. 2001, 73, 203–210.
- Voorhorst, W.G.; Eggen, I.R.; Luesink, E.J.; de Vos, W.M. Characterization of the celB gene coding for beta-glucosidase from the hyperthermophilic archaeon Pyrococcus furiosus and its expression and site-directed mutation in Escherichia coli. J. Bacteriol. 1995, 177, 7105–7111.
- Kado, Y.; Inoue, T.; Ishikawa, K. Structure of hyperthermophilic β-glucosidase from Pyrococcus furiosus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1473–1479.
- Lebbink, J.; Kaper, T.; Kengen, S.; Oost, J.; De Vos, W. β-glucosidase CelB from Pyrococcus furiosus: Production by Escherichia coli, purification, and in vitro evolution. Methods Enzym. 2001, 330, 364–379.
- Kim, H.W.; Ishikawa, K. Complete saccharification of cellulose at high temperature using endocellulase and beta-glucosidase from Pyrococcus sp. J. Microbiol. Biotechnol. 2010, 20, 889–892.
- Park, S.H.; Park, K.H.; Oh, B.C.; Alli, I.; Lee, B.H. Expression and characterization of an extremely thermostable beta-glycosidase (mannosidase) from the hyperthermophilic archaeon Pyrococcus furiosus DSM3638. N Biotechnol. 2011, 28, 639–648.
- Gumerov, V.; Rakitin, A.; Mardanov, A.; Ravin, N.V. A Novel Highly Thermostable Multifunctional Beta-Glycosidase from CrenarchaeonAcidilobus saccharovorans. Archaea 2015, 2015, 1–6.
- Boyce, A.; Walsh, G. Expression and characterisation of a thermophilic endo-1,4-β-glucanase from Sulfolobus shibatae of potential industrial application. Mol. Biol. Rep. 2018, 45, 2201–2211.
- Park, N.-Y.; Cha, J.; Kim, D.-O.; Park, C.-S. Enzymatic characterization and substrate specificity of thermostable beta-glycosidase from hyperthermophilic archaea, Sulfolobus shibatae, expressed in E. coli. J. Microbiol. Biotechnol. 2007, 17, 454–460.
- Rathore, A.S.; Gupta, R.D. Chitinases from Bacteria to Human: Properties, Applications, and Future Perspectives. Enzym. Res. 2015, 2015, 791907.
- Oyeleye, A.; Normi, Y.M. Chitinase: Diversity, limitations, and trends in engineering for suitable applications. Biosci. Rep. 2018, 38.
- Chen, L.; Wei, Y.; Shi, M.; Li, Z.; Zhang, S.-H. An Archaeal Chitinase With a Secondary Capacity for Catalyzing Cellulose and Its Biotechnological Applications in Shell and Straw Degradation. Front. Microbiol. 2019, 10, 1253.
- Le, B.; Yang, S.H. Microbial chitinases: Properties, current state and biotechnological applications. World J. Microbiol. Biotechnol. 2019, 35, 144.
- Kuzu, S.B.; Güvenmez, H.K.; Denizci, A.A. Production of a Thermostable and Alkaline Chitinase by Bacillus thuringiensis subsp. kurstaki Strain HBK-51. Biotechnol. Res. Int. 2012, 2012, 1–6.
- Das, S.; Chilukoti, N.; Pvsrn, S.; Jogi, M.; Pallinti, P.N.; Kaur, M.; Dutta, S.; Podile, A.R. Microorganisms in Environmental Management: Microbes and Environment; Elsevier: Dordrecht, The Netherlands, 2012; pp. 135–150.
- Tanaka, T.; Fujiwara, S.; Nishikori, S.; Fukui, T.; Takagi, M.; Imanaka, T. A Unique Chitinase with Dual Active Sites and Triple Substrate Binding Sites from the Hyperthermophilic Archaeon Pyrococcus kodakaraensis KOD1. Appl. Environ. Microbiol. 1999, 65, 5338–5344.
- Tanaka, T.; Fukui, T.; Imanaka, T. Different cleavage specificities of the dual catalytic domains in chitinase from the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1. J. Biol. Chem. 2001, 276, 35629–35635.
- Nakamura, T.; Mine, S.; Hagihara, Y.; Ishikawa, K.; Uegaki, K. Structure of the catalytic domain of the hyperthermophilic chitinase fromPyrococcus furiosus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 63, 7–11.
- Chettri, D.; Verma, A.K.; Verma, A.K. Innovations in CAZyme gene diversity and its modification for biorefinery applications. Biotechnol. Rep. 2020, 28, e00525.
- Graham, J.E.; Clark, M.E.; Nadler, D.C.; Huffer, S.; Chokhawala, H.A.; Rowland, S.E.; Blanch, H.W.; Clark, D.S.; Robb, F.T. Identification and characterization of a multidomain hyperthermophilic cellulase from an archaeal enrichment. Nat. Commun. 2011, 2, 375.
- Gavrilov, S.N.; Stracke, C.; Jensen, K.; Menzel, P.; Kallnik, V.; Slesarev, A.; Sokolova, T.; Zayulina, K.; Bräsen, C.; Bonch-Osmolovskaya, E.A.; et al. Isolation and Characterization of the First Xylanolytic Hyperthermophilic Euryarchaeon Thermococcus sp. Strain 2319x1 and Its Unusual Multidomain Glycosidase. Front. Microbiol. 2016, 7, 552.
- Antranikian, G.; Suleiman, M.; Schäfers, C.; Adams, M.W.W.; Bartolucci, S.; Blamey, J.M.; Birkeland, N.K.; Bonch-Osmolovskaya, E.; da Costa, M.S.; Cowan, D.; et al. Diversity of bacteria and archaea from two shallow marine hydrothermal vents from Vulcano Island. Extremophiles 2017, 21, 733–742.
- Schröder, C.; Eixenberger, D.; Suleiman, M.; Schäfers, C.; Antranikian, G. Characterization of an extremely thermo-active archaeal β-glucosidase and its activity towards glucan and mannan in concert with an endoglucanase. Appl. Microbiol. Biotechnol. 2019, 103, 9505–9514.
- Lewin, A.; Zhou, J.; Pham, V.T.T.; Haugen, T.; Zeiny, M.E.; Aarstad, O.; Liebl, W.; Wentzel, A.; Liles, M.R. Novel archaeal thermostable cellulases from an oil reservoir metagenome. AMB Express. 2017, 7, 183.
- Kaushal, G.; Kumar, J.; Sangwan, R.S.; Singh, S.P. Metagenomic analysis of geothermal water reservoir sites exploring carbohydrate-related thermozymes. Int. J. Biol. Macromol. 2018, 119, 882–895.
- Kapitonov, D.; Yu, R.K. Conserved domains of glycosyltransferases. Glycobiology 1999, 9, 961–978.
- Schmid, J.; Heider, D.; Wendel, N.J.; Esperl, N.; Sieber, V. Bacterial Glycosyltransferases: Challenges and Opportunities of a Highly Diverse Enzyme Class Toward Tailoring Natural Products. Front. Microbiol. 2016, 7, 182.
- Breton, C.; Šnajdrová, L.; Jeanneau, C.; Koca, J.; Imberty, A. Structures and mechanisms of glycosyltransferases. Glycobiology 2005, 16, 29R–37R.
- Liu, J.; Mushegian, A. Three monophyletic superfamilies account for the majority of the known glycosyltransferases. Protein Sci. 2003, 12, 1418–1431.
- Sinnott, M.L. Catalytic mechanism of enzymic glycosyl transfer. Chem. Rev. 1990, 90, 1171–1202.
- Liang, D.-M.; Liu, J.-H.; Wu, H.; Wang, B.-B.; Zhu, H.-J.; Qiao, J.-J. Glycosyltransferases: Mechanisms and applications in natural product development. Chem. Soc. Rev. 2015, 44, 8350–8374.
- Protein Data Bank PDB. Available online: https://www.rcsb.org/ (accessed on 1 October 2021).
- Coutinho, P.M.; Deleury, E.; Davies, G.; Henrissat, B. An Evolving Hierarchical Family Classification for Glycosyltransferases. J. Mol. Biol. 2003, 328, 307–317.
- Fukuda, M.; Bierhuizen, M.F.A.; Nakayama, J. Expression cloning of glycosyltransferases. Glycobiology 1996, 6, 683–689.
- Matsumoto, S.; Shimada, A.; Nyirenda, J.; Igura, M.; Kawano, Y.; Kohda, D. Crystal structures of an archaeal oligosaccharyltransferase provide insights into the catalytic cycle of N-linked protein glycosylation. Proc. Natl. Acad. Sci. USA 2013, 110, 17868–17873.
- Shrimal, S.; Gilmore, R. Oligosaccharyltransferase structures provide novel insight into the mechanism of asparagine-linked glycosylation in prokaryotic and eukaryotic cells. Glycobiology 2018, 29, 288–297.
- Jarrell, K.F.; Ding, Y.; Meyer, B.H.; Albers, S.-V.; Kaminski, L.; Eichler, J. N-Linked Glycosylation in Archaea: A Structural, Functional, and Genetic Analysis. Microbiol. Mol. Biol. Rev. 2014, 78, 304–341.
- Abu-Qarn, M.; Giordano, A.; Battaglia, F.; Trauner, A.; Hitchen, P.G.; Morris, H.R.; Dell, A.; Eichler, J. Identification of AglE, a Second Glycosyltransferase Involved in N Glycosylation of the Haloferax volcanii S-Layer Glycoprotein. J. Bacteriol. 2008, 190, 3140–3146.
- VanDyke, D.J.; Wu, J.; Logan, S.M.; Kelly, J.F.; Mizuno, S.; Aizawa, S.; Jarrell, K.F. Identification of genes involved in the assembly and attachment of a novel flagellin N-linked tetrasaccharide important for motility in the archaeon Methanococcus maripaludis. Mol. Microbiol. 2009, 72, 633–644.
- Meyer, B.H.; Peyfoon, E.; Dietrich, C.; Hitchen, P.; Panico, M.; Morris, H.R.; Dell, A.; Albers, S.-V. Agl16, a Thermophilic Glycosyltransferase Mediating the Last Step of N-Glycan Biosynthesis in the Thermoacidophilic Crenarchaeon Sulfolobus acidocaldarius. J. Bacteriol. 2013, 195, 2177.
- Elharar, Y.; Podilapu, A.R.; Guan, Z.; Kulkarni, S.S.; Eichler, J. Assembling Glycan-Charged Dolichol Phosphates: Chemoenzymatic Synthesis of a Haloferax volcanii N-Glycosylation Pathway Intermediate. Bioconjugate Chem. 2017, 28, 2461–2470.
- Igura, M.; Maita, N.; Kamishikiryo, J.; Yamada, M.; Obita, T.; Maenaka, K.; Kohda, D. Structure-guided identification of a new catalytic motif of oligosaccharyltransferase. EMBO J. 2007, 27, 234–243.
- Kohda, D. Structural Basis of Protein Asn-Glycosylation by Oligosaccharyltransferases. Adv. Exp. Med. Biol 2018, 1104, 171–199.
- Magidovich, H.; Eichler, J. Glycosyltransferases and oligosaccharyltransferases in Archaea: Putative components of the N-glycosylation pathway in the third domain of life. Fems. Microbiol. Lett. 2009, 300, 122–130.
- Cohen-Rosenzweig, C.; Guan, Z.; Shaanan, B.; Eichler, J. Substrate Promiscuity: AglB, the Archaeal Oligosaccharyltransferase, Can Process a Variety of Lipid-Linked Glycans. Appl. Environ. Microbiol. 2013, 80, 486–496.
- Zhu, B.C.; Laine, R.A. Dolichyl-phosphomannose synthase from the Archae Thermoplasma acidophilum. Glycobiology 1996, 6, 811–816.
- VanFossen, A.L.; Lewis, D.L.; Nichols, J.D.; Kelly, R.M. Polysaccharide Degradation and Synthesis by Extremely Thermophilic Anaerobes. Ann. N. Y. Acad. Sci. 2008, 1125, 322–337.
- Urushibata, Y.; Ebisu, S.; Matsui, I. A thermostable dolichol phosphoryl mannose synthase responsible for glycoconjugate synthesis of the hyperthermophilic archaeon Pyrococcus horikoshii. Extremophiles 2008, 12, 665–676.
- Takemasa, R.; Yokooji, Y.; Yamatsu, A.; Atomi, H.; Imanaka, T. Thermococcus kodakarensis as a Host for Gene Expression and Protein Secretion. Appl. Environ. Microbiol. 2011, 77, 2392–2398.
- Martin, D.D.; Ciulla, R.A.; Roberts, M.F. Osmoadaptation in Archaea. Appl. Environ. Microbiol. 1999, 65, 1815–1825.
- Kelly, R.; Dijkhuizen, L.; Leemhuis, H. Starch and α-glucan acting enzymes, modulating their properties by directed evolution. J. Biotechnol. 2009, 140, 184–193.
- Ubiparip, Z.; Beerens, K.; Franceus, J.; Vercauteren, R.; Desmet, T. Thermostable alpha-glucan phosphorylases: Characteristics and industrial applications. Appl. Microbiol. Biotechnol. 2018, 102, 8187–8202.
- Gao, Y.; Jiang, Y.; Liu, Q.; Wang, R.; Liu, X.; Liu, B. Enzymatic and regulatory properties of the trehalose-6-phosphate synthase from the thermoacidophilic archaeon Thermoplasma acidophilum. Biochimie 2014, 101, 215–220.
- Wang, J.-H.; Tsai, M.-Y.; Lee, G.-C.; Shaw, J.-F. Construction of a Recombinant Thermostable β-Amylase-Trehalose Synthase Bifunctional Enzyme for Facilitating the Conversion of Starch to Trehalose. J. Agric. Food Chem. 2007, 55, 1256–1263.
- Empadinhas, N.; Marugg, J.; Borges, N.; Santos, H.; da Costa, M. Pathway for the Synthesis of Mannosylglycerate in the Hyperthermophilic Archaeon Pyrococcus horikoshii. J. Biol. Chem. 2001, 276, 43580–43588.
- Horcajada, C.; Guinovart, J.J.; Fita, I.; Ferrer, J.C. Crystal structure of an archaeal glycogen synthase: Insights into oligomerization and substrate binding of eukaryotic glycogen synthases. J. Biol. Chem. 2006, 281, 2923–2931.
- Gruyer, S.; Legin, E.; Bliard, C.; Ball, S.; Duchiron, F. The Endopolysaccharide Metabolism of the Hyperthermophilic Archeon Thermococcus hydrothermalis: Polymer Structure and Biosynthesis. Curr. Microbiol. 2002, 44, 206–211.
- Zea, C.J.; MacDonell, S.W.; Pohl, N.L. Discovery of the Archaeal Chemical Link between Glycogen (Starch) Synthase Families Using a New Mass Spectrometry Assay. J. Am. Chem. Soc. 2003, 125, 13666–13667.
- Zea, C.J.; Pohl, N.L. Unusual sugar nucleotide recognition elements of mesophilic vs. thermophilic glycogen synthases. Biopolymers 2005, 79, 106–113.
- Mizanur, R.M.; Griffin, A.K.K.; Pohl, N.L. Recombinant production and biochemical characterization of a hyperthermostable α-glucan/maltodextrin phosphorylase fromPyrococcus furiosus. Archaea 2008, 2, 169–176.
- Rathore, R.S.; Garg, N.; Garg, S.; Kumar, A. Starch phosphorylase: Role in starch metabolism and biotechnological applications. Crit. Rev. Biotechnol. 2009, 29, 214–224.
- Zhu, Z.; Kin Tam, T.; Sun, F.; You, C.; Percival Zhang, Y.H. A high-energy-density sugar biobattery based on a synthetic enzymatic pathway. Nat. Commun. 2014, 5, 3026.
- Labes, A.; Schönheit, P. Unusual Starch Degradation Pathway via Cyclodextrins in the Hyperthermophilic Sulfate-Reducing Archaeon Archaeoglobus fulgidus Strain 7324. J. Bacteriol. 2007, 189, 8901–8913.
- Mueller, M.; Takemasa, R.; Schwarz, A.; Atomi, H.; Nidetzky, B. “Short-chain” α-1,4-glucan phosphorylase having a truncated N-terminal domain: Functional expression and characterization of the enzyme from Sulfolobus solfataricus. Biochim. Biophys. Acta 2009, 1794, 1709–1714.
- Ohtake, S.; Wang, Y.J. Trehalose: Current use and future applications. J. Pharm. Sci. 2011, 100, 2020–2053.
- Kim, H.-H.; Jung, J.-H.; Seo, D.-H.; Ha, S.-J.; Yoo, S.-H.; Kim, C.-H.; Park, C.-S. Novel enzymatic production of trehalose from sucrose using amylosucrase and maltooligosyltrehalose synthase-trehalohydrolase. World J. Microbiol. Biotechnol. 2011, 27, 2851–2856.
- Ryu, S.-I.; Park, C.-S.; Cha, J.; Woo, E.-J.; Lee, S.-B. A novel trehalose-synthesizing glycosyltransferase from Pyrococcus horikoshii: Molecular cloning and characterization. Biochem. Biophys. Res. Commun. 2005, 329, 429–436.
- Qu, Q.; Lee, S.J.; Boos, W. TreT, a novel trehalose glycosyltransferring synthase of the hyperthermophilic archaeon Thermococcus litoralis. J. Biol. Chem. 2004, 279, 47890–47897.
- Kouril, T.; Zaparty, M.; Marrero, J.; Brinkmann, H.; Siebers, B. A novel trehalose synthesizing pathway in the hyperthermophilic Crenarchaeon Thermoproteus tenax: The unidirectional TreT pathway. Arch. Microbiol. 2008, 190, 355.
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-phosphate: Connecting plant metabolism and development. Front. Plant Sci. 2011, 2, 70.
- Gonçalves, S.; Borges, N.; Santos, H.; Matias, P.M. Crystallization and preliminary X-ray analysis of mannosyl-3-phosphoglycerate synthase from Thermus thermophilus HB27. Acta Cryst. Sect. F 2009, 65, 1014–1017.
- Neves, C.; da Costa, M.S.; Santos, H. Compatible Solutes of the Hyperthermophile Palaeococcus ferrophilus: Osmoadaptation and Thermoadaptation in the Order Thermococcales. Appl. Environ. Microbiol. 2005, 71, 8091–8098.

