 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Olivier Hue | + 2223 word(s) | 2223 | 2021-10-28 11:28:08 | | | |
| 2 | Beatrix Zheng | Meta information modification | 2223 | 2021-10-29 05:39:45 | | |
Video Upload Options
To determine the relationships between limiting factors and neuromuscular activity during a self-paced 20-km cycling time trial and evaluate the effect of environmental conditions on fatigue indices. Methods: Ten endurance-trained and heat-acclimated athletes performed in three conditions (ambient temperature, relative humidity): HUMID (30 °C, 90%), DRY (35 °C, 46%) and NEUTRAL (22 °C, 55%). Voluntary muscular contractions and electromagnetic stimulations were recorded before and after the time trials to assess fatigue. The data on performance, temperature, heat storage, electromyogram, heart rate and rating of perceived exertion data were analyzed. Results: Performance was impaired in DRY and HUMID compared with NEUTRAL environment (p < 0.05). The force developed by the vastus lateral muscle during stimulation of the femoral nerve remained unchanged across conditions. The percentage of integrated electromyogram activity, normalized by the value attained during the pre-trial maximal voluntary contraction, decreased significantly throughout the trial only in HUMID condition (p < 0.01). Neuromuscular activity in peripheral skeletal muscle started to fall from the 11th km in HUMID and the 15th km in DRY condition, although core temperature did not reach critical values. Conclusions: These alterations suggest that afferences from core/skin temperature regulate the central neural motor drive, reducing the active muscle recruited during prolonged exercise in the heat in order to prevent the system from hyperthermia.
1. Introduction
2. Analysis on Results
Performance
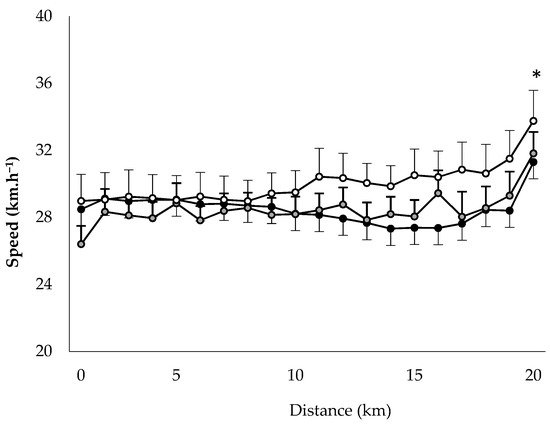
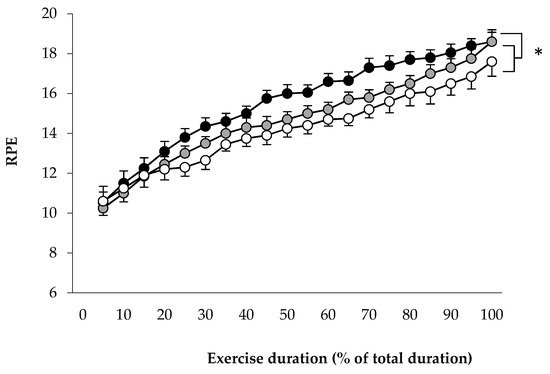
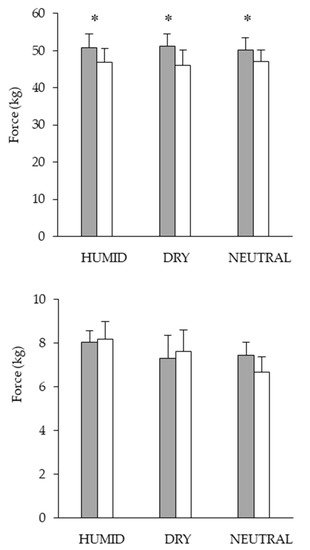

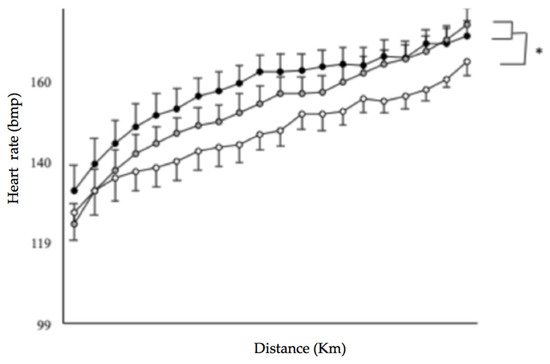
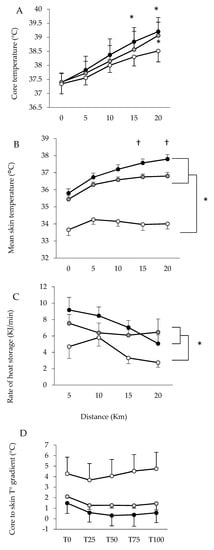
3. Current Insights
References
- Huan, L.; Schlader, Z.; Che Muhamed, A.; Zheng, H.; Stannard, S.; Kondo, N.; Cotter, J.; Mundel, T. Differences in dry-bulb temperature do not influence moderate-duration exercise performance in warm environments when vapor pressure is equivalent. Eur. J. Appl. Physiol. 2020, 120, 841–852.
- Tucker, R.; Rauch, L.; Harley, Y.X.; Noakes, T.D. Impaired exercise performance in the heat is associated with an anticipatory reduction in skeletal muscle recruitment. Pflugers Arch. 2004, 448, 422–430.
- Baillot, M.; Hue, O. Hydration and thermoregulation during a half-ironman performed in tropical climate. J. Sports Sci. Med. 2015, 14, 263–268.
- Baillot, M.; Le Bris, S.; Hue, O. Fluid replacement strategy during a 27-Km trail run in hot and humid conditions. Int. J. Sports Med. 2014, 35, 147–152.
- Nybo, L.; Nielsen, B. Hyperthermia and Central Fatigue during Prolonged Exercise in Humans. J. Appl. Physiol. 2001, 91, 1055–1060.
- Cuddy, J.S.; Hailes, W.S.; Ruby, B.C. A reduced core to skin temperature gradient, not a critical core temperature, affects aerobic capacity in the heat. J. Therm. Biol. 2014, 43, 7–12.
- González-Alonso, J.; Teller, C.; Andersen, S.L.; Jensen, F.B.; Hyldig, T.; Nielsen, B. Influence of Body Temperature on the Development of Fatigue during Prolonged Exercise in the Heat. J. Appl. Physiol. 1999, 86, 1032–1039.
- Walters, T.J.; Ryan, K.L.; Tate, L.M.; Mason, P.A. Exercise in the Heat Is Limited by a Critical Internal Temperature. J. Appl. Physiol. (1985) 2000, 89, 799–806.
- González-Alonso, J.; Calbet, J.A.; Nielsen, B. Muscle Blood Flow Is Reduced with Dehydration during Prolonged Exercise in Humans. J. Physiol. 1998, 513 Pt 3, 895–905.
- Maughan, R.J.; Otani, H.; Watson, P. Influence of Relative Humidity on Prolonged Exercise Capacity in a Warm Environment. Eur. J. Appl. Physiol. 2012, 112, 2313–2321.
- Cheuvront, S.N.; Kenefick, R.W.; Montain, S.J.; Sawka, M.N. Mechanisms of Aerobic Performance Impairment with Heat Stress and Dehydration. J. Appl. Physiol. 2010, 109, 1989–1995.
- Dugas, J.P.; Oosthuizen, U.; Tucker, R.; Noakes, T.D. Rates of Fluid Ingestion Alter Pacing but Not Thermoregulatory Responses during Prolonged Exercise in Hot and Humid Conditions with Appropriate Convective Cooling. Eur. J. Appl. Physiol. 2009, 105, 69–80.
- Lee, J.K.W.; Nio, A.Q.X.; Lim, C.L.; Teo, E.Y.N.; Byrne, C. Thermoregulation, Pacing and Fluid Balance during Mass Participation Distance Running in a Warm and Humid Environment. Eur. J. Appl. Physiol. 2010, 109, 887–898.
- Beis, L.Y.; Wright-Whyte, M.; Fudge, B.; Noakes, T.; Pitsiladis, Y.P. Drinking Behaviors of Elite Male Runners during Marathon Competition. Clin. J. Sport Med. 2012, 22, 254–261.
- Shirreffs, S.M.; Maughan, R.J. Urine Osmolality and Conductivity as Indices of Hydration Status in Athletes in the Heat. Med. Sci. Sports Exerc. 1998, 30, 1598–1602.
- Che Muhamed, A.M.; Yusof, H.A.; Stannard, S.R.; Mündel, T.; Thompson, M.W. The Efficacy of Ingesting Water on Thermoregulatory Responses and Running Performance in a Warm-Humid Condition. Front. Physiol. 2019, 10, 507.
- Candas, V.; Libert, J.P.; Vogt, J.J. Effect of Hidromeiosis on Sweat Drippage during Acclimation to Humid Heat. Eur. J. Appl. Physiol. Occup. Physiol. 1980, 44, 123–133.
- Galloway, S.D.; Maughan, R.J. Effects of Ambient Temperature on the Capacity to Perform Prolonged Cycle Exercise in Man. Med. Sci. Sports Exerc. 1997, 29, 1240–1249.
- Sawka, M.N.; Leon, L.R.; Montain, S.J.; Sonna, L.A. Integrated Physiological Mechanisms of Exercise Performance, Adaptation, and Maladaptation to Heat Stress. Compr. Physiol. 2011, 1, 1883–1928.
- Roberts, M.F.; Wenger, C.B.; Stolwijk, J.A.; Nadel, E.R. Skin Blood Flow and Sweating Changes Following Exercise Training and Heat Acclimation. J. Appl. Physiol. Respir Env. Exerc Physiol. 1977, 43, 133–137.
- Schlader, Z.J.; Simmons, S.E.; Stannard, S.R.; Mündel, T. The Independent Roles of Temperature and Thermal Perception in the Control of Human Thermoregulatory Behavior. Physiol. Behav. 2011, 103, 217–224.
- Tucker, R.; Marle, T.; Lambert, E.V.; Noakes, T.D. The rate of heat storage mediates an anticipatory reduction in exercise intensity during cycling at a fixed rating of perceived exertion. J. Physiol. 2006, 574 Pt 3, 905–915.
- Kayser, B. Exercise Starts and Ends in the Brain. Eur. J. Appl. Physiol. 2003, 90, 411–419.

