Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Noman Shoaib | + 3342 word(s) | 3342 | 2021-10-12 05:46:50 | | | |
| 2 | Peter Tang | Meta information modification | 3342 | 2021-10-14 04:48:44 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Shoaib, N. Plastidial Starch Phosphorylase in Starch Metabolism. Encyclopedia. Available online: https://encyclopedia.pub/entry/15019 (accessed on 26 July 2026).
Shoaib N. Plastidial Starch Phosphorylase in Starch Metabolism. Encyclopedia. Available at: https://encyclopedia.pub/entry/15019. Accessed July 26, 2026.
Shoaib, Noman. "Plastidial Starch Phosphorylase in Starch Metabolism" Encyclopedia, https://encyclopedia.pub/entry/15019 (accessed July 26, 2026).
Shoaib, N. (2021, October 14). Plastidial Starch Phosphorylase in Starch Metabolism. In Encyclopedia. https://encyclopedia.pub/entry/15019
Shoaib, Noman. "Plastidial Starch Phosphorylase in Starch Metabolism." Encyclopedia. Web. 14 October, 2021.
Copy Citation
Starch phosphorylase is a member of the GT35-glycogen-phosphorylase superfamily. Glycogen phosphorylases have been researched in animals thoroughly when compared to plants. Genetic evidence signifies the integral role of plastidial starch phosphorylase (PHO1) in starch biosynthesis in model plants. The counterpart of PHO1 is PHO2, which specifically resides in cytosol and is reported to lack L80 peptide in the middle region of proteins as seen in animal and maltodextrin forms of phosphorylases. The function of this extra peptide varies among species and ranges from the substrate of proteasomes to modulate the degradation of PHO1 in Solanum tuberosum to a non-significant effect on biochemical activity in Oryza sativa and Hordeum vulgare.
starch phosphorylase
plastidial starch phosphorylase
amyloplastic phosphorylase
synthetic activity
phosphorolytic activity
1. Introduction
Phosphorylase (EC 2.4.1.1), designated as starch phosphorylase (PHO) in plants and glycogen phosphorylase (gPHO) in animals, has been studied for decades due to its multiple functions. Phosphorylase was first discovered in the liver tissue of mammals in 1939 and was reported to work in the reversible transfer of the glucosyl unit from glucose-1-phosphate to glycogen with the release of inorganic phosphate [1]. In the 1940s, the same type of reaction was described in higher plants [2][3]. Phosphorylase shares a solid evolutionary relationship among animals, plants, algae, and bacteria. In comparison to animals, higher plants and green algae have two distinct forms of starch phosphorylases, a plastidial or amyloplastic form (PHO1 or SP-L; low affinity for glycogen) and a cytosolic form (PHO2 or SP-H; high affinity for glycogen) [4]. These two forms have significant sequence similarities to each other but largely differ in their molecular sizes, substrate specificities, and physiological roles [5]. The alignment of proposed phosphorylase sequences from a wide range of species have shown that phosphorylase in higher plants is structurally similar to the animals and bacterial forms (maltodextrin phosphorylase) but differ in transit peptides of up to 50 amino acids in the N-terminus [6][7][8] and L80 domain (an extra peptide of up to 82 amino acid residues). The L80 domain is present in the middle of PHO1 and absent in animal, bacteria, and PHO2 forms of higher plant phosphorylases [9]. The presence of this prominent extra peptide in the middle region of PHO1 can be responsible for the diverse biochemical activities of PHO1 [10][11]. Previous findings have claimed that L80 peptide contains a variable set of negatively charged amino acids, phosphorylation sites, and a potential PEST motif (rich in proline, glutamic acid, serine, and threonine) [12]. It has been reported that the L80 insertion sterically hinders the binding of large polysaccharides with PHO1 in Solanum tuberosum [13]. In addition, L80 insertion was found to be an active substrate for proteasomes (20S or 26S), when one of the N-terminal amino acid residues had been modified by phosphorylation in Ipomoea batatas [14]. The PHO1 in Solanum tuberosum maintains its intact structure in young tubers but steadily degrades into smaller peptides as the tubers become mature [7]. Both the intact (105 kDa) and degraded (55 kDa) forms of PHO1 have been reported in Vigna radiata. Interestingly, both forms were identified to make active catalytic complexes [15]. In contrast, the PHO1 of cereals including Oryza sativa [16][17], Zea mays [8], Hordeum vulgare [18], and Triticum aestivum [9] is not degraded by the proteasome, even though they all have a PEST motif in L80 insertion. A detailed study would be required to uncover these differences to reach a conclusion.
2. Starch Metabolism and Phosphorylase
2.1. Starch Biosynthesis
Starch is an important component of plants and has been subjected to profound research; however, understanding its granule synthesis remains obscure. Amylose and amylopectin are the major components of starch [19]. Some minor components such as minerals, lipids, and proteins are also important parts of starch granules (comprising a small percentage of the total weight of the granule) [20]. Starch biosynthesis is a complex mechanism that involves the activity of more than 15 enzymes [21]. These enzymes, each with various tissue-specific and developmental specific isoforms of starch synthases (SSs, EC 2.4.1.21), starch branching enzymes (SBEs, EC 2.4.1.18), and starch debranching enzymes (DBEs, 3.2.1.41; mainly isoamylases (ISAs)) are the major groups of enzymes involved in starch metabolism. ADP-Glc pyrophosphorylase (AGPase, EC 2.7.7.27) is the key enzyme in the first mandatory step of starch biosynthesis that produces the adenosine diphosphate glucose ADP-Glc from glucose-1-phosphate (Glc-1-P) and ATP [22]. ADP-Glc acts as a substrate to extend the α-1,4 linked glucans through SSs. There are five main isoforms of SSs essentially present in all crops, namely SSI, SSII, SSIII, SSIV, and granule bound starch synthase (GBSS). Among these SSI, SSII, and SSIII are involved in the synthesis of amylopectin [23][24], whereas GBSS is involved in amylose synthesis as the mutants that show loss of function of GBSS produce amylose-free starch [25][26]. SBEs introduce the branch point and cleave the elongated α-1,4 linked glucans and transfer that segment to the sides of an acceptor chain through α-1,6 linkage. DBEs further cleave the α-1,6 linked chains that are needed for proper amylopectin structure. In addition, they are thought to promote the crystallization of amylopectin through trimming and degrading the branched glucan [27][28]. Two of three isoforms of ISA, ISAI and ISAII, take part in amylopectin synthesis, whereas ISAIII is involved in the degradation of starch [29]. Furthermore, a study on the expression level of PHO1 in Hordeum vulgare [30][31], Arabidopsis thaliana [32], Triticum aestivum [9], and Oryza sativa [16] has indicated the involvement of PHO1 in starch metabolism. The comprehensive details of the structure and synthesis of starch can be found in recent reviews [28][33].
2.2. Initiation of Starch Granule in Comparison to Glycogen
Based on biochemical evidence, it was assumed that phosphorylase was involved in the initiation and amplification of glycogen and starch biosynthesis in animals and plants, respectively [9][16][34][35]. However, the discovery of glucosyl-transferase in the liver tissue ruled out this assumption for the glycogen synthesis of gPHO. Indeed, glycogenin is a precise initiator protein in the de novo biosynthesis of glycogen in fungi and animals in a process of self-glucosylation with UDP-glucose to generate covalently bound malto-oligosaccharides (MOs) [36]. Bacteria likely utilize the glycogen synthase (GS) to transfer the sugar moiety from ADP-Glc to distinctive binding amino acid residues in the process of de novo MO synthesis [37]. There are no experimental reports available to date that indicate the participation of phosphorylase in the glycogen initiation process. On the contrary, no self glucosylation proteins have been reported in plants [38]. Therefore, unlike animals the starch initiation mechanism in the plants could be different from the glycogen initiation mechanism in bacteria, fungi, and animals. In addition, there are indications that AGPase is not entirely involved in starch biosynthesis as it requires a primer of reasonable chain length (G2–G4) [39]. However, a study on potato tuber phosphorylase claimed that de novo synthesis of amylose occurs in the absence of primer [40]. AGPase and SSs were not present in the early stages of development in maize kernel, but some quantity of starch was synthesized already [41]. Moreover, they further suggested that the synthesis of starch must follow a different route at the start [41].
There can be two possible routes for starch synthesis (Figure 1), either through the PHO1 or AGPase/SSs, which depend on the availability of substrate-level (Glc-1-P) [42][43][44]. The PHO1 route is more energy-efficient than AGPase/SSs as it required a higher concentration of Glc-1-P [12]. PHO1 could have a special importance during the early stages of endosperm development because the activity of phosphorylase is 40-fold greater than SSs in initial stages and 10-fold greater in lateral stages of development in developing maize kernels [45]. A mutant (sta4) lacking the PHO1 displayed reduced levels of starch with abnormally shaped granules with high amylose content, arguing for a role of PHO1 in starch biosynthesis in Chlamydomonas [46]. Shrunken 4 (sh4), a mutant of Zea mays with no activity of phosphorylase resulted in the reduced starch accumulation and shrunken morphology of maize kernels [45], suggesting the PHO1 can determine the granule shape/morphology.
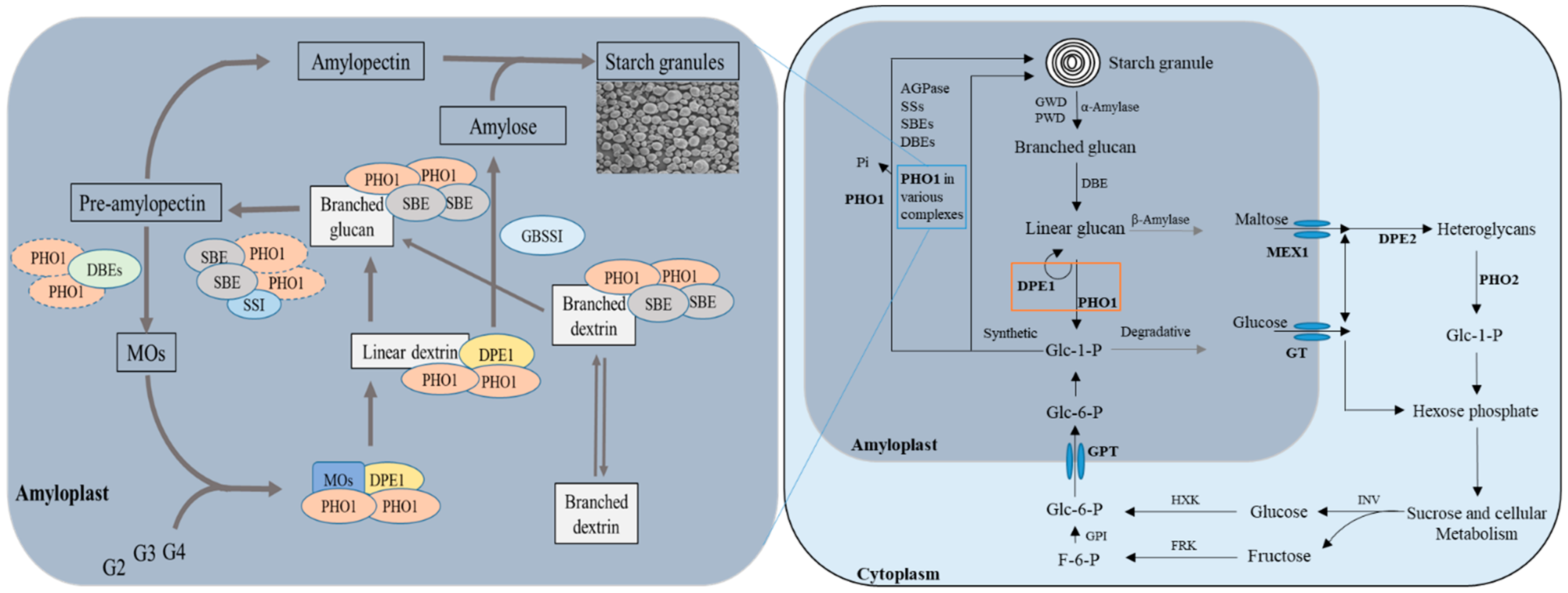
Figure 1. Putative roles of PHO1 and PHO2 in starch biosynthesis and degradation in the non-photosynthetic cell. Abbreviations: GWD: glucan water dikinase, PWD: phospho-glucan water dikinase, DBE: debranching enzyme, DPE1, plastidial disproportionating enzyme, PHO1, plastidial starch phosphorylase, AGPase: ADP-Glc pyrophosphorylase, SSs: starch synthases, SBEs: starch branching enzymes, DPE2: cytosolic disproportionating enzymes, PHO2: cytosolic starch phosphorylase, INV: invertase, HXK: hexose kinase, FRK: fructose kinase, GPI: glucose-6-phosphate isomerase, GT: glucose transporter, MEX1: maltose transporter, GPT: glucose-6-phosphate transporter, Pi: inorganic phosphate, Glc-1-P: glucose-1-phosphate, Glc-6-P: glucose-6-phoshphate, F-6-P: fructose-6-phosphate and MOs: malto oligosaccharide, G2: maltose, G3: maltotriose, and G4: maltotetraose. The orange box indicates the concerted action of DPE1 and PHO1, which might define the fate of the metabolites. The trimmed MOs that are generated by the action of DBEs and PHO1 and branched oligosaccharides enter the starch biosynthetic pathway together, where they are first elongated by the concerted action of DPE1 and PHO1 metabolon, followed by the further elongation of by the SBEs and PHO1 complex. Alternatively, branched oligosaccharides are further elongated and branched by the combined action of PHO1 and isozymes of SBEs. Fusion of the generated products leads to the formation of pre-amylopectin via the possible complex of PHO1, SSs, and SBEs isozymes (still unknown). Trimming of MOs on the pre-mature amylopectin leads to its maturation that further combines with amylose (generated via the action of GBSSI) to form the starch granules. Produced from [12][17][28][33][39][47][48][49].
2.3. Multiple Complex Formation
Increasing evidence supports that starch biosynthesis does not consist of some simple and isolated reactions. The protein–protein interaction and regulatory coordination among starch biosynthetic enzymes have been reported as the key features in starch biosynthesis [5][14][50]. A genetic study has reflected that disproportionating enzyme 1 (DPE1, EC 2.4.1.25) and PHO1 are potentially involved in the initiation of storage starch biosynthesis in plants [17]. Based on previously reported work [10][47], it has been revealed that PHO1 forms a complex with DPE1 and serves as a metabolon in the recycling of MOs back to growing amylopectin through Glc-1-P synthesis in Oryza sativa. The phosphorolytic reaction of PHO1 was stimulated by DPE1 in Chlamydomonas [51]. It has also been shown that PHO1 forms complexes with various SBEs and SSs in a phosphorylation manner to regulate the starch biosynthesis and is involved in elongation and branching of MOs alternatively, followed by further elongation via functional interaction between PHO1 and isozymes of branching enzymes (Figure 1). Another proposed mechanism explains the direct action of PHO1 on the surface of starch to add glucose units from Glc-1-P with the release of inorganic phosphate (Pi) [9]. These observations suggest a certain role of PHO1 in starch biosynthesis and metabolism of MOs.
2.4. Starch Degradation
Several starch-degrading enzymes have been identified [52][53] and can be generally classified into two categories: phosphorolytic and hydrolytic degrading enzymes. The branched and linear glucans are produced from the starch degradation through the actions of newly identified candidates: glucan water dikinase (GWD; EC 2.7.9.4) and phosphoglucan water dikinase (PWD; EC 2.7.9.5) [54]. However, α-amylase (EC 3.2.1.1) is thought to be the first enzyme for starch degradation. Branched glucans are further reduced to linear glucans by DBEs. PHO1 and β-amylase produce the neutral sugars from linear glucans and eventually generate metabolites that are transported to the cytosol by their specific transporters [32]. Concerted action of cytosolic forms of PHO and DPE produce heteroglycans (heteropolysaccharides containing two or more different monosaccharide units) and hexose phosphates, which subsequently enter sucrose and cellular metabolism pathways [48]. To keep the balance between cytosol and amyloplast, cellular metabolites can enter the starch biosynthesis pathway through the actions of invertase (INV), hexose kinase (HXK), and fructose kinase (FRK) enzymes (Figure 1).
2.5. Additional Role of PHO1
Besides storage starch synthesis in non-photosynthetic cells, it is stated that PHO1 may have an additional role in modulating the photosynthetic activity by interacting with the PsaC region of Photosystem-I (PSI) in photosynthetic cells of Oryza sativa [12][33]. It could influence the PSI activity indirectly, via the pentose phosphate pathway (PPP), as the PHO1− (lacking in PHO1) mutant (bmf136) of rice was reported to have different PSI activity and exhibited lower photosynthetic performance in comparison to wild type [16][42]. The phenotype of bmf136 was rescued to normal when it was complemented with the PHO1. However, when these plants were complemented with the PHO1∆L80 (PHO1 with excised L80 region) instead of typical, enhanced photosynthetic activity was observed. These findings suggest that PHO1 also acts as a modulator of photosynthesis, and the L80 domain is a controller in the photosynthesis modulation.
3. Synthetic and Phosphorolytic Activity of Phosphorylase
Both PHO1 and PHO2 can catalyze the phosphorolysis of glucans to generate Glc-1-P and Pi in a reversible reaction. Since the reaction is reversible, the appropriate catalyzing direction under physiological conditions is still in debate [12]. In the synthetic direction, phosphorylase is involved in the transfer of the glucosyl unit from Glc-1-P to the growing glucan chain with the release of inorganic phosphate [46][55][56]. In the phosphorolytic direction, the same reaction takes place in a reverse direction. Which reaction is favored in starch biosynthesis is dependent on the Pi/Glc-1-P ratios; a higher pi/Glc-1-P ratio favors the reaction in the phosphorolytic direction and a low Pi/Glc-1-P ratio favors the reaction in the synthetic direction [57]. However, evidence suggests that the direction of PHO1 activity can be also controlled by some other factors than available substrate levels [33]. In comparison to PHO1, PHO2 of higher plants has been reported to degrade the maltose phosphorolytically, a result of starch degradation [34][58]. However, PHO1 is supposed to play an efficient synthetic role in starch biosynthesis because of its expression and localization, in parallel to storage starch synthesis and accumulation therein [9]. A study of PHO1 in Zea mays has shown that it stimulates the reaction in the phosphorolytic direction with MOs [5][59], and in Oryza sativa PHO1 showed a preferable role in the synthetic direction over degradation even with the high level of Pi [38][44]. Phosphorylase from Tetraselmis subcordiformis (marine green microalga) preferred a two-fold higher affinity in the synthetic direction over phosphorolytic direction [60]. Further to supporting the synthetic role, PHO1 has been seen to be involved in actively taking up the Glc-1-P in tuber parenchyma cells and depositing it to storage starch granules in Solanum tuberosum [61]. PHO1 actively interacts with other starch synthetic enzymes to form the functionally active protein–protein complex and to modulate its synthetic activity [17][47][62][63]. PHO2 precisely exists in the cytoplasm where it can interact with DPE2 and modulates the degradation reactions only [17][33].
Further, to elaborate the synthetic and phosphorolytic activity of phosphorylase [64], Subsinghe, R.M., performed experiments on Zea mays PHO1 at various developmental stages and reported that PHO1 remains active throughout the endosperm development and showed synthetic activity. At 22 days after anthesis (DAA), PHO1 showed both synthetic and phosphorolytic activity when tested on native affinity zymograms [18]. In Hordeum vulgare, PHO1 also confirmed the synthetic activity in developing endosperm, which was highest at 12 DAA and gradually decreased afterward, but the activity does not correlate with the expression level, possibly because of substrate preferences. The preference of substrate by PHO1 has been intensively investigated in various plant species [8][46][65][66][67][68]. PHO1 in Ipomoea batatas showed low binding affinity toward starch (high molecular weight) and high binding affinity towards MOs (low molecular weight) [65]. In contrast, PHO1 in Zea mays showed high binding affinity toward high molecular weight compounds and low binding affinity toward low molecular weight compounds [66]. In Zea mays [8], Solanum tuberosum [67], and Spinacia oleracea [68] synthetic activity of PHO1 was reported to be high when amylopectin was used as a substrate in comparison to highly branched glycogen. In Solanum tuberosum L80 insertion was reported to block the binding affinity of PHO1 with high molecular weight compounds [65]. In cereals, whether L80 insertion affects the substrate affinity has not been identified or described yet.
4. Genomics of PHO1
During the past few years, great efforts have been made to sequence the whole genomes of plants. The availability of data is important in answering the biological questions in comparative studies [69][70]. In parallel, scientists have discovered the gene encoding phosphorylase (PHO) and interpreted the gene structure, identity, localization, conserved regions, and the expression in wide range of species including maize (Zea mays; NP_001296783.1) [8][55][71], brome (Brachypodium distachyon; XP_003559211.1) [72], barley (Hordeum vulgare; KAE8783983.1) [18], rice (Oryza sativa; XP_015631420.1) [16][44][49], wheat (Triticum aestivum; ACC59201.1) [9], potato (Solanum tuberosum; NP_001275215.1) [7][73][74], sweet potato (Ipomoea batatas; M64362.1) [14][75], orange (Citrus clementina; NW_0062622139.1), mung bean (Vigna radiata; LOC106770459) [15][76], cotton (Gossypium Arboreum; LOC108456840) [77], bunch grass (Panicum hallii; LOC112876003), tomato (Solanum lycopersicum; XP_0042234848.1) [78], and wild tomato (Solanum habrochaites; MG962532.1) [78]. gPHO in animals has been investigated comprehensively as compared to PHO1 in plants. Only PHO1 of Zea mays, Oryza sativa, and Hordeum vulgare has been investigated comprehensively in main cereal crops. PHO1 for Triticum aestivum, Setaria italic, and Ipomoea batatas has not been demonstrated and characterized yet and should be the target of future research.
5. Structure and Comparison of PHO1
The comparison of the PHO1 of Zea mays [79] with other plant, microorganism, and animal forms of phosphorylases indicated that they vary in size. Alignment of main phosphorylase protein sequences has revealed the conserved junctions for various functional domains or sites of the protein including the catalytic site, residue binding site, coil entry, glycine-rich peptides, glutamic acid-rich peptides, repeats, L80 peptides, pyridoxal phosphate (PLP) binding residues, acetylation, and phosphorylation sites (Figure 2). Four regions, the N-terminal, regulatory domain, L80 insertion, and catalytic domain, can be determined as indicated in the phosphorylase protein structure comparison analysis (Figure 2). It was found that PHO from each species consists of a highly conserved GT-35-glycogen-phosphorylase domain [80]. In addition, regulatory and catalytic domains have been found highly conserved among all proteins. The N-terminus and L80 insertion have been found to vary among species. Only the PHO1 of the plants has L80 insertion as compared to PHO2 of plants, maltodextrin phosphorylase, and gPHO (Figure 3). The PEST motif is found on the L80 insertion of PHO1 in various plants [12]. The PLP binding junction has been found conserved among all PHO proteins, but it tends to vary in residue numbers. The regulatory domain mainly consists of glycogen storage and glucose-6-phosphate (Glc-6-P) binding sites. However, the catalytic domain mainly consists of glucose binding (active site), PLP binding, and nucleoside inhibitor residues. Alignment of sequences and information for functional sites indicated that limited research on plant phosphorylases has been conducted as compared to the animal form of phosphorylase. In addition, for 3D structure insights of phosphorylase protein, mammalian phosphorylase has received considerable attention, and Hordeum vulgare is the only plant in which the 3D structure of PHO1 has been established completely [18].
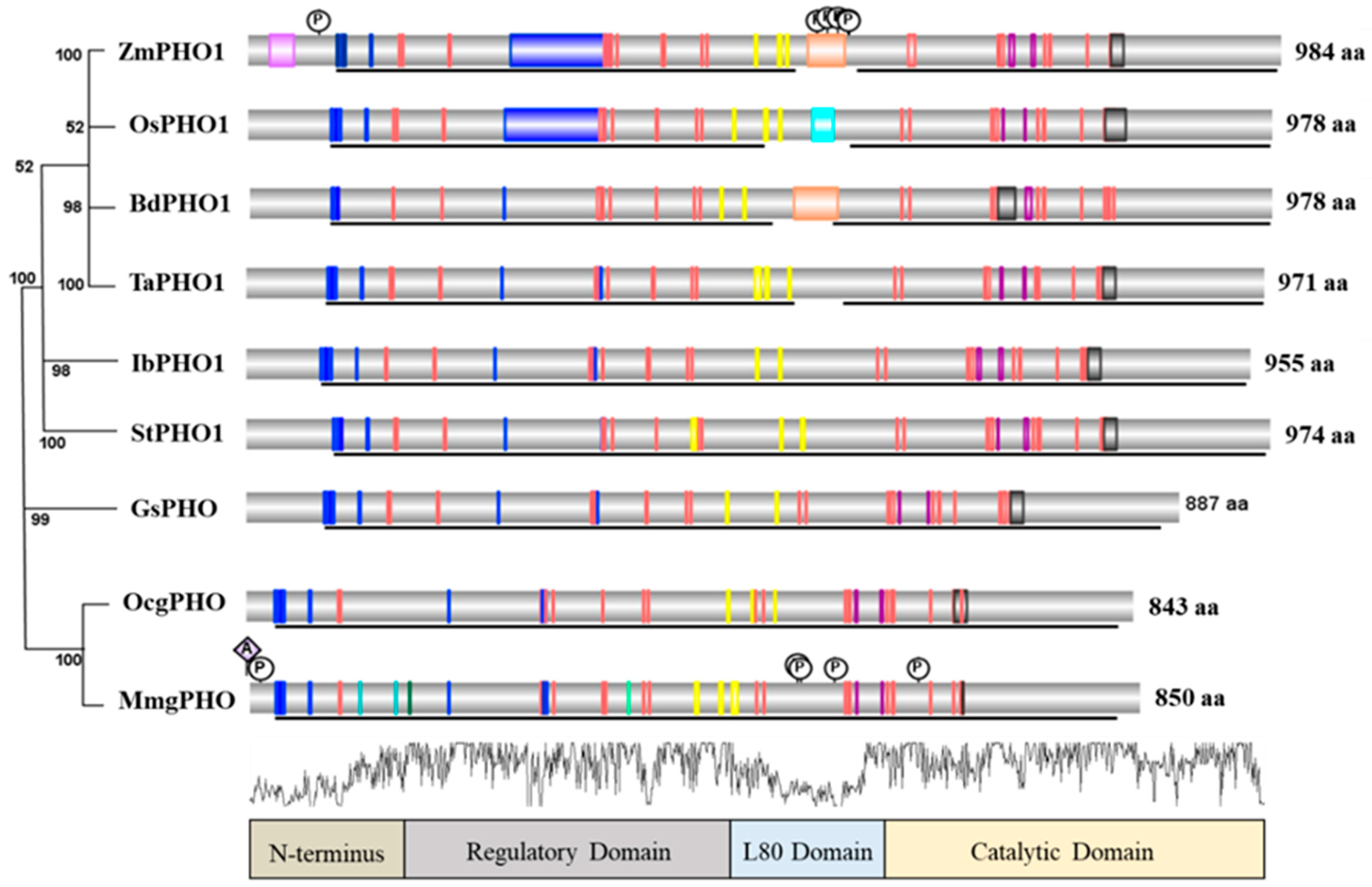
Figure 2. Annotated protein structure of phosphorylases from a wide range of species. Arranged in phylogenetic order. Representation is divided into four basic domains. Abbreviations: Zm: Zea mays, Os: Oryza sativa, Bd: Brachypodium distachyon, Ta: Triticum aestivum, Ib: Ipomoea batatas, St: Solanum tuberosum, Gs: Galdieria sulphuraria, Oc: Oryctolagus cuniculus, and Mm: Mus musculus. Amino acid numbers are varying among species. Vertical colored bars represent the different functional sites. Blue bars represent the homodimer interface for peptide binding, orange bars represent the catalytic regions, sky blue for coil entry, green for allosteric control region, gray for modification sites with pyridoxal phosphate (PLP), pink for glycine-rich domain/glutamic-acid-rich sites, yellow for glycogen storage sites, and purple for purine nucleotide inhibitor sites for caffeine. P represents the phosphorylation sites and A the acetylation sites. Brown brackets represent the PEST motif. Black lines indicate the highly conserved region that belongs to the GT35-glycogen-phosphorylase superfamily. Overall N-terminus and L80 domains are highly variable. Regulatory and catalytic domains display a high similarity index (more than 88%).

Figure 3. Alignment of middle regions (L80 peptide) of PHO1 from a wide range of species. Abbreviations are the same as in Figure 2. The percentage represents the identical index of the proteins that was calculated as a reference to Zea mays PHO1. Numbers express a portion of the amino acid sequence in the primary structure of proteins. Red lines indicate the potential PEST motif predicted by the software (https://emboss.bioinformatics.nl/cgi-bin/emboss/epestfind) accessed on 15 June 2021.
In relation to evolution, a highly divergent phylogenetic pattern has been observed among the phosphorylases (Figure 2). PHO1 of Zea mays showed a closer relationship with the PHO1 in Panicum hallii and Setaria italica. Hordeum vulgare showed a closer relationship with the PHO1 of Triticum aestivum than those in Zea mays and Oryza Sativa. All of the mentioned phosphorylases are likely to be evolved from a single species.
References
- Cori, G.T.; Cori, C.F.; Schmidt, G. The role of glucose-l-phosphate in the formation of blood sugar and synthesis of glycogen in the liver. J. Biol. Chem. 1939, 142, 355–366.
- Green, D.E.S.; Stumpf, P.K. Starch Phosphorylase of potato. J. Biol. Chem. 1942, 142, 355–366.
- Hanes, C.S. The breakdown and synthesis of starch by an enzyme system from pea seeds. Proc. R. Soc. Lond. Ser. B Boil. Sci. 1940, 128, 421–450.
- Tsai, C.Y.; Nelson, O.E. Phosphorylases I and II of Maize Endosperm. Plant Physiol. 1968, 43, 103–112.
- Subasinghe, R.M.; Liu, F.; Polack, U.C.; Lee, E.A.; Emes, M.J.; Tetlow, I.J. Multimeric states of starch phosphorylase determine protein–protein interactions with starch biosynthetic enzymes in amyloplasts. Plant Physiol. Biochem. 2014, 83, 168–179.
- Nakano, K.; Fukui, T. The complete amino acid sequence of potato alpha-glucan phosphorylase. J. Biol. Chem. 1986, 261, 8230–8236.
- Brisson, N.; Giroux, H.; Zollinger, M.; Camirand, A.; Simard, C. Maturation and Subcellular Compartmentation of Potato Starch Phosphorylase. Plant Cell 1989, 1, 559.
- Yu, Y.; Mu, H.H.; Wasserman, B.P.; Carman, G.M. Identification of the Maize Amyloplast Stromal 112-kD Protein as a Plastidic Starch Phosphorylase. Plant Physiol. 2001, 125, 351–359.
- Tickle, P.; Burrell, M.M.; Coates, S.A.; Emes, M.J.; Tetlow, I.J.; Bowsher, C.G. Characterization of plastidial starch phosphorylase in Triticum aestivum L. endosperm. J. Plant Physiol. 2009, 166, 1465–1478.
- Hwang, S.-K.; Singh, S.; Çakir, B.; Satoh, H.; Okita, T.W. The plastidial starch phosphorylase from rice endosperm: Catalytic properties at low temperature. Planta 2016, 243, 999–1009.
- Cuesta-Seijo, J.A.; Nielsen, M.M.; Ruzanski, C.; Krucewicz, K.; Beeren, S.R.; Rydhal, M.G.; Yoshimura, Y.; Striebeck, A.; Motawia, M.S.; Willats, W.G.T.; et al. In Vitro Biochemical Characterization of All Barley Endosperm Starch Synthases. Front. Plant Sci. 2016, 6, 1265.
- Hwang, P.; Koper, K.; Okita, T.W. The plastid phosphorylase as a multiple-role player in plant metabolism. Plant Sci. 2020, 290, 110303.
- Albrecht, T.; Greve, B.; Pusch, K.; Kossmann, J.; Buchner, P.; Wobus, U.; Steup, M. Homodimers and heterodimers of Pho1-type phosphorylase isoforms in Solanum tuberosum L. as revealed by sequence-specific antibodies. JBIC J. Biol. Inorg. Chem. 1998, 251, 343–352.
- Lin, Y.-C.; Chen, H.-M.; Chou, I.-M.; Chen, A.-N.; Chen, C.-P.; Young, G.-H.; Lin, C.-T.; Cheng, C.-H.; Chang, S.-C.; Juang, R.-H. Plastidial Starch Phosphorylase in Sweet Potato Roots Is Proteolytically Modified by Protein-Protein Interaction with the 20S Proteasome. PLoS ONE 2012, 7, e35336.
- Ko, Y.-T.; Chang, J.-Y.; Lee, Y.-T.; Wu, Y.-H. The Identification of Starch Phosphorylase in the Developing Mungbean (Vigna radiata L.). J. Agric. Food Chem. 2005, 53, 5708–5715.
- Satoh, H.; Shibahara, K.; Tokunaga, T.; Nishi, A.; Tasaki, M.; Hwang, S.-K.; Okita, T.; Kaneko, N.; Fujita, N.; Yoshida, M.; et al. Mutation of the Plastidial α-Glucan Phosphorylase Gene in Rice Affects the Synthesis and Structure of Starch in the Endosperm. Plant Cell 2008, 20, 1833–1849.
- Hwang, S.-K.; Koper, K.; Satoh, H.; Okita, T.W. Rice Endosperm Starch Phosphorylase (Pho1) Assembles with Disproportionating Enzyme (Dpe1) to Form a Protein Complex That Enhances Synthesis of Malto-oligosaccharides. J. Biol. Chem. 2016, 291, 19994–20007.
- Cuesta-Seijo, J.A.; Ruzanski, C.; Krucewicz, K.; Meier, S.; Hägglund, P.; Svensson, B.; Palcic, M.M. Functional and structural characterization of plastidic starch phosphorylase during barley endosperm development. PLoS ONE 2017, 12, e0175488.
- Maddelein, M.L.; Libessart, N.; Bellanger, F.; Delrue, B.; D’Hulst, C.; Koornhuyse, N.V.D.; Fontaine, T.; Wieruszeski, J.M.; Decq, A.; Ball, S. Toward an understanding of the biogenesis of the starch granule. Determination of granule-bound and soluble starch synthase functions in amylopectin synthesis. J. Biol. Chem. 1994, 269, 25150–25157.
- Putseys, J.A.; Derde, L.J.; Lamberts, L.; Östman, E.; Björck, I.M.; Delcour, J.A. Functionality of Short Chain Amylose-Lipid Complexes in Starch-Water Systems and Their Impact on In Vitro Starch Degradation. J. Agric. Food Chem. 2010, 58, 1939–1945.
- Goren, A.; Ashlock, D.; Tetlow, I.J. Starch formation inside plastids of higher plants. Protoplasma 2018, 255, 1855–1876.
- Yu, G.; Lv, Y.; Shen, L.; Wang, Y.; Qing, Y.; Wu, N.; Li, Y.; Huang, H.; Zhang, N.; Liu, Y.; et al. The Proteomic Analysis of Maize Endosperm Protein Enriched by Phos-tag(tm) Reveals the Phosphorylation of Brittle-2 Subunit of ADP-Glc Pyrophosphorylase in Starch Biosynthesis Process. Int. J. Mol. Sci. 2019, 20, 986.
- Fujita, N.; Kubo, A.; Suh, D.-S.; Wong, K.-S.; Jane, J.-L.; Ozawa, K.; Takaiwa, F.; Inaba, Y.; Nakamura, Y. Antisense inhibition of isoamylase alters the structure of amylopectin and the physicochemical properties of starch in rice endosperm. Plant Cell Physiol. 2003, 44, 607–618.
- Fujita, N.; Yoshida, M.; Asakura, N.; Ohdan, T.; Miyao, A.; Hirochika, H.; Nakamura, Y. Function and Characterization of Starch Synthase I Using Mutants in Rice. Plant Physiol. 2006, 140, 1070–1084.
- Hanashiro, I.; Itoh, K.; Kuratomi, Y.; Yamazaki, M.; Igarashi, T.; Matsugasako, J.-I.; Takeda, Y. Granule-Bound Starch Synthase I is Responsible for Biosynthesis of Extra-Long Unit Chains of Amylopectin in Rice. Plant Cell Physiol. 2008, 49, 925–933.
- Ortiz-Marchena, M.I.; Albi, T.; Lucas-Reina, E.; Said, F.E.; Romero-Campero, F.J.; Cano, B.; Ruiz, M.T.; Romero, J.M.; Valverde, F. Photoperiodic Control of Carbon Distribution during the Floral Transition in Arabidopsis. Plant Cell 2014, 26, 565–584.
- Streb, S.; Delatte, T.; Umhang, M.; Eicke, S.; Schorderet, M.; Reinhardt, D.; Zeeman, S.C. Starch Granule Biosynthesis in Arabidopsis Is Abolished by Removal of All Debranching Enzymes but Restored by the Subsequent Removal of an Endoamylase. Plant Cell 2009, 20, 3448–3466.
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807.
- Delatte, T.; Umhang, M.; Trevisan, M.; Eicke, S.; Thorneycroft, D.; Smith, S.; Zeeman, S. Evidence for Distinct Mechanisms of Starch Granule Breakdown in Plants. J. Biol. Chem. 2006, 281, 12050–12059.
- Ma, J.; Jiang, Q.-T.; Zhang, X.-W.; Lan, X.-J.; Pu, Z.-E.; Wei, Y.-M.; Liu, C.; Lu, Z.-X.; Zheng, Y.-L. Structure and expression of barley starch phosphorylase genes. Planta 2013, 238, 1081–1093.
- Higgins, J.E.; Kosar-Hashemi, B.; Li, Z.; Howitt, C.A.; Larroque, O.; Flanagan, B.; Morell, M.K.; Rahman, S. Characterization of starch phosphorylases in barley grains. J. Sci. Food Agric. 2013, 93, 2137–2145.
- Zeeman, S.C.; Thorneycroft, D.; Schupp, N.; Chapple, A.; Weck, M.; Dunstan, H.; Haldimann, P.; Bechtold, N.; Smith, A.M.; Smith, S. Plastidial α-Glucan Phosphorylase Is Not Required for Starch Degradation in Arabidopsis Leaves but Has a Role in the Tolerance of Abiotic Stress. Plant Physiol. 2004, 135, 849–858.
- Tetlow, I.J.; Bertoft, E. A Review of Starch Biosynthesis in Relation to the Building Block-Backbone Model. Int. J. Mol. Sci. 2020, 21, 7011.
- Roldán, I.; Wattebled, F.; Lucas, M.M.; Delvallé, D.; Planchot, V.; Jiménez, S.; Pérez, R.; Ball, S.; D’Hulst, C.; Mérida, Á. The phenotype of soluble starch synthase IV defective mutants of Arabidopsis thaliana suggests a novel function of elongation enzymes in the control of starch granule formation. Plant J. 2007, 49, 492–504.
- Leterrier, M.; Holappa, L.D.; Broglie, K.E.; Beckles, D.M. Cloning, characterisation and comparative analysis of a starch synthase IV gene in wheat: Functional and evolutionary implications. BMC Plant Biol. 2008, 8, 98.
- Roach, P.J.; DePaoli-Roach, A.A.; Hurley, T.; Tagliabracci, V.S. Glycogen and its metabolism: Some new developments and old themes. Biochem. J. 2012, 441, 763–787.
- Burrell, M.M. Starch: The need for improved quality or quantity—An overview. J. Exp. Bot. 2003, 54, 451–456.
- Tetlow, I.J.; Emes, M.J. A review of starch-branching enzymes and their role in amylopectin biosynthesis. IUBMB Life 2014, 66, 546–558.
- Crofts, N.; Nakamura, Y.; Fujita, N. Critical and speculative review of the roles of multi-protein complexes in starch biosynthesis in cereals. Plant Sci. 2017, 262, 1–8.
- Illingworth, B.; Brown, D.H.; Cori, C.F. The de novo synthesis of polysaccharide by phosphorylase. Proc. Natl. Acad. Sci. USA 1961, 47, 469–478.
- Tsai, C.Y.; Nelson, O.E. Two Additional Phosphorylases in Developing Maize Seeds. Plant Physiol. 1969, 44, 159–167.
- Grimaud, F.; Rogniaux, H.; James, M.G.; Myers, A.; Planchot, V. Proteome and phosphoproteome analysis of starch granule-associated proteins from normal maize and mutants affected in starch biosynthesis. J. Exp. Bot. 2008, 59, 3395–3406.
- Tetlow, I.J.; Beisel, K.G.; Cameron, S.; Makhmoudova, A.; Liu, F.; Bresolin, N.S.; Wait, R.; Morell, M.K.; Emes, M.J. Analysis of Protein Complexes in Wheat Amyloplasts Reveals Functional Interactions among Starch Biosynthetic Enzymes. Plant Physiol. 2008, 146, 1878–1891.
- Hwang, S.-K.; Nishi, A.; Satoh, H.; Okita, T. Rice endosperm-specific plastidial α-glucan phosphorylase is important for synthesis of short-chain malto-oligosaccharides. Arch. Biochem. Biophys. 2010, 495, 82–92.
- Ozbun, J.L.; Hawker, J.S.; Greenberg, E.; Lammel, C.; Preiss, J.; Lee, E.Y.C. Starch Synthetase, Phosphorylase, ADPglucose Pyrophosphorylase, and UDPglucose Pyrophosphorylase in Developing Maize Kernels. Plant Physiol. 1973, 51, 1–5.
- Dauvillée, D.; Chochois, V.; Steup, M.; Haebel, S.; Eckermann, N.; Ritte, G.; Ral, J.-P.; Colleoni, C.; Hicks, G.; Wattebled, F.; et al. Plastidial phosphorylase is required for normal starch synthesis in Chlamydomonas reinhardtii. Plant J. 2006, 48, 274–285.
- Nakamura, Y.; Ono, M.; Sawada, T.; Crofts, N.; Fujita, N.; Steup, M. Characterization of the functional interactions of plastidial starch phosphorylase and starch branching enzymes from rice endosperm during reserve starch biosynthesis. Plant Sci. 2017, 264, 83–95.
- Rathore, R.; Garg, N.; Garg, S.; Kumar, A. Starch phosphorylase: Role in starch metabolism and biotechnological applications. Crit. Rev. Biotechnol. 2009, 29, 214–224.
- Nakamura, Y.; Ono, M.; Utsumi, C.; Steup, M. Functional Interaction Between Plastidial Starch Phosphorylase and Starch Branching Enzymes from Rice During the Synthesis of Branched Maltodextrins. Plant Cell Physiol. 2012, 53, 869–878.
- Crofts, N.; Abe, N.; Oitome, N.F.; Matsushima, R.; Hayashi, M.; Tetlow, I.J.; Emes, M.J.; Nakamura, Y.; Fujita, N. Amylopectin biosynthetic enzymes from developing rice seed form enzymatically active protein complexes. J. Exp. Bot. 2015, 66, 4469–4482.
- Colleoni, C.; Dauvillée, D.; Mouille, G.; Morell, M.; Samuel, M.; Slomiany, M.-C.; Liénard, L.; Wattebled, F.; D’Hulst, C.; Ball, S. Biochemical Characterization of the Chlamydomonas reinhardtii α-1,4 Glucanotransferase Supports a Direct Function in Amylopectin Biosynthesis. Plant Physiol. 1999, 120, 1005–1014.
- Zeeman, S.C.; Delatte, T.; Messerli, G.; Umhang, M.; Stettler, M.; Mettler, T.; Streb, S.; Reinhold, H.; Kötting, O. Starch breakdown: Recent discoveries suggest distinct pathways and novel mechanisms. Funct. Plant Biol. 2007, 34, 465–473.
- Orzechowski, S. Starch metabolism in leaves. Acta Biochim. Pol. 2008, 55, 435–445.
- Fettke, J.; Eckermann, N.; Kötting, O.; Ritte, G.; Steup, M. Novel starch-related enzymes and carbohydrates. Cell. Mol. Boil. 2007, 52, 883–904.
- Mu, H.H.; Yu, Y.; Wasserman, B.P.; Carman, G.M. Purification and Characterization of the Maize Amyloplast Stromal 112-kDa Starch Phosphorylase. Arch. Biochem. Biophys. 2001, 388, 155–164.
- Mizuno, S.; Kamiyoshihara, Y.; Shiba, H.; Shinmachi, F.; Watanabe, K.; Tateishi, A. Plastidial starch phosphorylase is highly associated with starch accumulation process in developing squash (Cucurbita sp.) fruit. Physiol. Plant. 2018, 167, 264–275.
- Tiessen, A.; Nerlich, A.; Faix, B.; Hümmer, C.; Fox, S.; Trafford, K.; Weber, H.; Weschke, W.; Geigenberger, P. Subcellular analysis of starch metabolism in developing barley seeds using a non-aqueous fractionation method. J. Exp. Bot. 2012, 63, 2071–2087.
- Fettke, J.; Albrecht, T.; Hejazi, M.; Mahlow, S.; Nakamura, Y.; Steup, M. Glucose 1-phosphate is efficiently taken up by potato (Solanum tuberosum) tuber parenchyma cells and converted to reserve starch granules. N. Phytol. 2010, 185, 663–675.
- Malinova, I.; Mahlow, S.; Alseekh, S.; Orawetz, T.; Fernie, A.R.; Baumann, O.; Steup, M.; Fettke, J. Double Knockout Mutants of Arabidopsis Grown under Normal Conditions Reveal that the Plastidial Phosphorylase Isozyme Participates in Transitory Starch Metabolism. Plant Physiol. 2014, 164, 907–921.
- Jiang, J.; Yao, C.; Cao, X.; Liu, Y.; Xue, S. Characterization of starch phosphorylase from the marine green microalga (Chlorophyta) Tetraselmis subcordiformis reveals its potential role in starch biosynthesis. J. Plant Physiol. 2017, 218, 84–93.
- Fettke, J.; Leifels, L.; Brust, H.; Herbst, K.; Steup, M. Two carbon fluxes to reserve starch in potato (Solanum tuberosum L.) tuber cells are closely interconnected but differently modulated by temperature. J. Exp. Bot. 2012, 63, 3011–3029.
- Bustos, R.; Fahy, B.; Hylton, C.M.; Seale, R.; Nebane, N.M.; Edwards, A.; Martin, C.; Smith, A.M. Starch granule initiation is controlled by a heteromultimeric isoamylase in potato tubers. Proc. Natl. Acad. Sci. USA 2004, 101, 2215–2220.
- Tetlow, I.J.; Wait, R.; Lu, Z.; Akkasaeng, R.; Bowsher, C.G.; Esposito, S.; Kosar-Hashemi, B.; Morell, M.K.; Emes, M.J. Protein Phosphorylation in Amyloplasts Regulates Starch Branching Enzyme Activity and Protein–Protein Interactions. Plant Cell 2004, 16, 694–708.
- Subasinghe, R.M. Role and Regulation of Starch Phosphorylase and Starch Synthase IV in Starch Biosynthesis in Maize Endosperm Amyloplast. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2014.
- Young, G.-H.; Chen, H.-M.; Lin, C.-T.; Tseng, K.-C.; Wu, J.-S.; Juang, R.-H. Site-specific phosphorylation of L-form starch phosphorylase by the protein kinase activity from sweet potato roots. Planta 2005, 223, 468–478.
- Mori, H.; Tanizawa, K.; Fukui, T. A chimeric alpha-glucan phosphorylase of plant type L and H isozymes. Functional role of 78-residue insertion in type L isozyme. J. Biol. Chem. 1993, 268, 5574–5581.
- Liddle, A.M.; Manners, D.J.; Wright, A. Studies on carbohydrate-metabolizing enzymes. 6. The action of potato phosphorylase (P-enzyme) on starch-type polysaccharides. Biochem. J. 1961, 80, 304–309.
- Shimomura, S.; Nagai, M.; Fukui, T. Comparative Glucan Specificities of Two Types of Spinach Leaf Phosphorylase. J. Biochem. 1982, 91, 703–717.
- Young, N.D.; Debellé, F.; Oldroyd, G.E.D.; Geurts, R.; Cannon, S.B.; Udvardi, M.; Benedito, V.A.; Mayer, K.; Gouzy, J.; Schoof, H.; et al. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nat. Cell Biol. 2011, 480, 520–524.
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641.
- Chandler, V.L.; Brendel, V. The Maize Genome Sequencing Project. Plant Physiol. 2002, 130, 1594–1597.
- International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768.
- Nakano, K.; Mori, H.; Fukui, T. Molecular Cloning of cDNA Encoding Potato Amyloplast a-Glucan Phosphorylase and the Structure of Its Transit Peptide. J. Biochem. 1989, 106, 691–695.
- Camirand, A.; St-Pierre, B.; Marineau, C.; Brisson, N. Occurrence of a copia-like transposable element in one of the introns of the potato starch phosphorylase gene. Mol. Genet. Genom. 1990, 224, 33–39.
- Lin, C.-T.; Yeh, K.-W.; Lee, P.-D.; Su, J.-C. Primary Structure of Sweet Potato Starch Phosphorylase Deduced from its cDNA Sequence. Plant Physiol. 1991, 95, 1250–1253.
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.-K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J.; et al. Genome sequence of mungbean and insights into evolution within Vigna species. Nat. Commun. 2014, 5, 5443.
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964.
- Slugina, M.A.; Shchennikova, A.V.; Kochieva, E. The expression pattern of the Pho1a genes encoding plastidic starch phosphorylase correlates with the degradation of starch during fruit ripening in green-fruited and red-fruited tomato species. Funct. Plant Biol. 2019, 46, 1146.
- Walley, J.W.; Shen, Z.; Sartor, R.; Wu, K.J.; Osborn, J.; Smith, L.G.; Briggs, S.P. Reconstruction of protein networks from an atlas of maize seed proteotypes. Proc. Natl. Acad. Sci. USA 2013, 110, E4808–E4817.
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
14 Oct 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No