 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | John O'Doherty | + 2690 word(s) | 2690 | 2021-09-19 15:36:30 | | | |
| 2 | Conner Chen | Meta information modification | 2690 | 2021-10-08 09:01:31 | | |
Video Upload Options
Weaning is the most crucial event in commercial pig farms in terms of animal productivity and health. The newly weaned pig not only transits from milk to a solid and more complex diet, but is also subjected to additional stressors including separation from sow and littermates, co-mingling with unknown pigs, adaptation to new environmental settings, and increased pathogen exposure. All these stressors result in reduced feed intake, lasting up to 48 h post-weaning, which is the main driver of the observed gastrointestinal dysfunction, poor performance, and post-weaning diarrhoea (PWD). Marine polysaccharides from macroalgae and chitin provide an interesting source of novel bio-actives and are interesting group of natural dietary supplements for use in pig nutrition due to their prebiotic, antibacterial, and immunomodulatory activities. Hence, they offer great potential as preventatives and prophylactics in pig diets.
1. The Negative Biological Effects Associated with Weaning
2. Traditional and Alternative Dietary Interventions
3. Marine Polysaccharides
| Pig Age | Dietary Supplement | Dose | Time and Duration of Supplementation | Effect on Growth Performance and Diarrhoea Scores | Effect on Parameters of GIT Functionality and Health | Ref. |
|---|---|---|---|---|---|---|
| Weaned pigs | ||||||
| 24-day-old | Laminarin (Laminaria spp.) Fucoidan (Laminaria spp.) Laminarin + Fucoidan |
300 mg/kg 240 mg/kg 300 mg/kg + 240 mg/kg |
After weaning for 21 days | + ADG and G:F in pigs fed laminarin-supplemented diets + ADG in pigs fed with diet supplemented solely with fucoidan (interaction) − diarrhoea score in pigs fed laminarin-supplemented diets |
− faecal E. coli in pigs fed laminarin-supplemented diets + faecal Lactobacillus spp. in pigs fed with diet supplemented solely with fucoidan (interaction) |
[42] |
| 24-day-old | Laminarin (Laminaria spp.) Fucoidan (Laminaria spp.) Laminarin + Fucoidan |
150 or 300 mg/kg 240 mg/kg 150 or 300 mg/kg + 240 mg/kg |
After weaning for 35 days | + ADG in pigs fed 300 mg/kg laminarin-supplemented diets + G:F in pigs fed with diet supplemented solely with 300 mg/kg laminarin or fucoidan (interaction) − FS in pigs fed 150 or 300 mg/kg laminarin-supplemented diets and in pigs fed with diet supplemented solely with fucoidan (interaction) |
+ faecal Lactobacillus spp. in pigs fed fucoidan-supplemented diets 0 faecal E. coli, Bifidobacterium spp. |
[43] |
| 28-day-old | 65% laminarin-rich extract (Laminaria spp.) | 300 mg/kg | After weaning for 14 days | + ADG, ADFI 0 diarrhoea score |
+ VH in duodenum and jejunum and CD in jejunum − Enterobacteriaceae in caecum + Lactobacillus spp. in colon + butyrate in colon + gene expression of nutrient transporters in small intestine and colon − gene expression of tight junction proteins, mucins and immune markers in small intestine and colon |
[44] |
| 35-day-old | Dried seaweed (Ocean Harvest Technology) containing laminarin, fucoidan, alginate, mannitol, fucoxanthin and rhamnose sulphate. | 1500 mg/kg | After weaning for 52 days | 0 ADG, ADFI, G:F 0 diarrhoea score |
− VH in jejunum | [46] |
| 35-day-old | Dried sea weed (Ascophyllum nodosum) | 2.5 g/kg 5 g/kg 10 g/kg |
After weaning for 28 days | − ADG | ND | [48] |
| Finisher pigs | Dried seaweed extract (Ascophyllum nodosum) containing laminarin, fucoidan, alginate, mannitol, fucoxanthin and rhamnose sulphate. | 3 g/kg 6 g/kg 9 g/kg |
After weaning for 28 days | − ADG 0 ADFI, G:F |
ND | [49] |
| 28-day-old | 65% laminarin-rich extract (Laminaria spp.) | 300 mg/kg | After weaning for 14 days | + ADG, ADFI 0 diarrhoea score |
− abundance of OTUs assigned to Enterobacteriaceae + abundance of OTUs assigned to the genus Prevotella |
[57] |
| 24-day-old | Laminarin (Laminaria spp.) Fucoidan (Laminaria spp.) Laminarin + Fucoidan |
300 mg/kg 240 mg/kg 300 mg/kg + 240 mg/kg |
After weaning for 8 days | ND | − Enterobacteriaceae population in pigs offer fucoidan (interaction). − AEEC strains in pigs offer laminarin (interaction). + VH and VH:CD ratio in pigs offered laminarin or fucoidan (interaction). − IL-6, IL-17A and IL-1b mRNA expression in pigs offered laminarin |
[58] |
| 24-day-old | Laminarin (Laminaria spp.) | After weaning for 8 days | + ADG and ADFI − diarrhoea score |
ND | [59] | |
| 24-day-old | Laminarin (Laminaria spp.) |
0 mg/kg 240 mg/kg ZnO |
After weaning for 32 days | + ADG and G:F, similar effect to ZnO | + digestibility of GE + the expression of glucose transporters in small intestine compared with the basal diet. |
[60] |
| 24-day-old | 44% fucoidan-rich extract (Laminaria spp.) | 0 mg/kg 125 mg/kg 250 mg/kg |
After weaning for 14 days | − diarrhoea score 0 ADG, ADFI and G:F |
0 effect on VH − abundance of Prevotella and Lachnospiraceae + the abundance of Helicobacter |
[61] |
4. Laminarin
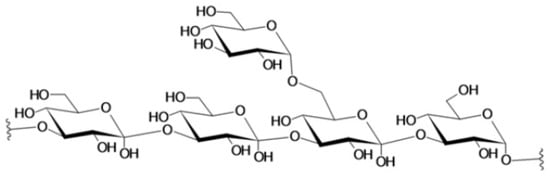
4.1. Antibacterial Activity
4.2. Prebiotic Activity
4.3. Immunomodulatory Activity
4.4. Effects of Laminarin-Rich Extracts on Pig GIT Functionality
References
- Lallès, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Weaning—A challenge to gut physiologists. Livest. Sci. 2007, 108, 82–93.
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236.
- Vente-Spreeuwenberg, M.A.; Verdonk, J.M.; Verstegen, M.W.; Beynen, A.C. Villus height and gut development in weaned piglets receiving diets containing either glucose, lactose or starch. Br. J. Nutr. 2003, 90, 907–913.
- Hedemann, M.S.; Højsgaard, S.; Jensen, B.B. Small intestinal morphology and activity of intestinal peptidases in piglets around weaning. J. Anim. Physiol. Anim. Nutr. 2003, 87, 32–41.
- Boudry, G.l.; Péron, V.; Le Huërou-Luron, I.; Lallès, J.P.; Sève, B. Weaning induces both transient and long-lasting modifications of absorptive, secretory, and barrier properties of piglet intestine. J. Nutr. 2004, 134, 2256–2262.
- Cera, K.R.; Mahan, D.C.; Cross, R.F.; Reinhart, G.A.; Whitmoyer, R.E. Effect of Age, Weaning and Postweaning Diet on Small Intestinal Growth and Jejunal Morphology in Young Swine. J. Anim. Sci. 1988, 66, 574–584.
- Montagne, L.; Boudry, G.; Favier, C.; Le Huerou-Luron, I.; Lalles, J.P.; Seve, B. Main intestinal markers associated with the changes in gut architecture and function in piglets after weaning. Br. J. Nutr. 2007, 97, 45–57.
- Spreeuwenberg, M.A.M.; Verdonk, J.M.A.J.; Gaskins, H.R.; Verstegen, M.W.A. Small intestine epithelial barrier function is compromised in pigs with low feed intake at weaning. J. Nutr. 2001, 131, 1520–1527.
- Bomba, L.; Minuti, A.; Moisa, S.J.; Trevisi, E.; Eufemi, E.; Lizier, M.; Chegdani, F.; Lucchini, F.; Rzepus, M.; Prandini, A.; et al. Gut response induced by weaning in piglet features marked changes in immune and inflammatory response. Funct. Integr. Genom. 2014, 14, 657–671.
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.; Blikslager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver. Physiol. 2010, 298, G352–G363.
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101.
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning anorexia may contribute to local inflammation in the piglet small intestine. J. Nutr. 1999, 129, 613–619.
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647.
- Pajarillo, E.A.B.; Chae, J.-P.; Balolong, M.P.; Kim, H.B.; Kang, D.-K. Assessment of fecal bacterial diversity among healthy piglets during the weaning transition. J. Gen. Appl. Microbiol. 2014, 60, 140–146.
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28.
- Chen, L.; Xu, Y.; Chen, X.; Fang, C.; Zhao, L.; Chen, F. The maturing development of gut microbiota in commercial piglets during the weaning transition. Front. Microbiol. 2017, 8, 1688.
- Wang, J.; Han, Y.; Meng, F.; Zhao, J.; Zhou, Z.; Fan, H. Fecal microbiota succession of piglets from birth to post-weaning by 454 pyrosequencing analysis. Trans. Tianjin Univ. 2017, 23, 211–220.
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54.
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.; Smidt, H.; de Vos, W.M. Post-natal development of the porcine microbiota composition and activities. Environ. Microbiol. 2006, 8, 1191–1199.
- Pieper, R.; Janczyk, P.; Zeyner, A.; Smidt, H.; Guiard, V.; Souffrant, W.B. Ecophysiology of the developing total bacterial and lactobacillus communities in the terminal small intestine of weaning piglets. Microb. Ecol. 2008, 56, 474–483.
- Urubschurov, V.; Janczyk, P.; Souffrant, W.B.; Freyer, G.; Zeyner, A. Establishment of intestinal microbiota with focus on yeasts of unweaned and weaned piglets kept under different farm conditions. FEMS Microbiol. Ecol. 2011, 77, 493–502.
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of early-life fecal microbiota in susceptible and healthy pigs to post-weaning diarrhoea. PLoS ONE 2017, 12, e0169851.
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873.
- Nagy, B.; Fekete, P.Z. Enterotoxigenic Escherichia coli (ETEC) in farm animals. Vet. Res. 1999, 30, 259–284.
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7.
- Kogan, G.; Kocher, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. Livest. Sci. 2007, 109, 161–165.
- Zanello, G.; Meurens, F.; Serreau, D.; Chevaleyre, C.; Melo, S.; Berri, M.; D’Inca, R.; Auclair, E.; Salmon, H. Effects of dietary yeast strains on immunoglobulin in colostrum and milk of sows. Vet. Immunol. Immunopathol. 2013, 152, 20–27.
- Castillo, M.; Martin-Orue, S.M.; Taylor-Pickard, J.A.; Perez, J.F.; Gasa, J. Use of mannanoligosaccharides and zinc chelate as growth promoters and diarrhea preventative in weaning pigs: Effects on microbiota and gut function. J. Anim. Sci. 2008, 86, 94–101.
- Searle, L.E.J.; Cooley, W.A.; Jones, G.; Nunez, A.; Crudgington, B.; Weyer, U.; Dugdale, A.H.; Tzortzis, G.; Collins, J.W.; Woodward, M.J.; et al. Purified galactooligosaccharide, derived from a mixture produced by the enzymic activity of Bifidobacterium bifidum, reduces Salmonella enterica serovar Typhimurium adhesion and invasion in vitro and in vivo. J. Med. Microbiol. 2010, 59, 1428–1439.
- Roselli, M.; Finamore, A.; Britti, M.S.; Bosi, P.; Oswald, I.; Mengheri, E. Alternatives to in-feed antibiotics in pigs: Evaluation of probiotics, zinc or organic acids as protective agents for the intestinal mucosa. A comparison of in vitro and in vivo results. Anim. Res. 2005, 54, 203–218.
- Stensland, I.; Kim, J.C.; Bowring, B.; Collins, A.M.; Mansfield, J.P.; Pluske, J.R. A comparison of diets supplemented with a feed additive containing organic acids, cinnamaldehyde and a permeabilizing complex, or zinc oxide, on post-weaning diarrhoea, selected bacterial populations, blood measures and performance in weaned pigs experimentally infected with Enterotoxigenic, E. coli. Animals 2015, 5, 1147–1168.
- Valeriano, V.D.; Balolong, M.P.; Kang, D.K. Probiotic roles of Lactobacillus sp. in swine: Insights from gut microbiota. J. Appl. Microbiol. 2017, 122, 554–567.
- Torrallardona, D. Spray Dried Animal Plasma as an Alternative to Antibiotics in Weanling Pigs—A Review. Asian-Aust. J. Anim. Sci. 2010, 23, 131–148.
- Torres-Pitarch, A.; Hermans, D.; Manzanilla, E.G.; Bindelle, J.; Everaert, N.; Beckers, Y.; Torrallardona, D.; Bruggeman, G.; Gardiner, G.E.; Lawlor, P.G. Effect of feed enzymes on digestibility and growth in weaned pigs: A systematic review and meta-analysis. Anim. Feed Sci. Technol. 2017, 233, 145–159.
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential oil and aromatic plants as feed additives in non-ruminant nutrition: A review. J. Anim. Sci. Biotechnol. 2015, 6, 7.
- Pluske, J.R. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1–7.
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597.
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. Central and storage carbon metabolism of the brown alga Ectocarpus siliculosus: Insights into the origin and evolution of storage carbohydrates in Eukaryotes. New Phytol. 2010, 188, 67–81.
- Wang, D.; Kim, D.H.; Kim, K.H. Effective production of fermentable sugars from brown macroalgae biomass. Appl. Microbiol. Biotechnol. 2016, 100, 9439–9450.
- Overland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24.
- Corino, C.; Modina, S.C.; Di Giancamillo, A.; Chiapparini, S.; Rossi, R. Seaweeds in Pig Nutrition. Animals 2019, 9, 1126.
- McDonnell, P.; Figat, S.; O’Doherty, J.V. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance, selected faecal microbial populations and volatile fatty acid concentrations. Animal 2010, 4, 579–585.
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; ’Doherty, J.V.O. Effect of supplementing varying inclusion levels of laminarin and fucoidan on growth performance, digestibility of diet components, selected faecal microbial populations and volatile fatty acid concentrations in weaned pigs. Anim. Feed Sci. Technol. 2013, 183, 151–159.
- Rattigan, R.; Sweeney, T.; Maher, S.; Thornton, K.; Rajauria, G.; O’Doherty, J.V. Laminarin-rich extract improves growth performance, small intestinal morphology, gene expression of nutrient transporters and the large intestinal microbial composition of piglets during the critical post-weaning period. Br. J. Nutr. 2020, 123, 255–263.
- Rattigan, R.; Sweeney, T.; Maher, S.; Ryan, M.T.; Thornton, K.; O’Doherty, J.V. Effects of reducing dietary crude protein concentration and supplementation with either laminarin or zinc oxide on the growth performance and intestinal health of newly weaned pigs. Anim. Feed Sci. Technol. 2020, 270, 114693.
- Satessa, G.D.; Kjeldsen, N.J.; Mansouryar, M.; Hansen, H.H.; Bache, J.K.; Nielsen, M.O. Effects of alternative feed additives to medicinal zinc oxide on productivity, diarrhoea incidence and gut development in weaned piglets. Animal 2020, 14, 1638–1646.
- Dierick, N.; Ovyn, A.; De Smet, S. Effect of feeding intact brown seaweed Ascophyllum nodosum on some digestive parameters and on iodine content in edible tissues in pigs. J. Sci. Food Agric. 2009, 89, 584–594.
- Michiels, J.; Skrivanova, E.; Missotten, J.; Ovyn, A.; Mrazek, J.; De Smet, S.; Dierick, N. Intact brown seaweed (Ascophyllum nodosum) in diets of weaned piglets: Effects on performance, gut bacteria and morphology and plasma oxidative status. J. Anim. Physiol. Anim. Nutr. 2012, 96, 1101–1111.
- Gardiner, G.E.; Campbell, A.J.; O’Doherty, J.V.; Pierce, E.; Lynch, P.B.; Leonard, F.C.; Stanton, C.; Ross, R.P.; Lawlor, P.G. Effect of Ascophyllum nodosum extract on growth performance, digestibility, carcass characteristics and selected intestinal microflora populations of grower–finisher pigs. Anim. Feed Sci. Technol. 2008, 141, 259–273.
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675.
- Perez, M.J.; Falque, E.; Dominguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52.
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020.
- Garcia-Vaquero, M.; Rajauria, G.; Miranda, M.; Sweeney, T.; Lopez-Alonso, M.; O’Doherty, J. Seasonal variation of the proximate composition, mineral content, fatty acid profiles and other phytochemical constituents of selected brown macroalgae. Mar. Drugs 2021, 19, 204.
- Shannon, E.; Abu-Ghannam, N. Antibacterial derivatives of marine algae: An overview of pharmacological mechanisms and applications. Mar. Drugs 2016, 14, 81.
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: applications and mode of action. Biomacromolecules 2003, 4, 1457–1465.
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food. Microbiol. 2010, 144, 51–63.
- Vigors, S.; O’Doherty, J.V.; Rattigan, R.; McDonnell, M.J.; Rajauria, G.; Sweeney, T. Effect of a laminarin rich macroalgal extract on the caecal and colonic microbiota in the post-weaned pig. Mar. Drugs 2020, 18, 157.
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of dietary laminarin and fucoidan on selected microbiota, intestinal morphology and immune status of the newly weaned pig. Br. J. Nutr. 2013, 110, 1630–1638.
- Bouwhuis, M.A.; Sweeney, T.; Mukhopadhya, A.; Thornton, K.; McAlpine, P.O.; O’Doherty, J.V. Zinc methionine and laminarin have growth-enhancing properties in newly weaned pigs influencing both intestinal health and diarrhoea occurrence. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1273–1285.
- Heim, G.; Walsh, A.M.; Sweeney, T.; Doyle, D.N.; O’Shea, C.J.; Ryan, M.T.; O’Doherty, J.V. Effect of seaweed-derived laminarin and fucoidan and zinc oxide on gut morphology, nutrient transporters, nutrient digestibility, growth performance and selected microbial populations in weaned pigs. Br. J. Nutr. 2014, 111, 1577–1585.
- Rattigan, R.; Sweeney, T.; Vigors, S.; Thornton, K.; Rajauria, G.; O’Doherty, A.J.V. The effect of increasing inclusion levels of a fucoidan-rich extract derived from Ascophyllum nodosum on growth performance and aspects of intestinal health of pigs post-weaning. Mar. Drugs 2019, 17, 680.
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31.
- Graiff, A.; Ruth, W.; Kragl, U.; Karsten, U. Chemical characterization and quantification of the brown algal storage compound laminarin—A new methodological approach. J. Appl. Phycol. 2016, 28, 533–543.
- Adams, J.M.; Ross, A.B.; Anastasakis, K.; Hodgson, E.M.; Gallagher, J.A.; Jones, J.M.; Donnison, I.S. Seasonal variation in the chemical composition of the bioenergy feedstock Laminaria digitata for thermochemical conversion. Bioresour. Technol. 2011, 102, 226–234.
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373.
- Kadam, S.U.; O’Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound assisted extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280.
- Liu, Z.; Xiong, Y.; Yi, L.; Dai, R.; Wang, Y.; Sun, M.; Shao, X.; Zhang, Z.; Yuan, S. Endo-beta-1,3-glucanase digestion combined with the HPAEC-PAD-MS/MS analysis reveals the structural differences between two laminarins with different bioactivities. Carbohydr. Polym. 2018, 194, 339–349.
- Sellimi, S.; Maalej, H.; Rekik, D.M.; Benslima, A.; Ksouda, G.; Hamdi, M.; Sahnoun, Z.; Li, S.; Nasri, M.; Hajji, M. Antioxidant, antibacterial and in vivo wound healing properties of laminaran purified from Cystoseira barbata seaweed. Int. J. Biol. Macromol. 2018, 119, 633–644.
- Smith, A.G.; O’Doherty, J.V.; Reilly, P.; Ryan, M.T.; Bahar, B.; Sweeney, T. The effects of laminarin derived from Laminaria digitata on measurements of gut health: Selected bacterial populations, intestinal fermentation, mucin gene expression and cytokine gene expression in the pig. Br. J. Nutr. 2011, 105, 669–677.
- Sweeney, T.; Collins, C.B.; Reilly, P.; Pierce, K.M.; Ryan, M.; O’Doherty, J.V. Effect of purified beta-glucans derived from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae on piglet performance, selected bacterial populations, volatile fatty acids and pro-inflammatory cytokines in the gastrointestinal tract of pigs. Br. J. Nutr. 2012, 108, 1226–1234.
- Murphy, P.; Dal Bello, F.; O’Doherty, J.; Arendt, E.K.; Sweeney, T.; Coffey, A. Analysis of bacterial community shifts in the gastrointestinal tract of pigs fed diets supplemented with beta-glucan from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae. Animal 2013, 7, 1079–1087.
- Lynch, M.B.; Sweeney, T.; Callan, J.J.; O’Sullivan, J.T.; O’Doherty, J.V. The effect of dietary Laminaria-derived laminarin and fucoidan on nutrient digestibility, nitrogen utilisation, intestinal microflora and volatile fatty acid concentration in pigs. J. Sci. Food Agric. 2010, 90, 430–437.
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The effects of the marine-derived polysaccharides laminarin and chitosan on aspects of colonic health in pigs challenged with dextran sodium sulphate. Mar. Drugs 2020, 18, 262.
- O’Shea, C.J.; O’Doherty, J.V.; Callanan, J.J.; Doyle, D.; Thornton, K.; Sweeney, T. The effect of algal polysaccharides laminarin and fucoidan on colonic pathology, cytokine gene expression and Enterobacteriaceae in a dextran sodium sulfate-challenged porcine model. J. Nutr. Sci. 2016, 5, e15.
- Ryan, M.T.; O’Shea, C.J.; Collins, C.B.; O’Doherty, J.V.; Sweeney, T. Effects of dietary supplementation with Laminaria hyperborea, Laminaria digitata, and Saccharomyces cerevisiae on the IL-17 pathway in the porcine colon. J. Anim. Sci. 2012, 90, 263–265.


