 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Victor Benno Meyer-Rochow | + 5544 word(s) | 5544 | 2021-09-07 04:42:57 | | | |
| 2 | Vivi Li | Meta information modification | 5544 | 2021-09-18 08:35:29 | | |
Video Upload Options
The ability to repair injuries among reptiles, i.e., ectothermic amniotes, is similar to that of mammals with some noteworthy exceptions. While large wounds in turtles and crocodilians are repaired through scarring, the reparative capacity involving the tail derives from a combined process of wound healing and somatic growth, the latter being continuous in reptiles. When the tail is injured in juvenile crocodilians, turtles and tortoises as well as the tuatara (Rhynchocephalia: Sphenodon punctatus, Gray 1842), the wound is repaired in these reptiles and some muscle and connective tissue and large amounts of cartilage are regenerated during normal growth. This process, here indicated as “regengrow”, can take years to produce tails with similar lengths of the originals and results in only apparently regenerated replacements.
1. Introduction and Overview
1.1. Wound Healing and Regeneration among Reptiles Generally
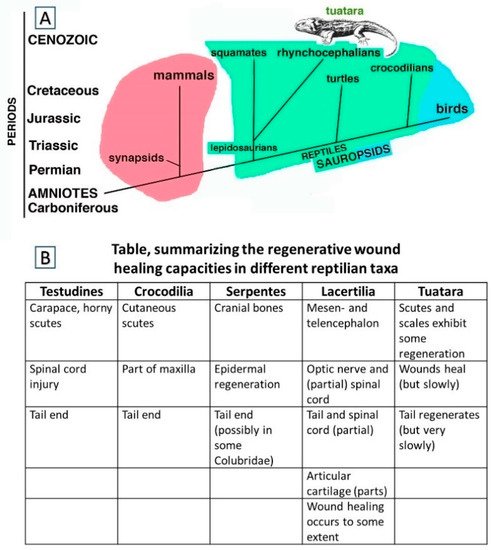
1.2. Focusing on Tail Autotomies in Reptiles
2. Caudal Autotomy in Tuatara
2.1. Sample Acquisition and Methodology
2.2. Tail Regeneration in the Tuatara Represents a Case of Regengrow
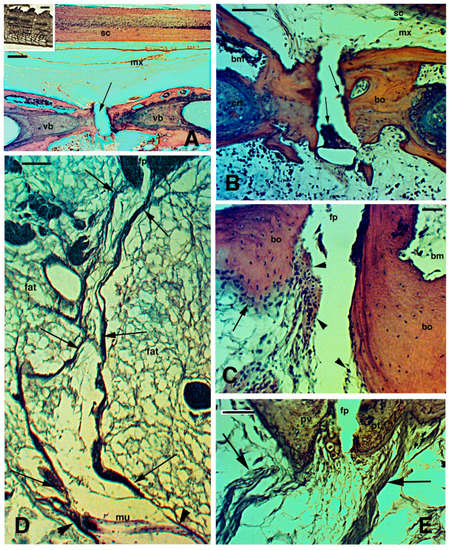
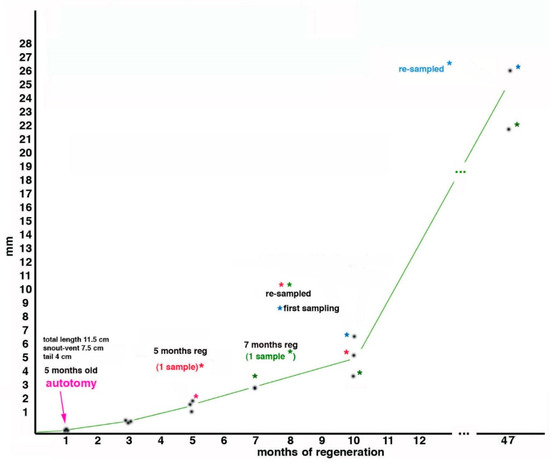
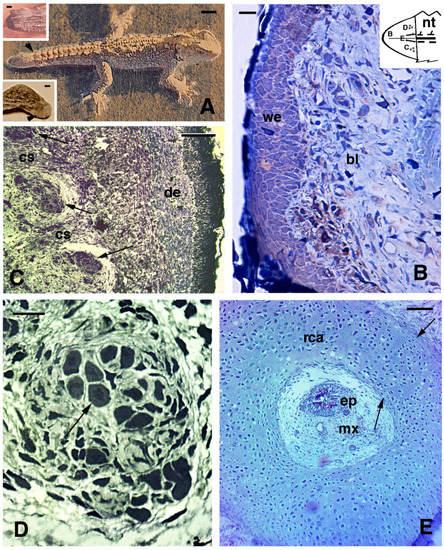
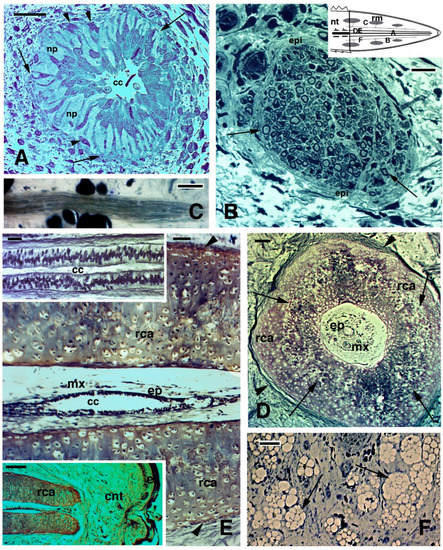
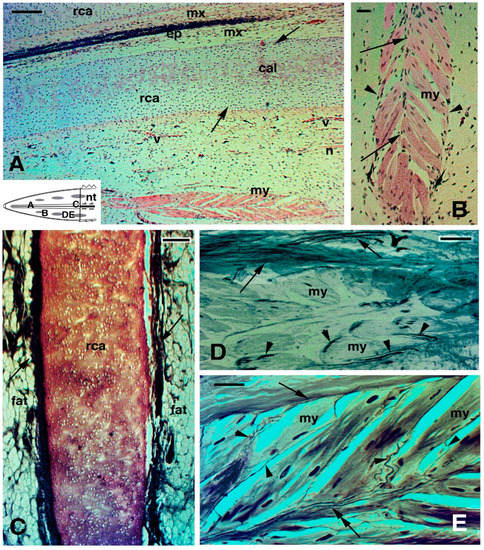
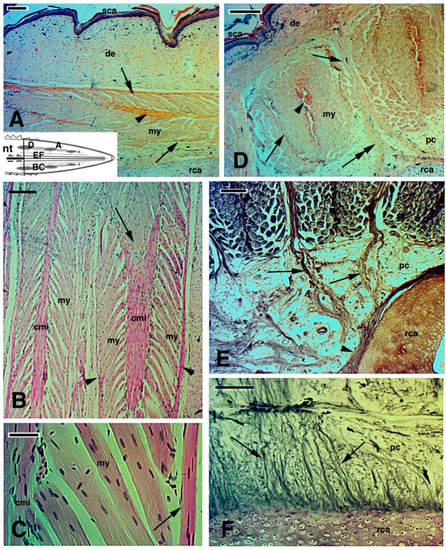
2.3. Histology of Regenerating and Regenerated Tails in the Tuatara
References
- Panini, G.P. The Prehistoric World; Treasure Press: London, UK, 1987.
- Pough, H.F.; Janis, C.M.; Heiser, J.B. Vertebrate Life; Pearson Benjamin Cummings: San Francisco, CA, USA, 2009.
- Alibardi, L. Review: Tail regeneration in lepidosauria as an exception to the generalized lack of organ regeneration in amniotes. J. Exp. Zool. B 2020, 336, 145–165.
- Evans, S.E.; Jones, M.E.H. The origin, early history and diversification of lepidosauromorph reptiles. In New Aspects of Mesozoic Biodiversity Lecture Notes in Earth Sciences (132); Bandyopadhyay, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 27–44.
- Ferguson, M.W.J.; O’Kane, S. Scar-free healing: From embryonic mechanisms to adult 235 therapeutic intervention. Philos. Trans. R. Soc. Lond. 2004, 359B, 839–850.
- Gadow, H. On the reproduction of the carapax in tortoises. J. Anat. 1886, 20, 220–224.
- Howey, C.A.F.; Roosenburg, W.M. Effects of prescribed fire on the eastern box turtle (Terrapene carolina carolina). North East. Nat. 2013, 20, 493–497.
- Negrini, J.; Ginel, P.J.; Novales, M.; Guerra, R.; Mozos, E. Clinical and histological findings of cutaneous wound healing in the red-eared slider turtle (Trachemys scripts elegans) housed in unheated outdoor enclosures. Vet. Dermatol. 2016, 27, 413-e106.
- Smith, H.M. Total regeneration of the carapace in a box turtle. Turtox News 1958, 36, 234–238.
- Kuchling, G. Restoration of epidermal scute patterns during regeneration of the chelonian carapace. Chelonian Conserv. Biol. 1997, 2, 500–506.
- Flemming, G.J. Clinical technique: Chelonian shell repair. J. Exot. Pet. Med. 2008, 17, 246–258.
- Sos, T. Two extreme cases of regeneration in Testudo graeca ibera Pallas 1814. Biharean Biologist 2012, 6, 128–131.
- Davenport, J. Regeneration of the tail spur in Testudo hermanni. Testudo 1995, 4, 79–80.
- Kuchling, G. Bifid tail regeneration in a turtle, Emydura sp. (Testudines:Chelidae). Chelonian Conserv. Biol. 2005, 4, 935–939.
- Martinez-Silvestre, A.; Soler-Massana, J. Regeneracion del caparazon en Testudo hermanni hermanni despues de un incendio forestal. Biol. Asoc. Herpetol. Esp. 2000, 11, 90–92.
- Mota-Rodrigues, J.E.; Feitosa-Silva, J.R. Phrynops tuberosus (Cotinga River toad-headed turtle); bifid tail. Herpetol. Rev. 2013, 44, 308–309.
- Reherman, M.I.; Marichal, N.; Russo, R.E.; Trujillo-Cenoz, O. Neural reconnection in the transected spinal cord of the freshwater turtle Trachemys dorbignyi. J. Comp. Neurol. 2009, 515, 197–214.
- Reherman, M.I.; Santinaque, F.F.; Lopez-Carro, B.; Russo, R.E.; Trujillo-Cenoz, O. Cell proliferation and cytoarchitectural remodeling during spinal cord reconnection in the fresh-water turtle Trachemys dorbignyi. Cell Tissue Res. 2011, 344, 415–433.
- Brazaitis, P. Maxillary regeneration in marsh crocodile, Crocodilus palustris. J. Herpetol. 1981, 15, 360–362.
- Kälin, J.A. Über Skeletalanomalien bei Crocodiliden. Zeitschrift fuer Morphol. OekolgieTiere 1937, 32, 327–347.
- Rashid, D.J.; Chapman, S.C. The long and the short of tails. Dev. Dyn. 2021, in press.
- Dathe, H. Schwanz-Regeneration beim Brillenkaiman. Natur Volk. 1960, 90, 289–292.
- Xu, C.; Palade, J.; Fisher, R.E.; Smith, C.I.; Clark, A.R.; Sampson, S.; Bourgeois, R.; Rawls, A.; Elsey, R.M.; Wilson-Rawls, J.; et al. Anatomical and histological analyses reveal that tail repair is coupled with regrowth in wild-caught, juvenile American alligators (Alligator mississippiensis). Sci. Rep. 2020, 10, 20122.
- Alibardi, L. Regeneration in anamniotes was replaced by regengrow and scarring in amniotes after land colonization and the evolution of terrestrial biological cycles. Dev. Dyn. 2021.
- Smith, D.A.; Barker, I.K. Healing of cutaneous wounds in the common garter snake (Thamnnophis sirtalis). Can. J. Vet. Res. 1988, 52, 111–119.
- Henle, K.; Grimm-Seyfarth, A. Exceptional occurrences of double, triple and quintuple tails in an Australian lizard community, with a review of supernumerary tails in natural populations of reptiles. Salamandra 2020, 56, 373–391.
- Redi, F. Osservazione intorno agli animali viventi che si trovano negli animali viventi; Lion d’Oro: Firenze, Italy, 1684; (cited in Henle & Grimm-Seyfahrt, 2020).
- Wallach, V. Axial bifurcation and duplication in snakes. Pt 1: A synopsis of authentic and anecdotal cases. Bull. Md. Herpetol. Soc. 2007, 43, 57–95, (cited in Henle & Grimm-Seyfahrt, 2020).
- Costa, H.C.; Moura, M.R.; Feio, R.N. A tale of lost tails: Pseudoautotomy in the neotropical snake genus Drymoliber (Serpentes; Colubridae). Can. J. Zool. 2014, 92, 811–816.
- Gans, C. The characteristics and affinities of the Amphisbaenia. Trans. Zool. Soc. Lond. 1978, 34, 347–416.
- Guedes, J.J.M.; Costa, H.C.; Moura, M.R. A new tale of lost tails: Correlates of tail breakage in the worm lizard Amphisbaena vermicularis. Ecol. Evol. 2020, 10, 14247–14255.
- Bailleul, A.M.; Hall, B.K.; Horner, J.R. Secondary cartilage revealed in a non-avian dinosaur embryo. PLoS ONE 2013, 8, e56937.
- Irwin, C.R.; Ferguson, M.W.J. Fracture repair of reptilian dermal bones: Can reptiles form secondary cartilage? J. Anat. 1986, 146, 53–64.
- LeBlanc, A.R.H.; MacDougall, M.J.; Haridy, Y.; Scott, D.; Reisz, R.R. Caudal autotomy as anti-predatory behavior in palaeozoic reptiles. Sci. Rep. 2018, 8, 3328.
- Bellairs, A.A.; Bryant, S.V. Autotomy and regeneration in reptiles. In Biology of the Reptilia; Gans, C., Billet, F., Maderson, P.F.A., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 302–410.
- Arnold, E.N. Evolutionary aspects of tail shedding in lizards and their relatives. J. Nat. Hist. 1984, 18, 127–169.
- Maginnis, T.L. The cost of autotomy and regeneration in animals: A review and framework for future research. Behav. Ecol. 2006, 17, 857–872.
- Dunoyer, L.A.; Seifert, A.W.; Van Cleve, J. Evolutionary bedfellows: Reconstructing the ancestral state of autotomy and regeneration. J. Exp. Zool. Mol. Dev. Evol. 2020, 336, 94–115.
- Gordeev, D.A.; Ananjeva, N.B.; Korost, D.V. Autotomy and regeneration in squamate reptiles (Squamata, Reptilia): Defensive behavior strategies and morphological characteristics (using computer microtomography methods). Biol. Bull. 2020, 47, 389–398.
- Werber, I. Regeneration der Kiefer bei der Eidechse Lacerta agilis. Wilhelm Roux’s Arch. Entwickl. Mechnaik 1905, 19, 248–258.
- Peacock, H.M.; Gilbert, E.A.; Vickaryous, M.K. Scar-free cutaneous wound healing in the leopard gecko, Eublepharis macularius. J. Anat. 2015, 227, 596–610.
- Alibardi, L.; Meyer-Rochow, V.B. Regeneration of adhesive tail pad scales in the New Zealand gecko (Hoplodactylus maculatus) (Reptilia; Squamata; Lacertilia) can serve as an experimental model to analyze setal formation in lizards generally. Zool. Res. 2017, 38, 191–197.
- Jacyniak, K.; Vickaryous, M.K. Constitutive cardiomyocyte proliferation in the leopard gecko (Eublepharis macularius). J. Morphol. 2018, 279, 1355–1367.
- Crnobrnja-Isailoic, J.; Corovic, J.; Halpern, B. Deliberate tail loss in Dolichophis caspius and Natrix tesselata (Serpentes: Colubridae) with a brief review of pseudoautotomy in contemporary snake families. North West. J. Zool. 2016, 12, 367–372.
- Ananjeva, N.B.; Orlov, N.I. Caudal autotomy in colubrid snake Xenochrophis piscator from Vietnam. Russ. J. Herpetol. 1994, 1, 169–171.
- Broadley, D.G. Caudal autotomy in African snakes of the genera Natriciteres Loveridge and Psammophis Boie. J. Herpetol. Assoc. Afr. 1987, 33, 18–19.
- Cooper, W.E., Jr.; Alfieri, K.J. Caudal autotomy in the eastern garter snake, Thamnophis s. sirtalis. Amphib. Reptil. 1993, 14, 86–89.
- Loveridge, A. Revision of the African snakes of the genera Dromophis and Psammophis. Bull. Mus. Comp. Zool. Harv. Coll. 1940, 87, 1–69.
- Sharma, B.D. A rare case of autotomy seen in Amphiesma stolata (Linn., Serpentes, Colubridae). Snake 1980, 12, 60.
- Mello, R.S.R.; Besson, A.A.; Hare, K.M.; Fay, V.; Smith, E.; Cree, A. Adjustment of juvenile tuatara to a cooler, southern climate: Operative temperatures, emergence behaviour, and growth rate. N. Z. J. Zool. 2013, 40, 290–303.
- Cree, A. Tuatara: Biology and conservation of a venerable survivor; Canterbury University Press: Christchurch, New Zealand, 2014.
- Alibardi, L.; Meyer-Rochow, V.B. Microscopical observations on the regenerating tail in the tuatara Sphenodon punctatus indicate a tendency to scarring, but also influence from somatic growth. J. Morphol. 2019, 280, 411–422.
- Batemen, P.W.; Fleming, P.A. To cut a long tail short: A review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool. 2009, 277, 1–14.
- Woodland, W.N.F. Some observations on caudal autotomy and regeneration in the gecko (Hemidactylus flaviviridis, Ruppel), with notes on the tails of Sphenodon and Pygopus. Q. J. Microsc. Sci. 1920, 65, 63–100.
- Ali, S.M. Studies on the comparative anatomy of the tail in sauria and rhynchocephalian Sphenodon punctatus Gray. Proc. Indian Acad. Sci. 1941, B13, 171–192.
- Seligmann, H.; Moravec, J.; Werner, Y.L. Morphological, functional and evolutionary aspects of tail autotomy and regeneration in the ‘living fossil’ Sphenodon (Reptilia, Rhynchocephalia). Biol. J. Linn. Soc. 2008, 93, 721–743.
- Quattrini, D. Piano di autotomia e rigenerazione della coda nei sauri. Arch. Ital. Anat. Embriol. 1954, 59, 225–282.
- Alibardi, L.; Meyer-Rochow, V.B. Ultrastructural survey of the spinal cord of young tuataras (Sphenodon punctatus) with emphasis on the glia. N. Z. J. Zool. 1990, 17, 73–85.
- Alibardi, L.; Meyer-Rochow, V.B. Comparative fine structure of the axial skeleton inside the regenerated tail of lizards and the tuatara (Sphenodon punctatus). Gegenbaurs Morphol. Jahrb. (Leipzig) 1989, 135, 705–716.
- Barr, J.I.; Boisvert, C.A.; Somaweera, R.; Trinajstic, K.; Bateman, P.W. Re-regeneration to reduce negative effects associated with tail loss in lizards. Sci. Rep. 2019, 9, 18717.
- Nelson, N.J.; Thompson, M.B.; Pledger, S.; Keall, S.N.; Daugherty, C.H. Egg mass determines hatchling size, and incubation temperature influences post-hatching growth, of tuatara Sphenodon punctatus. J. Zool. 2004, 263, 77–87.
- Alibardi, L.; Meyer-Rochow, V.B. Fine structure of regenerating caudal spinal cord in the tuatara (Sphenodon punctatus). J. Hirnforsch. 1990, 31, 613–621.
- Worthy, T.H.; Holdaway, R.N. The Lost World of the Moa: Prehistoric Life of New Zealand; Indiana University Press: Bloomington, IN, USA, 2002; p. 718. ISBN 13-978-0253340344.
- Meyer-Rochow, V.B.; Wohlfahrt, S.; Ahnelt, P.K. Photoreceptor cell types in the retina of the tuatara (Sphenodon punctatus) have cone characteristics. Micron 2005, 36, 423–428.
- Meyer-Rochow, V.B. Observations on the behaviour of young tuatara Sphenodon punctatus in conditions of total darkness. Tuatara 1988, 30, 36–38.
- Meyer-Rochow, V.B.; The, K.L. Visual predation by tuatara (Sphenodon punctatus) on the beach beetle Chaerodestrachyscelides as a selective force in the production of distinct colour morphs. Tuatara 1991, 31, 1–8.
- Alibardi, L.; Meyer-Rochow, V.B. General and specific microscopic characteristics of the dorsal tail scales and the spines of the crest in the tuatara Sphenodon pucntatus (Reptilia; Rhynchocephalia; Sphenodontidae). Micron 2020, 137, 102909.
- Alibardi, L. Electron microscopic observations on the myelination of the long-term regenerated caudal spinal cord in lizards and Sphenodon. Biol. Struct. Morphog. 1990, 3, 147–158.
- Alibardi, L.; Maderson, P.F.A. Observations on the histochemistry and ultrastructure of the epidermis of the tuatara, Sphenodon punctatus (Sphenodontida, Lepidosauri, Reptiles): A contribution to an understanding of the lepidosaurian epidermal generation and the evolutionary origin of the squamate shedding complex. J. Morphol. 2003, 256, 111–133.
- Alibardi, L.; Maderson, P.F.A. Observations on the histochemistry and ultrastructure of regenerating caudal epidermis of the tuatara Sphenodon punctatus (Sphenodontida, Lepidosauria, Reptilia). J. Morphol. 2003, 256, 134–145.

