Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hanaa Adsi | + 1657 word(s) | 1657 | 2021-09-01 10:44:52 | | | |
| 2 | Rita Xu | -59 word(s) | 1598 | 2021-09-15 03:57:27 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Adsi, H. Chemical Chaperones. Encyclopedia. Available online: https://encyclopedia.pub/entry/14183 (accessed on 25 July 2026).
Adsi H. Chemical Chaperones. Encyclopedia. Available at: https://encyclopedia.pub/entry/14183. Accessed July 25, 2026.
Adsi, Hanaa. "Chemical Chaperones" Encyclopedia, https://encyclopedia.pub/entry/14183 (accessed July 25, 2026).
Adsi, H. (2021, September 14). Chemical Chaperones. In Encyclopedia. https://encyclopedia.pub/entry/14183
Adsi, Hanaa. "Chemical Chaperones." Encyclopedia. Web. 14 September, 2021.
Copy Citation
Chemical chaperones are well known as inhibitors of protein and peptide self-assembly, as well as structural stabilizers of misfolded enzymes. We discovered that chemical chaperones inhibit adenine self-assembly both in a yeast model and in vitro. According to our study, chemical chaperones might have a dual function as enhancers of enzyme stability and as inhibitors of toxic metabolite self-assembly. This novel approach could be implemented for the rational design of inhibitors that target metabolite self-assembly as therapeutics for inborn errors of metabolism.
chemical chaperones
osmolytes
hydrophobic compounds
metabolite assemblies
amyloid formation
1. Introduction
Proteins are highly functionally and structurally diverse biomolecules. To function properly, proteins usually obtain a specific conformational state that is considered ‘native’. However, the folded state of proteins is often only marginally stable [1][2][3]. Certain genetic, physiological, and environmental conditions can induce protein misfolding, resulting in the formation of small aggregation-prone species that tend to self-assemble into supramolecular assemblies [4]. In some cases, these assemblies obtain a fibrillar morphology and a set of canonical biophysical and biochemical traits, which are the hallmarks of amyloids [4][5]. Amyloid formation is associated with a wide range of disorders, such as type 2 diabetes, Alzheimer’s disease, and Parkinson’s disease, and is thus considered a central target for therapeutic intervention [4].
Chaperones play a pivotal role as the safeguards of the protein quality control system and take part in enhancing the proper folding of proteins, thus counteracting the potentially deleterious effect of protein misfolding [6]. They are classified into three main groups: molecular, pharmacological, and chemical chaperones [7]. Molecular chaperones (such as heat shock proteins) facilitate the proper folding of other proteins and are central players in the proteostasis network [6][7][8][9]. Pharmacological chaperones are small molecules that specifically bind misfolded proteins and induce refolding or structure stabilization [10]. Unlike pharmacological chaperones, chemical chaperones are small molecules that do not bind a single misfolded protein-ligand but rather function in a non-specific manner [10]. Chemical chaperones are divided into two structurally and functionally distinct groups: osmolytes and hydrophobic compounds [11]. Osmolytes are low molecular weight compounds such as amino acids, polyols, and methylamines (e.g., glycine, glycerol, dimethyl sulfoxide (DMSO), and trimethylamine N-oxide (TMAO)). Osmolytes increase the stability of the folded state by sequestering water molecules, thereby generating a preferential depletion of the osmolyte adjacent to the protein surface, leading to protein stabilization and conformational changes [12]. Hydrophobic compounds include several bile acids (e.g., deoxycholic acid (DCA) and ursodeoxycholic acid (UDCA)). Their mode of action is postulated to include direct interactions with the hydrophobic regions of misfolded proteins, thus favoring the folded form. However, the exact mechanism of action of this group is still not fully understood [11].
Chemical chaperones were shown to reduce endoplasmic reticulum stress, correct the misfolded form of the p53 tumor suppressor protein and restore glucose homeostasis [13][14][15][16][17]. The effect of different chemical chaperones in ameliorating protein misfolding in neurodegenerative and prion diseases has been well documented both in vitro and in vivo [18][19][20][21][22][23][24][25]. Chemical chaperones thus could potentially serve as scaffolds for the development of novel therapeutics for protein misfolding diseases.
A reductionist approach has allowed to re-define the minimal building block required for amyloid formation, demonstrating that this phenomenon is not restricted to proteins and peptides. Specifically, we have established that metabolites such as amino acids, nucleobases, and other small molecules can self-assemble into well-ordered amyloid-like fibrillar structures that share many common biophysical and biochemical properties with classical protein amyloids (Figure 1A). The extension of the canonical amyloid hypothesis has provided a novel paradigm for the etiology of human inborn errors of metabolism (IEMs), wherein a congenital enzymatic deficiency results in the accumulation of one or more metabolites [26]. We have recently established a salvage yeast model for adenine accumulation and self-assembly by recapitulating the enzymatic deficiency of patients suffering from metabolic disorders associated with the accumulation of adenine and its derivatives [27]. This model demonstrates robust sensitivity to adenine that is attributed to the formation of toxic adenine amyloid-like supramolecular structures, as indicated by staining with an amyloid-specific dye and specific antibodies raised against the fibrils. These effects were decreased upon treatment with generic fibrillation-modifying polyphenolic compounds such as tannic acid, thus supporting the potential of the model as a drug discovery platform [27][28].
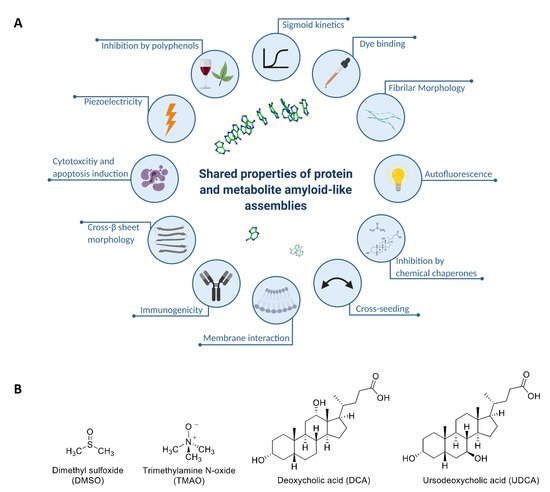
Figure 1. (A) Illustration of the shared properties of metabolite amyloid-like structures and protein or peptide amyloids: nanoscale fibrillar morphology [29][30][31][32], β-sheet secondary structure [33], amyloid-specific fluorescent dye binding [27][29][30], cytotoxicity and apoptosis induction [27][29][30][31][32][34][35], membrane interaction [36][37][38][39][40], inhibition by polyphenols [27][41], cross-seeding with protein amyloids [32][42], immunogenicity [27][29][32][43], intrinsic fluorescence [44][45], sigmoid growth curve [27], piezoelectricity [46], and inhibition by chemical chaperones (this study). Created with BioRender.com. (B) Molecular structure of DMSO, TMAO, DCA, and UDCA.
While the effect of molecular and chemical chaperones on misfolded proteins and the formation of proteinaceous amyloid assemblies is well characterized [6][47][48], it is unknown whether similar chaperoning mechanisms affect metabolite self-assembly. In this study, we applied a multimodal in vivo-in vitro-in silico approach to characterize the effect of chemical chaperones on metabolite self-assembly and amyloid formation. For this purpose, we examined the effect of four naturally occurring representative chemical chaperones: the DMSO osmolyte (found in various fruits, vegetables, grains, and beverages), TMAO osmolyte (one of the gut-derived metabolites found in diverse marine organisms), and the hydrophobic bile acids DCA and UDCA (Figure 1B) [11][49][50].
2. Chemical Chaperones Suppress Adenine Toxicity and Inhibit Intracellular Amyloid Formation in a Mutant Yeast Salvage Model
We first utilized our established salvage yeast model of adenine self-assembly (aah1Δapt1Δ) to examine the effect of DMSO, TMAO, DCA, and UDCA on phenotypes associated with adenine toxicity [27]. In line with their reported ability to inhibit protein and peptide amyloid formation [11], we observed a significant dose-dependent rescue of the adenine toxicity in yeast (Figure 2).
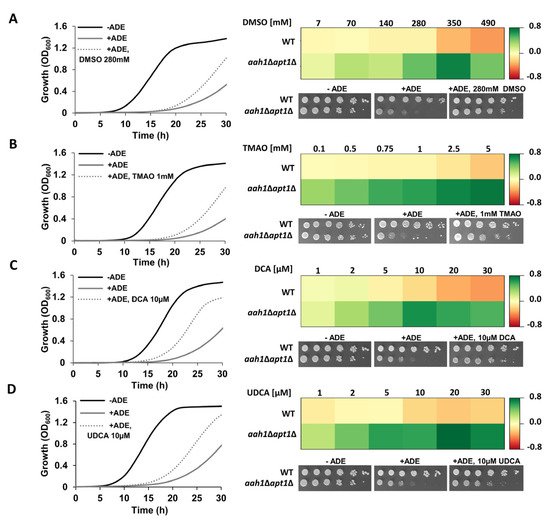
Figure 2. Inhibition of adenine toxicity by chemical chaperones in a yeast model. Left panel: Growth curves of aah1Δapt1Δ cells under the indicated conditions, at selected chaperones concentrations. Cells were grown at 30 °C and the absorbance at OD600 was measured over time (see Material and Methods). The results are representative of three independent biological repeats. Right panel: Dose-response heat maps representing the difference in optical density between a chaperone-supplemented culture and the control condition (+ADE) when the control cells reached OD600 = 0.6. Green, rescue. Red, toxicity. WT and aah1Δapt1Δ yeast were serially diluted and spotted on synthetic defined (SD) media lacking adenine (−ADE), SD media with adenine supplemented at 2 mg/L as a control for DMSO and TMAO, or with 2 mg/L adenine and 0.1% DMSO as a control for DCA and UDCA (+ADE), and SD with 2 mg/L adenine and the indicated concentration of the chemical chaperone. (A) DMSO; (B) TMAO; (C) DCA; (D) UDCA.
In order to examine whether the chaperone-induced growth improvement correlates with inhibition of amyloid aggregation, we stained the cells with the ProteoStat amyloid-specific dye, previously demonstrated to stain both protein and metabolite amyloids in vivo [27][51]. Flow cytometry and confocal microscopy were applied to visualize and analyze the intracellular amyloid content. As previously reported, adenine supplementation to the salvage mutant dramatically increased the intracellular amyloid content (Figure 3A,B), and resulted in a punctate staining pattern (Figure 3C,D) [27]. In accordance with the growth assays (Figure 2), the supplementation of chemical chaperones resulted in a significant reduction in the intracellular aggregate content in the mutant strain (Figure 3A,B). Furthermore, treatment with the chaperones significantly reduced the intracellular staining intensity (Figure 3C,D).
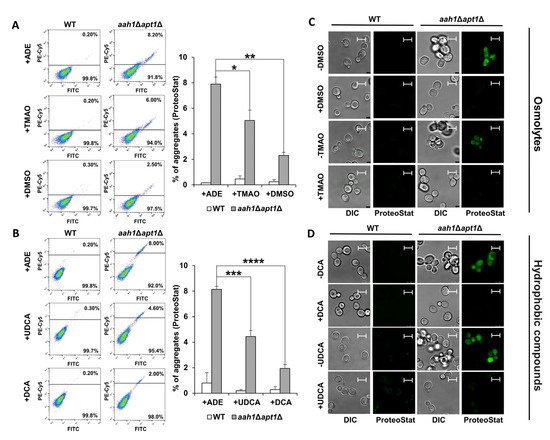
Figure 3. Chemical chaperones inhibit adenine aggregation in the salvage yeast model. (A,B) Flow cytometry analysis of wild type (WT) and aah1Δapt1Δ yeast cells under the indicated conditions following ProteoStat staining. Left: Flow cytometry data. Right: Quantification of ProteoStat positive cells under the indicated conditions. * p < 0.05; ** p < 0.005; *** p < 0.001; **** p < 0.0001 (Student’s t-test). Values are the average ± s.d. of three independent biological repeats. (A) 2 mg/L adenine (+ADE), 2 mg/L adenine and 1 mM TMAO (+TMAO) or 140 mM DMSO (+DMSO). (B) 2 mg/L adenine containing 0.05% DMSO (+ADE), 2 mg/L adenine containing 0.05% DMSO, 10 µM UDCA (+UDCA) or 10 µM DCA (+DCA). (C,D) Representative confocal microscopy images of WT and aah1Δapt1Δ cells under the same conditions as in (A,B). Scale bar is 5 μm.
3. Chemical Chaperones Alter the Biophysical Properties of Adenine Amyloid-like Assemblies
We next aimed to elucidate the mechanism of action underlying the effect of chemical chaperones on the cellular phenotypes investigated (Figure 2 and Figure 3). Therefore, we examined whether chemical chaperones directly impede adenine self-assembly in vitro by monitoring the binding kinetics of the fluorescent amyloid-specific dye Thioflavin-T (ThT). In line with the yeast assays data, all the tested chemical chaperones significantly reduced the adenine self-assembly rates in a dose-dependent manner, as demonstrated by inhibition of ThT binding (Figure 4A–D). Similar results were obtained using ProteoStat, providing the first demonstration of its utilization for staining metabolite amyloids in vitro. The reduction in the ProteoStat fluorescence in a dose-dependent manner, reinforces the notion that chemical chaperones possess the ability to inhibit adenine self-assembly.
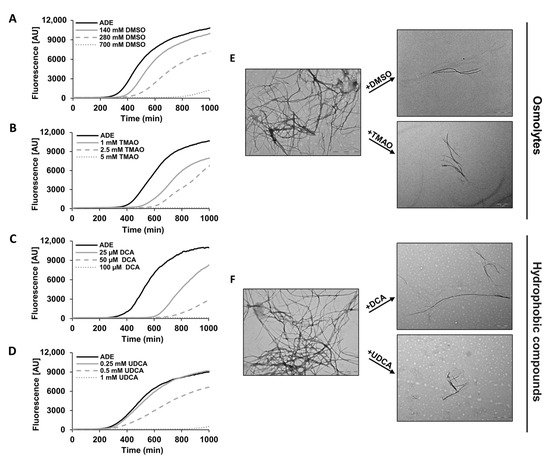
Figure 4. Inhibition of adenine fibril formation by different chemical chaperones in vitro. (A–D) Kinetic analysis of adenine aggregation as measured in a ThT binding assay. Adenine was dissolved at 6 mg/mL in PBS and heated to 90 °C to obtain a monomeric solution, followed by the addition of the chemical chaperones at the indicated concentrations (see Materials and Methods). ThT was added to a final concentration of 40 µM, and fluorescence was measured over time with excitation and emission wavelengths of 450 nm and 480 nm, respectively. The results are representative of three independent repeats. (E,F) Representative TEM micrographs of 1 mg/mL adenine in the presence or absence of the indicated chemical chaperone at 1.4 mM DMSO, 0.1 mM TMAO, 50 µM DCA, and 50 µM UDCA. Scale bar is 1 µm. The images are representatives of three double-blinded repeats.
References
- Vendruscolo, M.; Knowles, T.P.; Dobson, C.M. Protein Solubility and Protein Homeostasis: A Generic View of Protein Misfolding Disorders. Cold Spring Harb. Perspect. Biol. 2011, 3, a010454.
- Gazit, E. The ”Correctly Folded” state of proteins: Is it a metastable state? Angew. Chem. Int. Ed. 2002, 41, 257–259.
- Baldwin, A.J.; Knowles, T.; Tartaglia, G.G.; Fitzpatrick, A.W.; Devlin, G.L.; Shammas, S.L.; Waudby, C.; Mossuto, M.F.; Meehan, S.; Gras, S.; et al. Metastability of Native Proteins and the Phenomenon of Amyloid Formation. J. Am. Chem. Soc. 2011, 133, 14160–14163.
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68.
- Ke, P.C.; Zhou, R.; Serpell, L.C.; Riek, R.; Knowles, T.P.J.; Lashuel, H.A.; Gazit, E.; Hamley, I.W.; Davis, T.P.; Fändrich, M.; et al. Half a century of amyloids: Past, present and future. Chem. Soc. Rev. 2020, 49, 5473–5509.
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. In Vivo aspects of protein folding and quality control. Science 2016, 353, 4354.
- Ringe, D.; Petsko, G. What are pharmacological chaperones and why are they interesting? J. Biol. 2009, 8, 80.
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nat. Cell Biol. 2011, 475, 324–332.
- Labbadia, J.; Morimoto, R.I. The Biology of Proteostasis in Aging and Disease. Annu. Rev. Biochem. 2015, 84, 435–464.
- Bernier, V.; Lagacé, M.; Bichet, D.-G.; Bouvier, M. Pharmacological chaperones: Potential treatment for conformational diseases. Trends Endocrinol. Metab. 2004, 15, 222–228.
- Cortez, L.; Sim, V. The therapeutic potential of chemical chaperones in protein folding diseases. Prion 2014, 8, 197–202.
- Kumar, R. Role of naturally occurring osmolytes in protein folding and stability. Arch. Biochem. Biophys. 2009, 491, 1–6.
- Lee, C.S.; Hanna, A.; Wang, H.; Dagnino-Acosta, A.; Joshi, A.D.; Knoblauch, M.; Xia, Y.; Georgiou, D.K.; Xu, J.; Long, C.; et al. A chemical chaperone improves muscle function in mice with a RyR1 mutation. Nat. Commun. 2017, 8, 14659.
- Gong, B.; Zhang, L.Y.; Lam, D.S.C.; Pang, C.P.; Yam, G.H.F. Sodium 4-phenylbutyrate ameliorates the effects of cataract-causing mutant γD-crystallin in cultured cells. Mol. Vis. 2010, 16, 997–1003.
- Ozcan, U.; Yilmaz, E.; Özcan, L.; Furuhashi, M.; Vaillancourt, E.; Smith, R.O.; Görgün, C.Z.; Hotamisligil, G.S. Chemical Chaperones Reduce ER Stress and Restore Glucose Homeostasis in a Mouse Model of Type 2 Diabetes. Science 2006, 313, 1137–1140.
- Perlmutter, D.H. Chemical Chaperones: A Pharmacological Strategy for Disorders of Protein Folding and Trafficking. Pediatr. Res. 2002, 52, 832–836.
- Basseri, S.; Lhoták, Š.; Sharma, A.M.; Austin, R.C. The chemical chaperone 4-phenylbutyrate inhibits adipogenesis by modulating the unfolded protein response. J. Lipid Res. 2009, 50, 2486–2501.
- Getter, T.; Zaks, I.; Barhum, Y.; Ben-Zur, T.; Böselt, S.; Gregoire, S.; Viskind, O.; Shani, T.; Gottlieb, H.; Green, O.; et al. A Chemical Chaperone-Based Drug Candidate is Effective in a Mouse Model of Amyotrophic Lateral Sclerosis (ALS). ChemMedChem 2015, 10, 850–861.
- Pujols, J.; Díaz, S.P.; Pallares, I.; Ventura, S. Chemical Chaperones as Novel Drugs for Parkinson’s Disease. Trends Mol. Med. 2020, 26, 408–421.
- Bandyopadhyay, A.; Saxena, K.; Kasturia, N.; Dalal, V.; Bhatt, N.; Rajkumar, A.; Maity, S.; Sengupta, S.; Chakraborty, K. Chemical chaperones assist intracellular folding to buffer mutational variations. Nat. Chem. Biol. 2012, 8, 238–245.
- Sato, S.; Ward, C.L.; Krouse, M.E.; Wine, J.J.; Kopito, R.R. Glycerol Reverses the Misfolding Phenotype of the Most Common Cystic Fibrosis Mutation. J. Biol. Chem. 1996, 271, 635–638.
- Ip, P.; Sharda, P.R.; Cunningham, A.; Chakrabartty, S.; Pande, V.; Chakrabartty, A. Quercitrin and quercetin 3-β-d-glucoside as chemical chaperones for the A4V SOD1 ALS-causing mutant. Protein Eng. Des. Sel. 2017, 30, 431–440.
- Chang, K.-H.; Chiu, Y.-J.; Chen, S.-L.; Huang, C.-H.; Lin, C.-H.; Lin, T.-H.; Lee, C.-M.; Ramesh, C.; Wu, C.-H.; Huang, C.-C.; et al. The potential of synthetic indolylquinoline derivatives for Aβ aggregation reduction by chemical chaperone activity. Neuropharmacology 2016, 101, 309–319.
- Bennion, B.J.; Daggett, V. Counteraction of urea-induced protein denaturation by trimethylamine N-oxide: A chemical chaperone at atomic resolution. Proc. Natl. Acad. Sci. USA 2004, 101, 6433–6438.
- Perez-Miller, S.; Younus, H.; Vanam, R.; Chen, C.-H.; Mochly-Rosen, D.; Hurley, T.D. Alda-1 is an agonist and chemical chaperone for the common human aldehyde dehydrogenase 2 variant. Nat. Struct. Mol. Biol. 2010, 17, 159–164.
- Gazit, E. Metabolite amyloids: A new paradigm for inborn error of metabolism disorders. J. Inherit. Metab. Dis. 2016, 39, 483–488.
- Laor, D.; Sade, D.; Shaham-Niv, S.; Zaguri, D.; Gartner, M.; Basavalingappa, V.; Raveh, A.; Pichinuk, E.; Engel, H.; Iwasaki, K.; et al. Fibril formation and therapeutic targeting of amyloid-like structures in a yeast model of adenine accumulation. Nat. Commun. 2019, 10, 62.
- Rencus-Lazar, S.; DeRowe, Y.; Adsi, H.; Gazit, E.; Laor, D. Yeast Models for the Study of Amyloid-Associated Disorders and Development of Future Therapy. Front. Mol. Biosci. 2019, 6, 15.
- Adler-Abramovich, L.; Vaks, L.; Carny, O.; Trudler, D.; Magno, A.; Caflisch, A.; Frenkel, D.; Gazit, E. Phenylalanine assembly into toxic fibrils suggests amyloid etiology in phenylketonuria. Nat. Chem. Biol. 2012, 8, 701–706.
- Shaham-Niv, S.; Adler-Abramovich, L.; Schnaider, L.; Gazit, E. Extension of the generic amyloid hypothesis to nonproteinaceous metabolite assemblies. Sci. Adv. 2015, 1, e1500137.
- Shaham-Niv, S.; Rehak, P.; Vuković, L.; Adler-Abramovich, L.; Král, P.; Gazit, E. Formation of Apoptosis-Inducing Amyloid Fibrils by Tryptophan. Isr. J. Chem. 2017, 57, 729–737.
- Tavassoly, O.; Sade, D.; Bera, S.; Shaham-Niv, S.; Vocadlo, D.J.; Gazit, E. Quinolinic Acid Amyloid-like Fibrillar Assemblies Seed α-Synuclein Aggregation. J. Mol. Biol. 2018, 430, 3847–3862.
- Zaguri, D.; Shaham-Niv, S.; Chakraborty, P.; Arnon, Z.A.; Makam, P.; Bera, S.; Rencus-Lazar, S.; Stoddart, P.R.; Gazit, E.; Reynolds, N.P. Nanomechanical Properties and Phase Behavior of Phenylalanine Amyloid Ribbon Assemblies and Amorphous Self-Healing Hydrogels. ACS Appl. Mater. Interfaces 2020, 12, 21992–22001.
- Zaguri, D.; Shaham-Niv, S.; Naaman, E.; Mimouni, M.; Magen, D.; Pollack, S.; Kreiser, T.; Leibu, R.; Rencus-Lazar, S.; Adler-Abramovich, L.; et al. Induction of retinopathy by fibrillar oxalate assemblies. Commun. Chem. 2020, 3, 1–9.
- De Luigi, A.; Mariani, A.; De Paola, M.; Depaolini, A.R.; Colombo, L.; Russo, L.; Rondelli, V.; Brocca, P.; Adler-Abramovich, L.; Gazit, E.; et al. Doxycycline hinders phenylalanine fibril assemblies revealing a potential novel therapeutic approach in phenylketonuria. Sci. Rep. 2015, 5, 15902.
- Shaham-Niv, S.; Rehak, P.; Zaguri, D.; Kolusheva, S.; Král, P.; Gazit, E. Metabolite amyloid-like fibrils interact with model membranes. Chem. Commun. 2018, 54, 4561–4564.
- Griffith, E.; Perkins, R.; Telesford, D.-M.; Adams, E.M.; Cwiklik, L.; Allen, H.C.; Roeselová, M.; Vaida, V. Interaction of l-Phenylalanine with a Phospholipid Monolayer at the Water–Air Interface. J. Phys. Chem. B 2015, 119, 9038–9048.
- Cutró, A.; Hollmann, A.; Cejas, J.; Maturana, P.; Disalvo, E.; Frías, M. Phenylalanine interaction with lipid monolayers at different pHs. Colloids Surf. B Biointerfaces 2015, 135, 504–509.
- Rosa, A.S.; Cutro, A.C.; Frías, M.A.; Disalvo, E.A. Interaction of Phenylalanine with DPPC Model Membranes: More Than a Hydrophobic Interaction. J. Phys. Chem. B 2015, 119, 15844–15847.
- Sankaranarayanan, K. Fibrils of phenylalanine adsorbed to Langmuir-Blodgett films: Role of Lipids. Soft Mater. 2015, 13, 219–224.
- Shaham-Niv, S.; Rehak, P.; Zaguri, D.; Levin, A.; Adler-Abramovich, L.; Vuković, L.; Král, P.; Gazit, E. Differential inhibition of metabolite amyloid formation by generic fibrillation-modifying polyphenols. Commun. Chem. 2018, 1, 25.
- Sade, D.; Shaham-Niv, S.; Arnon, Z.A.; Tavassoly, O.; Gazit, E. Seeding of proteins into amyloid structures by metabolite assemblies may clarify certain unexplained epidemiological associations. Open Biol. 2018, 8, 8.
- Zaguri, D.; Kreiser, T.; Shaham-Niv, S.; Gazit, E. Antibodies towards Tyrosine Amyloid-Like Fibrils Allow Toxicity Modulation and Cellular Imaging of the Assemblies. Molecules 2018, 23, 1273.
- Chen, X.; Luo, W.; Ma, H.; Peng, Q.; Yuan, W.Z.; Zhang, Y. Prevalent intrinsic emission from nonaromatic amino acids and poly(amino acids). Sci. China Chem. 2018, 61, 351–359.
- Shaham-Niv, S.; Arnon, Z.A.; Sade, D.; Lichtenstein, A.; Shirshin, E.A.; Kolusheva, S.; Gazit, E. Intrinsic Fluorescence of Metabolite Amyloids Allows Label-Free Monitoring of Their Formation and Dynamics in Live Cells. Angew. Chem. Int. Ed. 2018, 57, 12444–12447.
- Guerin, S.; Stapleton, A.; Chovan, D.; Mouras, R.; Gleeson, M.; McKeown, C.; Noor, M.R.; Silien, C.; Rhen, F.M.F.; Kholkin, A.; et al. Control of piezoelectricity in amino acids by supramolecular packing. Nat. Mater. 2018, 17, 180–186.
- Leandro, P. Protein Misfolding in Conformational Disorders: Rescue of Folding Defects and Chemical Chaperoning. Mini-Rev. Med. Chem. 2008, 8, 901–911.
- Kim, Y.E.; Hipp, M.S.; Bracher, A.; Hayer-Hartl, M.; Ulrich Hartl, F. Molecular Chaperone Functions in Protein Folding and Proteostasis. Annu. Rev. Biochem. 2013, 82, 323–355.
- Ganguly, P.; Boserman, P.; Van Der Vegt, N.F.A.; Shea, J.-E. Trimethylamine N-oxide Counteracts Urea Denaturation by Inhibiting Protein–Urea Preferential Interaction. J. Am. Chem. Soc. 2018, 140, 483–492.
- Pearson, T.W.; Dawson, H.J.; Lackey, H.B. Naturally occurring levels of dimethyl sulfoxide in selected fruits, vegetables, grains, and beverages. J. Agric. Food Chem. 1981, 29, 1089–1091.
- Oshinbolu, S.; Shah, R.; Finka, G.; Molloy, M.; Uden, M.; Bracewell, D.G. Evaluation of fluorescent dyes to measure protein aggregation within mammalian cell culture supernatants. J. Chem. Technol. Biotechnol. 2017, 93, 909–917.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Revisions:
2 times
(View History)
Update Date:
15 Sep 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No