 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Nico Ruprecht | + 1673 word(s) | 1673 | 2021-09-07 05:43:15 | | | |
| 2 | Rita Xu | -74 word(s) | 1599 | 2021-09-09 05:07:13 | | |
Video Upload Options
Platinum compounds such as cisplatin (cisPt) embody the backbone of combination chemotherapy protocols against advanced lung cancer. However, their efficacy is primarily limited by inherent or acquired platinum resistance, the origin of which has not been fully elucidated yet, although of paramount interest.
1. Introduction
2. Determination of Intracellular cisPt in Wild Type (wt) and Resitant Cells Using SC-ICP-MS
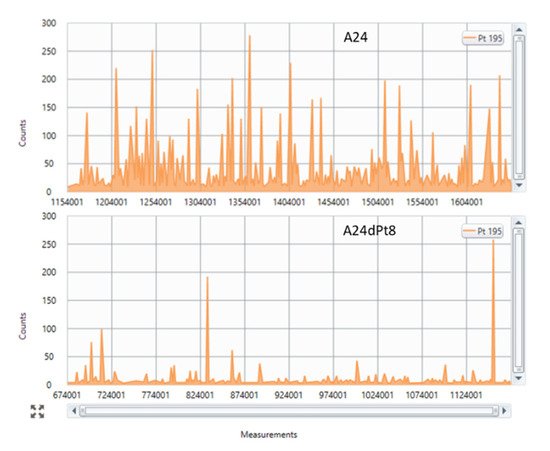
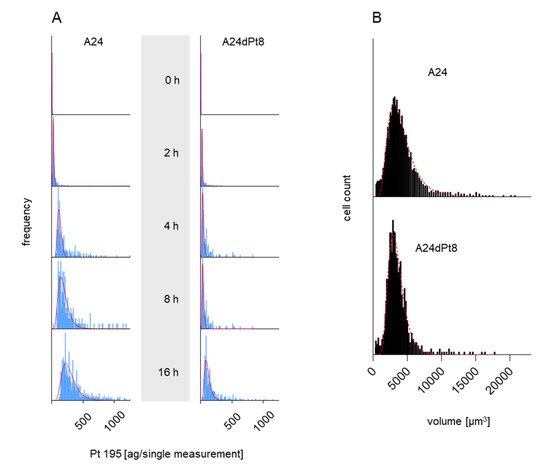
3. Intracellular cisPt Concentration in A24 and A24dPt8 Cells
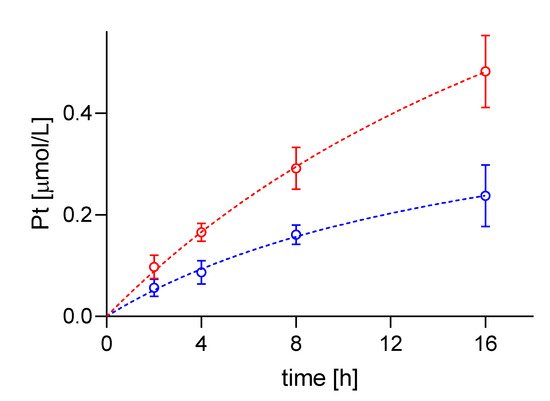
4. Difference in cisPt Uptake between wt and Resistant Cells
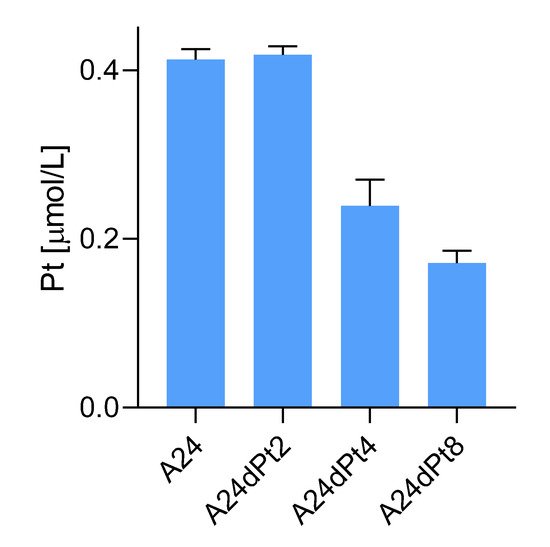
5. Difference in nuclear cisPt Uptake between wt and Resistant Cells
Average concentration of cisPt in nuclei of sensitive and resistant cells were determined, using measured nuclear volumes. The average cisPt concentration detected by SC-ICP-MS for 8 h in A24 cells was 0.6 ± 0.08 µM cisPt per nucleus, and 1.02 ± 0.27 µM cisPt per nucleus after 16 hours of incubation. In contrast, determination of nuclear cisPt concentration in resistant A24dPt8 cells revealed an average concentration of 0.23 ± 0.06 µM cisPt per nucleus at 8 h. At 16 h, an average of 0.59 ± 0.17 µM cisPt per nucleus was detected. CisPt concentrations measured in nuclei or cells within the same cell line are significantly higher in nuclei than in cells. From SC-ICP-MS measurements, an approximate 2-fold higher concentration of cisPt was found in nuclei compared to A24 cells at 8 h and 16 h, respectively (Fig. 7).
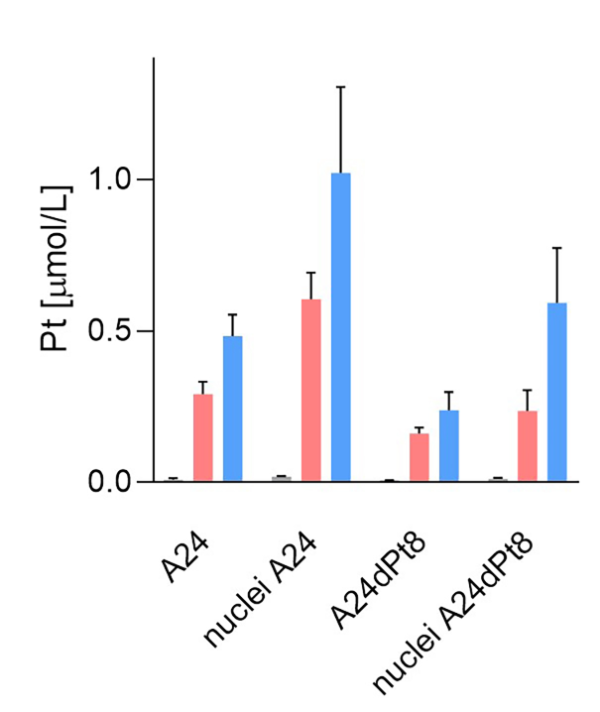
Figure 7. Nuclear and cellular cisPt concentration after incubation of A24 and A24DPt8 cells for ▐ 0h ▐ 8h ▐ 16 hours with 30 µM cisPt.
6. Conclusion
By SC ICP-MS quantification, it was shown that uptake of cisPt by A24 wt cells is higher than the one of A24dPt8 cisPt resistant cells. Nuclear cisPt concentration is higher than cellular cisPt is both WT and resistant cell lines.
References
- Simpson, P.V.; Desai, N.M.; Casari, I.; Massi, M.; Falasca, M. Metal-based antitumor compounds: Beyond cisplatin. Futur. Med. Chem. 2019, 11, 119–135.
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2011, 31, 1869–1883.
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584.
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279.
- Kelland, L.R. Preclinical perspectives on platinum resistance. Drugs 2000, 59, 1–8.
- Yoshida, M.; Khokhar, A.R.; Siddik, Z.H. Biochemical pharmacology of homologous alicyclic mixed amine platinum (II) complexes in sensitive and resistant tumor cell lines. Cancer Res. 1994, 54, 3468–3473.
- Kalayda, G.V.; Wagner, C.H.; Jaehde, U. Relevance of copper transporter 1 for cisplatin resistance in human ovarian car-cinoma cells. J. Inorg. Biochem. 2012, 116, 1–10.
- Shenberger, Y.; Shimshi, A.; Ruthstein, S. EPR spectroscopy shows that the blood carrier protein, human serum albumin, closely interacts with the N-terminal domain of the copper transporter, Ctr1. J. Phys. Chem. B 2015, 119, 4824–4830.
- Samimi, G.; Safaei, R.; Katano, K.; Holzer, A.K.; Rochdi, M.; Tomioka, M.; Goodman, M.; Howell, S.B. Increased expression of the copper efflux transporter ATP7A mediates resistance to cisplatin, carboplatin, and oxaliplatin in ovarian cancer cells. Clin. Cancer Res. 2004, 10, 4661–4669.
- Qasem, Z.; Pavlin, M.; Ritacco, I.; Gevorkyan-Airapetov, L.; Magistrato, A.; Ruthstein, S. The pivotal role of MBD4–ATP7B in the human Cu (I) excretion path as revealed by EPR experiments and all-atom simulations. Metallomics 2019, 11, 1288–1297.
- Wessels, L.F.; Voss, F.K.; Ullrich, F.; Xu, G.; Blomen, V.A.; Borst, P.; Gerhards, N.M.; Rottenberg, S.; Lutter, D.; Guyader, C.; et al. Subunit composition of VRAC channels determines substrate specificity and cellular resistance to P t-based anti-cancer drugs. EMBO J. 2015, 34, 2993–3008.
- Rabik, C.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat. Rev. 2007, 33, 9–23.
- Kartalou, M.; Essigmann, J.M. Mechanisms of resistance to cisplatin. Mutat. Res. 2001, 478, 23–43.
- Galluzzi, L.; Vitale, I.; Michels, J.; Brenner, C.; Szabadkai, G.; Harel-Bellan, A.; Castedo, M.; Kroemer, G. Systems biology of cisplatin resistance: Past, present and future. Cell Death Dis. 2014, 5, e1257.
- Fennell, D.; Summers, Y.; Cadranel, J.; Benepal, T.; Christoph, D.; Lal, R.; Das, M.; Maxwell, F.; Visseren-Grul, C.; Ferry, D. Cisplatin in the modern era: The backbone of first-line chemotherapy for non-small cell lung cancer. Cancer Treat. Rev. 2016, 44, 42–50.
- Voets, T.; Nilius, B.; Vennekens, R. VRAC s swallow platinum drugs. EMBO J. 2015, 34, 2985–2987.
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424.
- Pirker, R. Conquering lung cancer: Current status and prospects for the future. Pulmonology 2020, 26, 283–290.
- Pirker, R. Biomarkers for immune checkpoint inhibitors in advanced nonsmall cell lung cancer. Curr. Opin. Oncol. 2019, 31, 24–28.
- Lai, G.-M.; Ozols, R.F.; Smyth, J.F.; Young, R.C.; Hamilton, T.C. Enhanced DNA repair and resistance to cisplatin in human ovarian cancer. Biochem. Pharmacol. 1988, 37, 4597–4600.
- Fujii, R.; Mutoh, M.; Sumizawa, T.; Chen, Z.-S.; Yoshimura, A.; Akiyama, S.-I. Adenosine triphosphate-dependent transport of leukotriene C4 by membrane vesicles prepared from cisplatin-resistant human epidermoid carcinoma tumor cells. J. Natl. Cancer Inst. 1994, 86, 1781–1784.
- Twentyman, P.R.; Wright, K.A.; Mistry, P.; Kelland, L.R.; Murrer, B.A. Sensitivity to novel platinum compounds of panels of human lung cancer cell lines with acquired and inherent resistance to cisplatin. Cancer Res. 1992, 52, 5674–5680.
- Ruprecht, N.; Hofmann, L.; Hungerbühler, M.N.; Kempf, C.; Heverhagen, J.T.; von Tengg-Kobligk, H. Generation of stable cisPt resistant lung adenocarcinoma cells. Pharmaceuticals 2020, 13, 109.
- Sørensen, B.H.; Thorsteinsdottir, U.A.; Lambert, I.H. Acquired cisplatin resistance in human ovarian A2780 cancer cells correlates with shift in taurine homeostasis and ability to volume regulate. Am. J. Physiol. Cell Physiol. 2014, 307, C1071–C1080.
- Sørensen, B.H.; Dam, C.S.; Stürup, S.; Lambert, I.H. Dual role of LRRC8A-containing transporters on cisplatin resistance in human ovarian cancer cells. J. Inorg. Biochem. 2016, 160, 287–295.
- Sørensen, B.H.; Nielsen, D.; Thorsteinsdottir, U.A.; Hoffmann, E.K.; Lambert, I.H. Downregulation of LRRC8A protects human ovarian and alveolar carcinoma cells against Cisplatin-induced expression of p53, MDM2, p21Waf1/Cip1, and Caspase-9/-3 activation. Am. J. Physiol. Cell Physiol. 2016, 310, C857–C873.


