 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Derek T Lamport | + 1929 word(s) | 1929 | 2021-08-03 06:21:55 | | | |
| 2 | Lindsay Dong | Meta information modification | 1929 | 2021-08-27 03:11:29 | | |
Video Upload Options
Novel molecular pinball machines of the plasma membrane control cytosolic Ca2+ levels that regulate plant metabolism. The essential components involve: 1. an auxin-activated proton pump; 2. arabinogalactan glycoproteins (AGPs); 3. Ca2+ channels; 4. auxin-efflux “PIN” proteins. Typical pinball machines release pinballs that trigger various sound and visual effects. However, in plants, “proton pinballs” eject Ca2+ bound by paired glucuronic acid residues of numerous glycomodules in periplasmic AGP-Ca2+. Freed Ca2+ ions flow down the electrostatic gradient through open Ca2+ channels into the cytosol, thus activating numerous Ca2+-dependent activities.
1. Introduction
Sixty years ago [1], the discovery of hydroxyproline (Hyp) firmly bound to the cell wall was the “founder event” for a new field in plant biology. Proteins specific to the cell wall had not previously been considered as components of an otherwise polysaccharide structure, apart from occasional hints in the literature. Indeed, the Hill reaction defined the photolysis of water as the ancient source of atmospheric oxygen [2][3]. On the evolutionary timescale, an increased atmospheric oxygen level eventually led to its use as a terminal electron acceptor but also a direct source of the hydroxyproline hydroxyl. The biosynthesis of hydroxyproline, first shown in sycamore cell suspensions, involved the direct incorporation of 18O2 into the hydroxyproline hydroxyl group [4]. Hence, the first step in understanding the structural and dynamic roles of the Hyp-rich proteins unique to plants was taken. Quite remarkably, mammalian systems have recruited prolyl hydroxylase as an oxygen sensor for hypoxia inducible factor (HIF), which in turn plays a crucial role in foetal development [5].
The first analyses of TCA-soluble cytoplasmic protein–polysaccharide complexes purified by preparative isoelectric focusing [6] were subsequently identified as arabinogalactan proteins. Data from [6] show representative species across the plant kingdom included dicots, a gymnosperm and a bryophyte. These were the first AGP analyses notable for their high alanine and low tyrosine, with a high galactose and arabinose content.
Such proteins, later named arabinogalactan proteins (AGPs), located mainly between the plasma membrane and cell wall, were periplasmic, analogous to Peter Mitchell’s bacterial periplasm [7]. However, they were not precursors to wall-bound proteins, based on their composition and absence of turnover in 14C-proline pulse-chase experiments [8].
2. The Origin of Ion Gradients
3. Plasma Membrane Dynamics
4. Proton Pumps
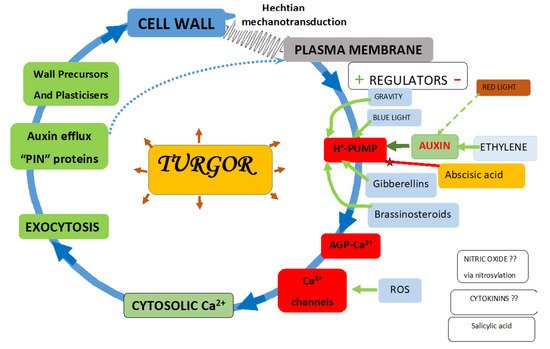
5. Indirect Evidence for the Role of AGPs in Ca2+ Homeostasis
An early indication of a connection between AGPs and Ca2+ appeared in 1991; the wound response of Acacia senegal and its secretory product, gum Arabic, consists of polysaccharides and glycoproteins related to AGPs [32]. Significantly, gum Arabic contains glucuronic acid (~10%) and binds approximately ~1% by weight of Ca2+. In the same year [33], the analysis of isolated plasma membranes revealed a bound AGP content of ~10% w/w. These AGPs were hydroxyproline-rich, with a significant glucuronic acid content (~10%). However, the crucial connection between Hyp-glycosubstituents and the pH-dependent Ca2+ binding property of AGPs with Ca2+ homeostasis only appeared quite recently when a molecular model depicted paired glucuronic acid residues of a Hyp-glycomodule that bound Ca2+ in a molecular dynamics simulation, which was then confirmed through an in vitro assay [34].
6. Direct Evidence for AGP Regulation of Ca2+ Homeostasis—A New Paradigm
6.1. AGP Glucuronic Acid Is Essential for Growth
6.2. AGP Glucuronic Acid Enables AGP-Ca2+ Binding
6.3. AGP-Ca2+ Binding Is a Major Source of Cytosolic Ca2+
6.4. Auxin Increases Cytosolic Ca2+
6.5. Ca2+ ATPase Recycles Cytosolic Ca2+
6.6. Ca2+ Waves Are Essential for Root Growth
6.7. AGPs Respond to Salt Stress
References
- Lamport, D.T.A.; Northcote, D.H. Hydroxyproline in primary cell walls of higher plants. Nature 1960, 188, 665–666.
- Hill, R. Oxygen Evolved by Isolated Chloroplasts. Nature 1937, 139, 881–882.
- Bendall, D.S. Robert Hill 2 April 1899–15 March 1991. Biogr. Mems. Fell. R. Soc. 1994, 40, 142–170.
- Lamport, D.T.A. Oxygen fixation into hydroxyproline of plant cell wall protein. J. Biol. Chem. 1963, 238, 1438–1440.
- Dunwoodie, S.L. The Role of Hypoxia in Development of the Mammalian Embryo. Dev. Cell 2009, 17, 755–773.
- Lamport, D.T.A. Cell wall metabolism. Ann. Rev. Plant Physiol. 1970, 21, 235–270.
- Mitchell, P. Approaches to the analysis of specific membrane transport. In Biological Structure and Function; 1st IUB/IUBS Intern. Symp., Stockholm, Sweden; Academic Press: London, UK, 1961; pp. 581–603.
- Pope, D.G. Relationships between hydroxyproline-containing proteins secreted into the cell wall and medium by suspension-cultured Acer pseudoplatanus cells. Plant Physiol. 1977, 59, 894–900.
- Mort, A.J.; Lamport, D.T.A. Anhydrous hydrogen fluoride deglycosylates glycoproteins. Analyt. Biochem. 1977, 82, 289–309.
- Gendler, S.J.; Burchell, J.M.; Duhig, T.; Lamport, D.T.A.; White, R.; Parker, M.; Taylor-Papadimitriou, J. Cloning the cDNA coding for differentiation and tumour- associated mucin glycoproteins expressed by human mammary epithelium. Proc. Natl. Acad. Sci. USA 1987, 84, 6060–6064.
- Chen, J.; Varner, J.E. Isolation and characterization of cDNA clones for carrot extensin and a proline-rich 33-kDa protein. Proc. Natl. Acad. Sci. USA 1985, 82, 4399–4403.
- Lamport, D.T.A. Hydroxyproline-O-glycosidic linkage of the plant cell wall glycoprotein extensin. Nature 1967, 216, 1322–1324.
- Tan, L.; Varnai, P.; Lamport, D.T.A.; Yuan, C.; Xu, J.; Qiu, F.; Kieliszewski, M.J. Plant O-Hydroxyproline Arabinogalactans Are Composed of Repeating Trigalactosyl Subunits with Short Bifurcated Side Chains. J. Biol. Chem. 2010, 285, 24575–24583.
- Lopez-Hernandez, F.; Tryfona, T.; Rizza, A.; Yu, X.L.; Harris, M.O.B.; Webb, A.A.R.; Kotake, T.; Dupree, P. Calcium Binding by Arabinogalactan Polysaccharides Is Important for Normal Plant Development. Plant Cell 2020, 32, 3346–3369.
- Zhang, Y.; Held, M.A.; Showalter, A.M. Elucidating the roles of three β-glucuronosyltransferases (GLCATs) acting on arabinogalactan-proteins using a CRISPR-Cas9 multiplexing approach in Arabidopsis. BMC Plant Biol. 2020, 20, 221.
- Ajayi, O.O.; Held, M.A.; Showalter, A.M. Three β-Glucuronosyltransferase Genes Involved in Arabinogalactan Biosynthesis Function in Arabidopsis Growth and Development. Plants 2021, 10, 1172.
- Liu, S.; Liu, C.C.; Froyd, K.D.; Schill, G.P.; Murphy, D.M.; Bui, T.P.; Dean-Day, J.M.; Weinzierl, B.; Dollner, M.; Diskin, G.S.; et al. Sea spray aerosol concentration modulated by sea surface temperature. Proc. Natl. Acad. Sci. USA 2021, 118, e2020583118.
- Dobson, C.M.; Ellison, G.B.; Tuck, A.F.; Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proc. Natl. Acad. Sci. USA 2000, 97, 11864.
- Stoeckenius, W. The Purple Membrane of Salt-loving Bacteria. Sci. Am. 1976, 234, 38–46.
- Haruta, M.; Sussman, M.R. The effect of a genetically reduced plasma membrane protonmotive force on vegetative growth of Arabidopsis. Plant Physiol. 2012, 158, 1158–1171.
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1048–1058.
- Mitchell, P. David Keilin’s Respiratory Chain Concept and Its Chemiosmotic Consequences. In Nobel Lectures in Chemistry 1971–1980; World Scientific: Singapore, 1993; pp. 295–330.
- Abrahams, J.P.; Leslie, A.G.W.; Lutter, R.; Walker, J.E. Structure at 2.8 A resolution of F1·ATPase from bovine heart mitochondria. Nature 1994, 370, 621–628.
- Falhof, J.; Pedersen, J.T.; Fuglsang, J.T.; Palmgren, M. Plasma Membrane H+-ATPase Regulation in the Center of Plant Physiology. Mol. Plant 2016, 9, 323–337.
- Pacheco-Villalobos, D.; Diaz-Moeeno, M.; van der Schuren, A.; Tamaki, T.; Kang, Y.H.; Gujas, B.; Novak, O.; Jaspert, N.; Li, Z.; Wolf, S.; et al. The Effects of High Steady State Auxin Levels on Root Cell Elongation in Brachypodium. Plant Cell 2016, 28, 1009–1024.
- Cieslak, M.; Runions, A.; Prusinkiewicz, P. Auxin-driven patterning with unidirectional fluxes. J. Exp. Bot. 2015, 66, 5083–5102.
- Leyser, O. Auxin Signaling. Plant Physiol. 2018, 176, 465–479.
- Habets, M.E.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377.
- Rubery, P.H.; Sheldrake, A.R. Carrier-mediated Auxin Transport. Planta 1974, 118, 101–121.
- Zazimalova, E.; Murphy, A.S.; Yang, H.; Hoyerova, K.; Hosek, P. Auxin Transporters—Why So Many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552.
- Vanneste, S.; Friml, J. Calcium: The missing link in auxin action. Plants 2013, 2, 650–675.
- Qi, W.; Fong, C.; Lamport, D.T.A. Gum arabic glycoprotein is a twisted hairy rope: A new model based on O-galactosylhydroxyproline as the polysaccharide attachment site. Plant Physiol. 1991, 96, 848–855.
- Komalavilas, P.; Zhu, J.; Nothnagel, E.A. Arabinogalactan-proteins from the suspension culture medium and plasma membrane of rose cells. J. Biol. Chem. 1991, 266, 15956–15965.
- Lamport, D.T.A.; Varnai, P. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytol. 2013, 197, 58–64.
- Behera, S.; Zhaolong, X.; Luoni, L.; Bonza, M.C.; Doccula, F.G.; De Michelis, M.I.; Morris, M.J.; Schwarzlander, M.; Costa, A. Cellular Ca2+ Signals Generate Defined pH Signatures in Plants. Plant Cell 2018, 30, 2704–2719.
- De Vriese, K.; Himschoot, E.; Dunser, K.; Nguyen, L.; Drozdzecki, A.; Costa, A.; Nowack, M.K.; Kleine-Vehn, J.; Audenaert, D.; Beeckman, T.; et al. Identification of Novel Inhibitors of Auxin-Induced Ca2+ Signaling via a Plant-Based Chemical Screen. Plant Physiol. 2019, 180, 480–496.
- Junge, W.; Nelson, N. ATP Synthase. Annu. Rev. Biochem. 2015, 84, 631–657.
- Rahmati Ishka, M.; Brown, E.; Rosenberg, A.; Romanowsky, S.; Davis, J.A.; Choi, W.G.; Harper, J.F. Arabidopsis Ca2+-ATPases 1, 2, and 7 in the endoplasmic reticulum contribute to growth and pollen fitness. Plant Physiol. 2021, 185, 1966–1985.
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446.
- Lamport, D.T.A.; Kieliszewksi, M.J.; Showalter, A.M. Salt-stress upregulates periplasmic arabinogalactan-proteins: Using salt-stress to analyse AGP function. New Phytol. 2006, 169, 479–492.
- Green, A.E.; Unsworth, R.K.F.; Chadwick, M.A.; Jones, P.J.S. Historical Analysis Exposes Catastrophic Seagrass Loss for the United Kingdom. Front. Plant Sci. 2021, 12, 629962.
- Pfeifer, L.; Shafee, T.; Johnson, K.L.; Bacic, A.; Classen, B. Arabinogalactan-proteins of Zostera marina L. contain unique glycan structures and provide insight into adaption processes to saline environments. Nat. Sci. Rep. 2020, 10, 8232.

