Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Abdul Razak Mariatulqabtiah | + 2714 word(s) | 2714 | 2021-07-14 11:46:51 | | | |
| 2 | Nora Tang | Meta information modification | 2714 | 2021-08-23 09:47:24 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Mariatulqabtiah, A.R. Surveillance of Avian Influenza Viruses. Encyclopedia. Available online: https://encyclopedia.pub/entry/13435 (accessed on 25 June 2026).
Mariatulqabtiah AR. Surveillance of Avian Influenza Viruses. Encyclopedia. Available at: https://encyclopedia.pub/entry/13435. Accessed June 25, 2026.
Mariatulqabtiah, Abdul Razak. "Surveillance of Avian Influenza Viruses" Encyclopedia, https://encyclopedia.pub/entry/13435 (accessed June 25, 2026).
Mariatulqabtiah, A.R. (2021, August 21). Surveillance of Avian Influenza Viruses. In Encyclopedia. https://encyclopedia.pub/entry/13435
Mariatulqabtiah, Abdul Razak. "Surveillance of Avian Influenza Viruses." Encyclopedia. Web. 21 August, 2021.
Copy Citation
The measures employed for curbing AI outbreaks and morbidities have played major roles in the journey towards global AI control. Although commendable progress has been made, more effective surveillance programmes, treatment regimens and prophylactic vaccines, which are capable of tackling the rapidly evolving AIV need to be developed. Effective control measures are the best option at preventing an AI panzootic which at present seems inevitable. To achieve this, AIV surveillance serves as an important tool for the understanding of the evolutionary patterns and epidemiology of AIVs.
avian influenza
surveillance
avian influenza vaccine
VLP vaccine
M2e
veterinary avian influenza vaccine
universal influenza vaccines
virus
1. AIV Surveillance in Wild Birds
AIV surveillance programmes in wild birds are essential because of their role as natural hosts for AIVs. The birds are suitable systems within which AIVs can evolve and circulate [1]. Apart from AIVs of the H17 and H18 subtypes found in bats, surveillance has shown that all other subtypes are perpetuated in wild birds, especially migrating aquatic birds [2][3]. However, the large population sizes and long-distance migrations of wild birds could be limiting factors to effective surveillance. As shown in Figure 1, certain criteria are considered in selecting a target sample population for surveillance, and these include: The population should have a history of AIV infection, the wild bird species should be one in which AIVs are prevalent, and the sample population should be one that is ecologically at risk of AI especially if environmental samples from their habitat are infected with AIVs [4].
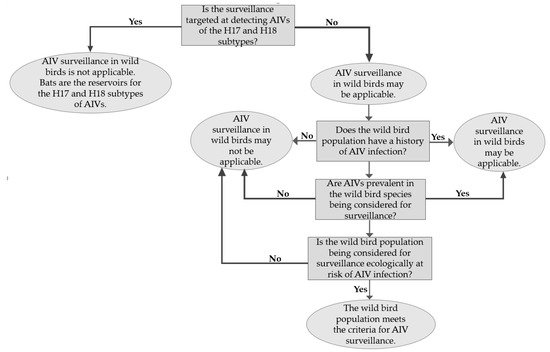
Figure 1. A flowchart representation of the criteria for selecting a wild bird population for avian influenza virus (AIV) surveillance.
In March 2013, a novel AIV H7N9 was detected in humans in six provinces and municipalities of China. Subsequently, genomic correlation, coalescent, and phylogenetic analyses were employed to ascertain the origin of the virus and the likely re-assortment events that gave rise to the novel AIV strain. It was established that the HA gene of H7N9 AIV originated from duck AIVs, and the NA gene was most likely from migratory birds of East Asian origin [5]. Since 2013, there had been five outbreaks of the zoonotic HPAI H7N9 in birds and humans from September 2013 to 2017 although it was asserted that the AIV transmission between species was unidirectional [6]. In March 2019, another outbreak occurred in domestic birds and humans in China, which suggested that the virus had effectively expanded its host range beyond wild birds, and was adapted to both poultry and humans. Genetic analysis of the H7N9 strain implicated revealed significant antigenic drift, which made it necessary to update the available H7N9 vaccine [7]. Interestingly, regular AIV surveillance of poultry-related environments had been embarked upon in China since 2008, whereby 50 to 70 environmental samples were analysed per selected region per year. However, H7N9 was not detected by the regular passive surveillance. The virus was only detected in poultry when the surveillance was reinforced following the detection of H7N9 in a human carrier [7]. While having a regular surveillance programme in place is commendable, it could be improved upon by employing a larger sample size, a more extensive surveillance area, and more frequent sample analyses per year.
Wild birds are known to maintain most of the various subtypes of AIVs, especially during periods when AI outbreaks in domestic birds are not recorded [8][9][10][11]. Consequently, they have been implicated in the re-emergence of some strains of AIVs after long periods of perceived eradication [12]. A typical case was reported after the 2005 outbreak of HPAI H5N1 in migratory birds in Lake Qinghai, China. In May 2006, another outbreak occurred in the same location, and six AIV strains were isolated from the infected birds and were subjected to whole-genome sequencing. Five out of the six strains were similar to H5N1 A/Cygnus olor/Croatia/1/05 that was implicated in the 2005 H5N1 outbreak in swans in Croatia. The sixth strain was related to the H5N1 A/duck/Novosibirsk/02/05 that was isolated from ducks in Novosibirsk, Russia during the 2005 epizootic [13]. Considering how geographically distant Croatia is from China, it is interesting that five strains of H5N1 genetically similar to a Croatian strain would be found in infected wild birds in China. This indicates how important the role of migratory wild birds is, in AIV transmission. Before 2005, HPAIV surveillance in wild birds was rare. Surveillance studies in wild birds increased rapidly after the outbreak of HPAI H5N1 in several wild bird populations from 2005 to 2006. Most of these more recent studies were targeted at early detections of HPAIVs in wild avian fauna before transmission to domestic birds could occur. Detection of HPAIVs in wild birds should also be taken into consideration in vaccine development and administration. This is because the presence of a HPAIV strain in a given wild bird population is an indication that transmission to domestic birds in that geographical location may occur. Hence, such HPAIV detection should be followed with prophylactic vaccination of the domestic birds that are at risk of being infected.
Until now, the prevalence of HPAIV in wild birds is low, and recorded cases are mostly clustered. This makes it difficult for surveillance programmes targeted at early detection of HPAIVs to effectively achieve their aim of clearly establishing the source of infection during every given HPAIV outbreak [14]. Some surveillance programmes also strive to provide insights on the epidemiology of LPAIVs in wild bird reservoirs. Bergervoet et al. [15] performed a study highlighting the circulation of LPAIVs in wild birds and poultry in the Netherlands from 2006 to 2016. They evaluated strain diversity, geographical distribution, and genetic similarities and differences between wild bird and poultry AIVs. It was established that there was a correlation between host species and the HA and NA subtypes of the infecting AIV. The fact that some LPAIVs detected in wild birds were not found in poultry suggested that the transmission between hosts could be selective, and some viral factors could limit the host range of some LPAIVs. However, the same subtypes of LPAIVs were shown to circulate in poultry, geese, and mallards [9][15]. Interestingly, it was shown that LPAIV infections of ducks were seasonal, and these seasons coincided with the peaks of LPAIV prevalence in wild ducks. It was also highlighted that circulating viruses went undetected for prolonged periods, and reassortment of new LPAIV strains with those in circulation in wild birds occurred frequently [15]. These reassortment events may be responsible for the evolution of virulence from LPAIV to HPAIV. This suggests that surveillance programmes focused on LPAIV are as necessary as their HPAIV-focused counterparts.
Surveillance of AIVs in wild birds is mostly limited to cases where deaths are observed. However, AIVs are mostly non-pathogenic or low pathogenic in wild birds, when compared to poultry birds. Widespread deaths of AIV infected wild birds are relatively uncommon. Prior to HPAI H5N1, the only AI enzootic in wild birds that led to widespread deaths was recorded in South Africa in 1961 following HPAI H5N3 infection of the common terns (Sterna hirundo) [1]. AIV surveillance programmes that focus on dead wild birds will not provide sufficient insights into the LPAIV genotypes harboured by wild birds [14]. Consequently, this will limit the preparedness in the event when transmissions to poultry and humans occur. Furthermore, certain instances of AIV surveillance in wild birds appear to be embarked upon during AI outbreaks that spark global concern. Such surveillance programmes are mostly short-lived and provide little insights into the evolutionary trends and transmission cycles of AIVs over time [12][14].
Furthermore, surveillance of AIV in wild birds appears to be more focused on the virus than the hosts. This explains why these avian reservoirs are poorly defined with regards to reservoir species, geographical distribution of these species and the strains of AIV that they predominantly harbour. For most species of wild birds, it is unclear if there is any relationship between viral maintenance and age, sex, migration behaviour, habitat, and feeding behaviour. It has been shown that the avian reservoir species, flock size and flock structure are essential for the maintenance and transmission of AIVs [16]. In-depth surveillance of AIV in wild birds that focuses on the reservoir as much as it does on the AIVs will lead to a clearer understanding of the control strategies to be employed [1]. We believe that the introduction of surveillance programmes that are focused on the reservoir hosts is essential in elucidating the role of these hosts in effective HPAIV transmission.
2. AIV Surveillance in Domestic Birds
The objectives of AI surveillance in domestic birds vary among programmes. Surveillance programmes in developed economies are mainly focused on how AI affects the trade of poultry products, while those in developing economies are aimed at the effects of AI outbreaks on the development of their poultry industry [17]. Surveillance programmes in domestic birds are also adapted to the areas under study and the available resources. Furthermore, surveillance programmes in poultry have the common limitation of mostly focusing on the AIVs implicated in the most recent epizootics [12]. Consequently, this decreases the preparedness for tackling outbreaks by less recently encountered AIVs. Globally, AI surveillances in domestic birds from 2005 to 2010 were predominantly focused on monitoring the spread of HPAI H5N1 that was reported in over 60 countries in Africa, Asia and Europe [17][18]. HPAI H5N1 in domestic birds remains a global health burden for the severe economic losses incurred by poultry farmers and the threat of zoonotic transmission to humans. Early detection constitutes an important measure in the prevention of devastating AI enzootics [11]. As of 2010, 23 European nations, members of the World Organisation for Animal Health (OIE), had active HPAI and LPAI surveillances in domestic birds. These programmes are ongoing and are either locally funded or funded by the European Union (EU). The administrations of prophylactic vaccines against HPAI during the said period were reported in the Netherlands and Germany while LPAI preventive vaccine administrations were reported in Italy, Portugal, and France [17]. It is likely that less developed economies, especially those without access to EU funding, will be unable to effectively carry out AIV surveillance in domestic birds. This will consequently lower their ability for early detection and control of AI enzootics.
The first outbreak of HPAI H5N1 in Africa was reported in Nigeria in 2006 [19]. Locally performed surveillance studies reported no HPAI H5N1 outbreak from 2007 to mid-2008. However, HPAIV of clade 2.2.1 re-emerged in July 2008 [20]. As reported by the OIE, AI surveillance programmes in poultry in African nations in the 2005 to 2010 period, were performed as four separate projects. The first surveillance programme focused on four nations, the second focused on six nations, the third focused on 15 nations, and the most encompassing surveillance programme was carried out in 47 African nations. The major drawback of these surveillance programmes was that they only lasted from one to three years [17]. More recently, the administration of prophylactic HPAI vaccines to poultry birds in African nations is more widely practised. Despite the administration of prophylactic vaccines, HPAI H5N1 has remained enzootic in poultry birds in Egypt [21]. Since 2011, there has been no report of AI outbreaks involving H5N1 of the subclade 2.2.1.1. However, in 2014, H5N1 A/chicken/Egypt/Fadllah-7/2014 was isolated from symptomatic poultry birds in Egypt, and genetic analysis of the virus revealed ~98% similarity with the H5N1 strain of subclade 2.2.1.1 [22]. More recently, AI surveillance in Egypt from 2016 to 2018 showed that the AIVs of H9N2, H5N8 and H5N1 were still in circulation in domestic chickens and ducks [21]. While this shows the importance of continuous AIV surveillance in poultry, it highlights the need for a universal vaccine as strain-specific AI outbreaks are sporadic, and it is impractical to produce and administer protective vaccines against every AIV strain.
From 2005 to 2009, seven Asian nations reported HPAI outbreaks in poultry. Of these countries, China and Thailand reported having locally funded ongoing surveillance programmes [17]. China, Indonesia, and Vietnam reported having preventive vaccination programmes against HPAI in poultry [17]. In Malaysia, HPAI outbreaks were reported in poultry in 2004, 2006, and 2007. After a decade of no recorded outbreak, a re-emergence was reported in 2017. Inter-district spread of HPAI H5N1 in Malaysia is believed to be due to the smuggling of poultry birds, trading of poultry and poultry products, and infection from wild avian fauna [23]. AI surveillance was carried out in domestic waterfowl from 2007 to 2012 in Bangladesh. It was reported that 4.4% of the tested birds were shown to harbour the AIVs of H1N1, H1N3, H3N2, H3N6, H3N8, H4N1, H4N2, H4N6, H5N1, H5N2, H6N1, H7N9, H9N2, H11N2, H11N3, and H11N6. Interestingly, 99% of the AIV-positive birds were asymptomatic carriers, suggesting that they were vital for maintaining AIVs in circulation, which presented an imminent threat of an outbreak [24]. The effectiveness of bird culling, vaccination, and surveillance as control measures to curb AI transmission in Bangladesh was analysed. The ideal control measure was one that employed all three measures [25]. Furthermore, from the first LPAI H9N2 outbreak in China in 1994 and the first HPAI H5N1 outbreak in 1996, several outbreaks of these AIVs were recorded and they were described as being enzootic in China [26][27]. Also, novel reassortants of AIVs of the H5N6, H7N9, and H10N8 subtypes had been reported to be circulating in poultry birds in China [7][27]. The live animal market practices in China are believed to be the cause of keeping the AIVs in circulation [27]. Hence, consistent surveillance and control programmes are essential as the nation has been described as being geographically conducive for the emergence of novel AIVs [18]. In China, vaccination programmes to curb the spread of AIVs in poultry have been implemented. However, the vaccines have not been reported to completely eradicate the enzootic AIVs [28]. Strict restrictions on the live animal market practices may curtail the AIV circulation in China.
3. Antiviral Treatment Regimens for Avian Influenza
The first line of control during an AI outbreak is the culling of the infected birds, which is then followed by an antiviral treatment regimen [25]. Current AI treatments include antiviral agents that disrupt viral assembly, attachment, or replication, while other AI regimens act via RNA interference [29][30]. Antiviral agents such as amantadine hydrochloride have been effectively used in HPAIV H5N9-infected turkeys and H5N2-infected chickens. They disrupt viral replication by blocking the AIV M2 ion channel protein [31][32]. Although amantadine is suitable for all bird species, AIVs easily develop resistance to it and the drug residues are found in the meat and eggs of treated birds [30][33]. Some other antiviral agents such as oseltamivir and zanamivir, which inhibit the AIV NA protein, have been effective for AI treatment in chickens [34]. These NA inhibitors are effective against all AIV subtypes [35]. However, a prolonged administration is required for effective treatment of infected birds [30].
Interestingly, the use of herbs in the treatment of AI had also been demonstrated. Essential oils of eucalyptus and peppermint administered to H9N2-infected broilers caused a reduction in microscopic lesions in their trachea, and a significant decrease in mortality [36][37]. Chicken models were also protected after the administration of green tea extracts to birds challenged with H1N1 virus A/NWS/33 [38]. Furthermore, a Chinese herbal medicine, NAS preparation, protected H9N2-infected chickens from exhibiting symptoms. However, the treated chickens were still capable of transmitting the virus to healthy chickens [39]. Some of the studies tested the efficacies of these herbs on AIV-infected cell cultures. Leaf extracts of black currant (Ribes nigrum folium) were shown to prevent the entry of H7N7 and H1N1 AIVs into Madin Darby canine kidney (MDCK) cells [40]. Also, Isatis indigotica root extracts were shown to prevent viral attachment of H1N1, H3N2, H6N2, and H9N2 AIVs to MDCK cells [41].
More recently, avian cytokines had been employed in the treatment of AI. They showed broad-spectrum efficacy against various AIV subtypes and this efficacy was not affected by the antigenic shift of AIVs [30]. Treated birds were also reported to recover rapidly. On the other hand, the cytokines were unstable and as such, were not developed for field applications [35]. Also, the high cost of production was a limiting factor. Short interfering RNAs (siRNA) had been highlighted as a prospective AI treatment [30]. The advantage of siRNAs as a treatment regimen was their specificity to the virus for which they were designed. However, this specificity implied a lack of broad-spectrum for a wide range of AIVs. Also, they were quickly degraded by the host’s immune defence mechanisms [30][35].
References
- Stallknecht, D.E.; Brown, J.D. Wild birds and the epidemiology of avian influenza. J. Wildl. Dis. 2007, 43, 15–20.
- Venkatesh, D.; Poen, M.J.; Bestebroer, T.M.; Scheuer, R.D.; Vuong, O.; Chkhaidze, M.; Machablishvili, A.; Mamuchadze, J.; Ninua, L.; Fedorova, N.B.; et al. Avian Influenza Viruses in Wild Birds: Virus Evolution in a Multihost Ecosystem. J. Virol. 2018, 92, 1–20.
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza a viruses. Microbiol. Rev. 1992, 56, 152–179.
- Gomaa, M.R.; Khalil, A.A.; Kandeil, A.; Sabir, J.S.M.; Kayed, A.; Moatasim, Y.; El saied, M.F.; El-safty, M.M.; Kayali, G.; Ali, M.A. Development of an effective contemporary trivalent avian influenza vaccine against circulating H5N1, H5N8, and H9N2 in Egypt. Poult. Sci. 2019, 98, 6289–6295.
- Liu, D.; Shi, W.; Shi, Y.; Wang, D.; Xiao, H.; Li, W.; Bi, Y.; Wu, Y.; Li, X.; Yan, J.; et al. Origin and diversity of novel avian influenza A H7N9 viruses causing human infection: Phylogenetic, structural, and coalescent analyses. Lancet 2013, 381, 1926–1932.
- Lin, Y.P.; Shu, L.L.; Wright, S.; Bean, W.J.; Sharp, G.B.; Shortridge, K.F.; Webster, R.G. Analysis of the influenza virus gene pool of avian species from Southern China. Virology 1994, 198, 557–566.
- Yu, D.; Xiang, G.; Zhu, W.; Lei, X.; Li, B.; Meng, Y.; Yang, L.; Jiao, H.; Li, X.; Huang, W.; et al. The re-emergence of highly pathogenic avian influenza H7N9 viruses in humans in Mainland China, 2019. Eurosurveillance 2019, 24, 1–9.
- Jang, Y.H.; Seong, B.L. The quest for a truly universal influenza vaccine. Front. Cell. Infect. Microbiol. 2019, 9, 1–24.
- Van Gils, J.A.; Munster, V.J.; Radersma, R.; Liefhebber, D.; Fouchier, R.A.M.; Klaassen, M. Hampered foraging and migratory performance in swans infected with low-pathogenic avian influenza A virus. PLoS ONE 2007, 2, 1–6.
- Osterhaus, A.; Fouchier, R.; Rimmelzwaan, G. Towards universal influenza vaccines? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2766–2773.
- De Vries, R.D.; Herfst, S.; Richard, M. Avian influenza: A virus pandemic preparedness and vaccine development. Vaccines 2018, 6, 46.
- Machalaba, C.C.; Elwood, S.E.; Forcella, S.; Smith, K.M.; Hamilton, K.; Jebara, K.B.; Swayne, D.E.; Webby, R.J.; Mumford, E.; Mazet, J.A.; et al. Global avian influenza surveillance in wild birds: A strategy to capture viral diversity. Emerg. Infect. Dis. 2015, 21.
- Wang, G.; Zhan, D.; Li, L.; Lei, F.; Liu, B.; Liu, D.; Xiao, H.; Feng, Y.; Li, J.; Yang, B.; et al. H5N1 avian influenza re-emergence of Lake Qinghai: Phylogenetic and antigenic analyses of the newly isolated viruses and roles of migratory birds in virus circulation. J. Gen. Virol. 2008, 89, 697–702.
- Hoye, B.J.; Munster, V.J.; Nishiura, H.; Klaassen, M. Surveillance of Wild Birds for Avian Influenza Virus. Emerg. Infect. Dis. 2010, 16, 1–18.
- Bergervoet, S.A.; Pritz-Verschuren, S.B.E.; Gonzales, J.L.; Bossers, A.; Poen, M.J.; Dutta, J.; Khan, Z.; Kriti, D.; van Bakel, H.; Bouwstra, R.; et al. Circulation of low pathogenic avian influenza (LPAI) viruses in wild birds and poultry in the Netherlands, 2006–2016. Sci. Rep. 2019, 9, 1–12.
- Fouchier, R.A.M.; Munster, V.J. Epidemiology of low pathogenic avian influenza viruses in wild birds. OIE Revue Scientifique Technique 2009, 28, 49–58.
- Pavade, G.; Weber-vintzel, L.; Hamilton, K.; Dehove, A. OFFLU Review of Avian Influenza Surveillance and Epidemiological Projects in some European, African, and Asian Countries. 2011. Available online: https://rr-africa.oie.int/en/news/offlu-review-of-avian-influenza-surveillance-and-epidemiological-projects/ (accessed on 11 May 2020).
- Guan, Y.; Smith, G.J.D. The emergence and diversification of panzootic H5N1 influenza viruses. Virus Res. 2013, 178, 35–43.
- Teru, V.C.; Manu, S.A.; Ahmed, G.I.; Junaidu, K.; Newman, S.; Nyager, J.; Iwar, V.N.; Mshelbwala, G.M.; Joannis, T.; Maina, J.A.; et al. Situation-based survey of avian influenza viruses in possible “ bridge ” species of wild and domestic birds in Nigeria. Influenza Res. Treatment 2012, 2012, 1–8.
- Joannis, T.M.; Meseko, A.C.; Oladokun, A.T.; Ularamu, H.; Hussaini, G.; Egbuji, A.N.; Solomon, P.; Nyam, D.C.; Gado, A.D.; Luka, P.; et al. Serologic and virologic surveillance of avian influenza in Nigeria, 2006–2007. Eurosurveillance 2008, 42, 1–5.
- Kandeil, A.; Hicks, J.T.; Young, S.G.; Taweel, A.N.; El Kayed, A.S.; Moatasim, Y.; Kutkat, O.; Bagato, O.; Mckenzie, P.P.; Cai, Z.; et al. Active surveillance and genetic evolution of avian influenza viruses in Egypt. Emerg. Microbes Infect. 2019, 8, 1370–1382.
- Rohaim, M.A.; El-Naggar, R.F.; Hamoud, M.M.; Nasr, S.A.; Ismael, E.; Laban, S.E.; Ahmed, H.A.; Munir, M. Re-emergence of a novel H5N1 avian influenza virus variant subclade 2.2.1.1 in Egypt during 2014. Transbound. Emerg. Dis. 2017, 64, 1306–1312.
- Wan Norulhuda, W.A.W.; Tariq, J. An overview of highly pathogenic avian influenza (H5N1) outbreak cases in Kelantan, West Malaysia in year 2017. MJVR 2018, 9, 102–108.
- Khan, S.U.; Gurley, E.S.; Gerloff, N.; Rahman, M.Z.; Simpson, N.; Rahman, M.; Haider, M.; Chowdhury, S.; Balish, A.; Zaman, R.U.; et al. Avian influenza surveillance in domestic waterfowl and environment of live bird markets in Bangladesh, 2007–2012. Sci. Rep. 2018, 8, 1–10.
- Hill, E.M.; House, T.; Dhingra, M.S.; Kalpravidh, W.; Morzaria, S.; Osmani, M.G.; Brum, E.; Yamage, M.; Kalam, M.A.; Prosser, D.J.; et al. The impact of surveillance and control on highly pathogenic avian influenza outbreaks in poultry in Dhaka division, Bangladesh. PLoS Comput. Biol. 2018, 14, 1–27.
- Peacock, T.P.; James, J.; Sealy, J.E.; Iqbal, M. A global perspective on H9N2 avian influenza virus. Viruses 2019, 11, 620.
- Su, S.; Bi, Y.; Wong, G.; Gray, G.C.; Gao, G.F.; Li, S. Epidemiology, Evolution, and Recent Outbreaks of Avian Influenza Virus in China. J. Virol. 2015, 89, 8671–8676.
- Fan, X.; Hu, Y.; Zhang, G.; Wang, M. Veterinary influenza vaccines against avian influenza in China. Future Virol. 2015, 10, 585–595.
- Skalickova, S.; Heger, Z.; Krejcova, L.; Pekarik, V.; Bastl, K.; Janda, J.; Kostolansky, F.; Vareckova, E.; Zitka, O.; Adam, V.; et al. Perspective of use of antiviral peptides against influenza virus. Viruses 2015, 7, 5428–5442.
- Abdelwhab, E.M.; Hafez, H.M. Insight into alternative approaches for control of avian influenza in poultry, with emphasis on highly pathogenic H5N1. Viruses 2012, 4, 3179–3208.
- Lang, G.; Narayan, O.; Rouse, B.T. Prevention of malignant avian iniluenza by 1-adamantanamine hydrochloride. Arch. Ges. Virusforsch. 1970, 32, 171–184.
- Beard, C.W.; Brugh, M.; Webster, R.G. Emergence of amantadine-resistant H5N2 avian influenza virus during a simulated layer flock treatment program. Avian Dis. 1987, 31, 533–537.
- Rollinson, E.A. Prospects for antiviral chemotherapy in veterinary medicine: 2. Avian, piscine, canine, porcine, bovine and equine virus diseases. Antivir. Chem. Chemo. 1992, 3, 311–326.
- Meijer, A.; van der Goot, J.A.; Koch, G.; van Boven, M.; Kimman, T.G. Oseltamivir reduces transmission, morbidity, and mortality of highly pathogenic avian influenza in chickens. Int. Cong. 2004, 1263, 495–498.
- Abdelwhab, E.M.; Hafez, H.M. Control of avian influenza in poultry with antivirals and molecular manipulation control of avian influenza in poultry with antivirals and molecular manipulation. In Epidemiology II Theory, Research and Practice Publisher; iConcept Press: Hong Kong, China, 2015; ISBN 978-1-922227-76-8.
- Barbour, E.K.; El-hakim, R.G.; Kaadi, M.S.; Shaib, H.A.; Gerges, D.D.; Nehme, P.A. Evaluation of the histopathology of the respiratory system in essential oil-treated broilers following a challenge with Mycoplasma gallisepticum and/or H9N2 influenza virus. Intern. J. Appl. Res. Vet. Med. 2006, 4, 293–300.
- Barbour, E.K.; El-hakim, R.G.; Kaadi, M.S.; Shaib, H.A.; Gerges, D.D.; Nehme, P.A. Evaluation of essential oils in the treatment of broilers co-infected with multiple respiratory etiologic agents. Intern. J. Appl. Res. Vet. Med. 2009, 9, 317–323.
- Lee, H.J.; Lee, Y.N.; Youn, H.N.; Lee, D.H.; Kwak, J.H.; Seong, B.L.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Immunology, health, and disease, anti-influenza virus activity of green tea by-products in vitro and efficacy against influenza virus infection in chickens. Poult. Sci. 2012, 91, 66–73.
- Shang, R.F.; Liang, J.P.; Na, Z.Y.; Yang, H.J.; Lu, Y.; Hua, L.Y.; Guo, W.Z.; Cui, Y.; Wang, L. In vivo inhibition of NAS preparation on H9N2 Subtype AIV. Virol. Sin. 2010, 25, 145–150.
- Ehrhardt, C.; Dudek, S.E.; Holzberg, M.; Urban, S.; Hrincius, E.R.; Haasbach, E.; Seyer, R.; Lapuse, J.; Planz, O.; Ludwig, S. A Plant Extract of Ribes nigrum folium possesses anti-influenza virus activity in vitro and in vivo by preventing virus entry to host cells. PLoS ONE 2013, 8.
- Yang, Z.; Wang, Y.; Zhong, S.; Zhao, S.; Zeng, X.; Mo, Z.; Qin, S.; Guan, W.; Li, C.; Zhong, N. In vitro inhibition of influenza virus infection by a crude extract from Isatis indigotica root resulting in the prevention of viral attachment. Mol. Med. Rep. 2012, 5, 793–799.
More
Information
Subjects:
Virology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
897
Revisions:
2 times
(View History)
Update Date:
23 Aug 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No