 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | David Bishop | + 2990 word(s) | 2990 | 2021-08-02 11:12:53 | | | |
| 2 | Camila Xu | -8 word(s) | 2982 | 2021-11-18 10:01:15 | | | | |
| 3 | Camila Xu | Meta information modification | 2982 | 2021-11-19 01:56:41 | | |
Video Upload Options
Caffeine (1,3,7-trimethylxanthine) is an alkaloid widely used to improve exercise performance.
1. Introduction
2. The Pulmonary System
A question arising from the above is whether caffeine ingestion can attenuate the imposed respiratory system limitations during high-intensity whole-body exercise. There is some research suggesting that caffeine influences ventilation ( E), end-tidal O2 partial pressure (PETO2), and SaO2 during progressive treadmill exercise to exhaustion in athletes with moderate-to-severe exercise-induced hypoxemia [20]. Specifically, caffeine increased PETO2 and SaO2 during the highest workloads (75 to 90% of O2max), which was associated with an increased E. Interestingly, caffeine increased E at 100% of O2max but this was not enough to counteract the reduction in SaO2 with the increase in workload [31]. It was suggested that a large portion of the increased E with caffeine at this exercise intensity may have reached dead space ventilation rather than alveolar ventilation.
The mechanisms by which caffeine increases E and prevents arterial O2 desaturation during high-intensity whole-body exercise are not fully known. Studies in resting animals [32] and humans [33] suggest a centrally-mediated effect of caffeine, where caffeine may directly act on the respiratory center and increase its sensitivity to carbon dioxide. Equivocal results, however, have been reported about the action of caffeine on peripheral chemoreceptors. While caffeine seems to increase the resting ventilatory response to progressive isocapnic hypoxia and hyperoxic hypercapnia in healthy men [34], a study with well-trained athletes failed to find such effects [31]. Whether such central and/or peripheral effects of caffeine are preserved during high-intensity whole-body exercise is difficult to experimentally assess. Caffeine also promotes relaxation of smooth muscles of the bronchi, inducing bronchodilatation and protecting from bronchoconstriction induced by a bronchoprovocation challenge with dry gas or exercise [35,36]. As near-maximal bronchodilatation probably already occurs during high-intensity whole-body exercise [37,38], it is not known whether this caffeine-induced bronchodilation effect will remain important in such conditions. Finally, caffeine augments the contractility and endurance of the inspiratory muscles, while concomitantly reducing the sense of effort associated with fatiguing inspiratory muscle contractions, probably by overcoming a fatigue-induced reduction in the outflow from the central respiratory motoneuron pool and/or altering respiratory motor neuron firing patterns [39].
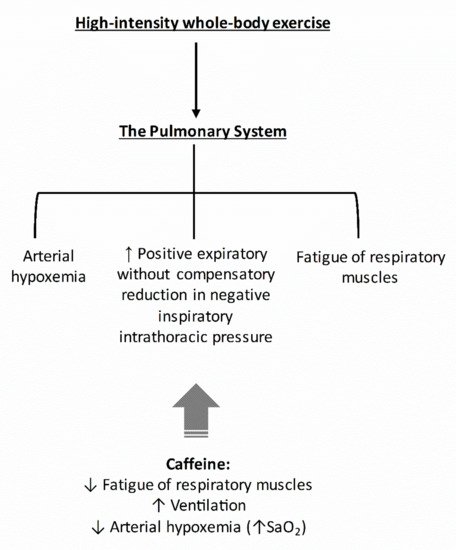
As the pulmonary system imposes considerable limitations during high-intensity whole-body exercise, the caffeine-induced increases in E and the preservation of near resting SaO2 may account for part of the ergogenic effects of caffeine in this mode of exercise. In Figure 3, we provided a hypothetical construct by which caffeine might affect the pulmonary system.
3. The Cardiovascular System
The cardiovascular system provides the link between pulmonary ventilation and oxygen usage at the cellular level. During high-intensity whole-body exercise, heart rate and stroke volume (and consequently cardiac output, Q ̇) increase progressively until reaching maximal values [40]. Leg blood flow (Q ̇L) will also be close to maximal and might limit both muscle O2 delivery and endurance exercise performance [28]. As written earlier, although controversial [41], Q ̇L during high-intensity whole-body exercise may be reflexively reduced via vasoconstriction of the vasculature of the exercising limb due to the increased O2 demand by the respiratory musculature [28]. In addition, a substantial augmentation of expiratory positive pressures during strenuous exercise may decrease stroke volume, further exacerbating the limitation of Q ̇L [42].
Data related to the effects of caffeine ingestion on stroke volume, Q ̇, and Q ̇L during high-intensity whole-body exercise are limited. To our knowledge, only one study has investigated this question and found no increase in stroke volume or Q ̇ during an incremental, symptom-limited, maximal supine bicycle exercise after caffeine ingestion [43]. There are more data regarding the effects of caffeine on maximal heart rate, but results are contradictory with some studies showing increased maximal heart rate [44][45] and others not [46][47][48]. However, most studies showing no effect of caffeine on maximal heart rate were conducted using multiple, 60-s, maximal cycling bouts [46][47], which may not have allowed sufficient time to reach maximal heart rate. Studies showing a positive effect of caffeine on maximal heart rate were performed using a maximal incremental test [44][45]. Indeed, using a constant-load, high-intensity whole-body exercise performed until volitional exhaustion, it was demonstrated that caffeine increased end-exercise heart rate by ~3% [16]. Whether this marginal effect on heart rate will be enough to affect Q ̇L is not known, but as stroke volume might plateau at about 40% of V ̇O2max with no further increase despite increasing intensity of exercise [49], a caffeine-induced increase in end-exercise heart rate during strenuous exercise might result in a small increase in Q ̇. Further research will be necessary to test this assumption.
As Q ̇ will be close to maximal during high-intensity whole-body exercise, several systems will be competing for the redistribution of this “limited” Q ̇. Many factors can influence the redistribution of blood flow during exercise, such as active muscle mass [50][51] and respiratory muscle work [30]. Furthermore, the well-known adenosine-receptor antagonism of caffeine may also impact blood flow redistribution. Indeed, adenosine causes vasodilation in several regional circulations and attenuates the release of renin [52]; therefore, blockage of adenosine receptors in blood vessels and the kidney via caffeine might alter blood flow redistribution. Although no data exist related to high-intensity whole-body exercise, it was demonstrated that caffeine acts as a powerful vasoconstrictor of inactive regions during dynamic leg exercise (cycle ergometry) at moderate intensity (65% V ̇O2max), increasing plasma angiotensin II (a powerful vasoconstrictor) and attenuating forearm blood flow (−53%) and forearm vascular conductance (−50%), which results in an increased mean arterial blood pressure [53]. Extrapolation of these results to near-maximal exercise should be done with caution, but some evidence suggests caffeine exacerbates the blood pressure response and plasma epinephrine concentration (another vasoconstrictor) during maximal exercise [43]. In Figure 4, we provided a hypothetical construct by which caffeine might affect the cardiovascular system. Further studies, however, will be necessary to investigate whether caffeine can increase Q ̇L during high-intensity whole-body exercise.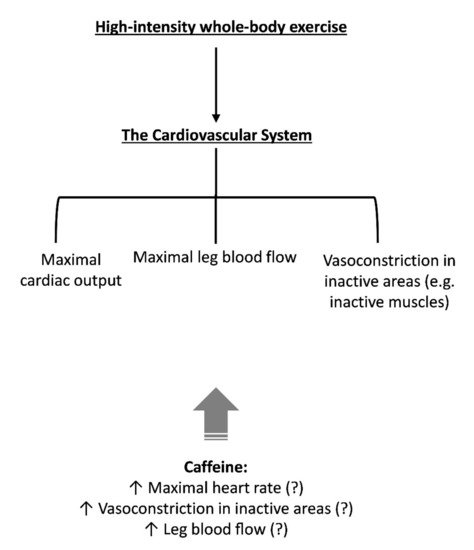
4. The Skeletal Muscle
Locomotor muscle oxygen uptake is a function of the product of Q ̇L and arterial-mixed venous oxygen difference (the Fick principle). The reduction in O2 delivery to active muscles by O2 desaturation and/or reduced blood flow may constrain high-intensity whole-body exercise performance by accelerating locomotor muscle fatigue [26]. During a high-intensity whole-body exercise carried out at identical work rates and for equal durations, preventing the normal O2 desaturation incurred during exercise reduces by about one-half the amount of exercise-induced quadriceps’ fatigue (as measured by the pre- to post-exercise reduction in evoked twitch force by supramaximal electrical or magnetic peripheral motor nerve stimulation) [26]. When SaO2 is “clamped” at 85% (moderate hypoxemia), locomotor muscle fatigue development over exercise time is exacerbated [54].
Recent findings suggest that caffeine increases active muscle oxygen saturation (measured via near-infrared spectrometer, NIRS) during submaximal workloads (30 to 60% of V ̇O2max) during a maximal incremental test with 1-min steps [48]. However, caffeine failed to increase muscle saturation at the highest workloads. It is noteworthy that only a main effect of substance was found. An absence of substance vs. workload interaction prevents a separate analysis for each workload without inflating type I error. A more suitable conclusion would be that caffeine increases muscle oxygen saturation during all workloads. In addition, it is important to consider that muscle oxygen saturation measured via NIRS-derived signals reflect the relationship between local muscle O2 delivery and muscle O2 utilization within the region of NIRS interrogation; thus, it is not possible to affirm from this data whether caffeine increased blood flow and consequently O2 delivery to the active muscles.
Despite the scarcity of studies investigating the effect of caffeine on Q ̇L, there is accumulating evidence using single-leg knee extensor exercise that caffeine attenuates locomotor muscle fatigue development [55,56]. While a direct relationship between increased O2 delivery and a reduction in locomotor muscle fatigue cannot be concluded from current data available in the literature, a direct effect of caffeine on skeletal muscles may also explain a reduced rate of peripheral fatigue development. It is, however, difficult to experimentally isolate a direct effect of caffeine on skeletal muscle in humans. In an attempt to isolate the muscular effects of caffeine, a study with electrical stimulation of the ulnar nerve (i.e., without CNS influence) showed an increase in muscle tension after caffeine ingestion before and after a protocol of muscle fatigue in healthy adults [57]. Caffeine also increased exercise tolerance in a group of spinal-cord-injured men during functional electrical stimulation of their paralyzed limbs to the point of fatigue [58]. Another study showed that caffeine potentiated the force of contraction during the final minute of a 2-min tetanic stimulation of the common peroneal nerve in healthy men [59]. However, a recent study failed to find an effect of caffeine ingestion on evoked forces in fresh quadricep muscles of healthy men [60]. Together, the results of these studies using electrical stimulation techniques argue in favor of a potential effect of caffeine on skeletal muscles, but its action seems to be more evident in studies in exercised/fatigued [58][59][61][62][63] than fresh muscles [60][64].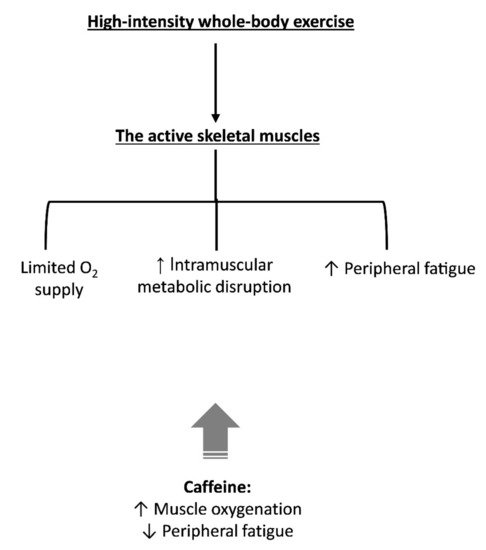
References
- Desbrow, B.; Leveritt, M. Awareness and use of caffeine by athletes competing at the 2005 Ironman Triathlon World Championships. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 545–558.
- Guest, N.S.; VanDusseldorp, T.A.; Nelson, M.T.; Grgic, J.; Schoenfeld, B.J.; Jenkins, N.D.M.; Arent, S.M.; Antonio, J.; Stout, J.R.; Trexler, E.T.; et al. International society of sports nutrition position stand: Caffeine and exercise performance. J. Int. Soc. Sports Nutr. 2021, 18, 1.
- Magkos, F.; Kavouras, S.A. Caffeine use in sports, pharmacokinetics in man, and cellular mechanisms of action. Crit. Rev. Food Sci. Nutr. 2005, 45, 535–562.
- Graham, T.E.; Spriet, L.L. Metabolic, catecholamine, and exercise performance responses to various doses of caffeine. J. Appl. Physiol. 1995, 78, 867–874.
- Ståhle, L.; Segersvärd, S.; Ungerstedt, U. Drug distribution studies with microdialysis II. Caffeine and theophylline in blood, brain and other tissues in rats. Life Sci. 1991, 49, 1843–1852.
- Grgic, J.; Grgic, I.; Pickering, C.; Schoenfeld, B.J.; Bishop, D.J.; Pedisic, Z. Wake up and smell the coffee: Caffeine supplementation and exercise performance—An umbrella review of 21 published meta-analyses. Br. J. Sports Med. 2020, 54, 681–688.
- Spineli, H.; Pinto, M.P.; Dos Santos, B.P.; Lima-Silva, A.E.; Bertuzzi, R.; Gitaí, D.L.G.; de Araujo, G.G. Caffeine improves various aspects of athletic performance in adolescents independent of their 163 C > A CYP1A2 genotypes. Scand. J. Med. Sci. Sports 2020, 30, 1869–1877.
- Chia, J.S.; Barrett, L.A.; Chow, J.Y.; Burns, S.F. Effects of Caffeine Supplementation on Performance in Ball Games. Sports Med. 2017, 47, 2453–2471.
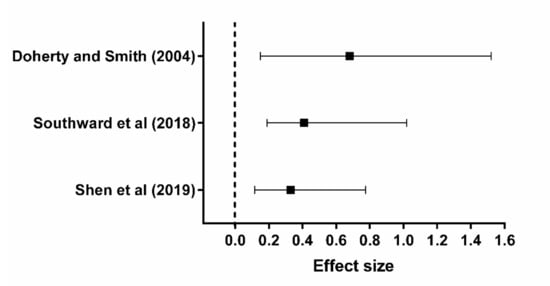
- Southward, K.; Rutherfurd-Markwick, K.J.; Ali, A. The Effect of Acute Caffeine Ingestion on Endurance Performance: A Systematic Review and Meta-Analysis. Sports Med. 2018, 48, 1913–1928.
- McLellan, T.M.; Caldwell, J.A.; Lieberman, H.R. A review of caffeine’s effects on cognitive, physical and occupational performance. Neurosci. Biobehav. Rev. 2016, 71, 294–312.
- Shen, J.G.; Brooks, M.B.; Cincotta, J.; Manjourides, J.D. Establishing a relationship between the effect of caffeine and duration of endurance athletic time trial events: A systematic review and meta-analysis. J. Sci. Med. Sport 2019, 22, 232–238.
- Doherty, M.; Smith, P.M. Effects of caffeine ingestion on exercise testing: A meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 626–646.
- Weavil, J.C.; Amann, M. Neuromuscular fatigue during whole body exercise. Curr. Opin. Physiol. 2019, 10, 128–136.
- Gastin, P.B. Energy system interaction and relative contribution during maximal exercise. Sports Med. 2001, 31, 725–741.
- Jones, A.M.; Vanhatalo, A.; Burnley, M.; Morton, R.H.; Poole, D.C. Critical power: Implications for determination of VO2max and exercise tolerance. Med. Sci. Sports Exerc. 2010, 42, 1876–1890.
- Silveira, R.; Andrade-Souza, V.A.; Arcoverde, L.; Tomazini, F.; Sansonio, A.; Bishop, D.J.; Bertuzzi, R.; Lima-Silva, A.E. Caffeine Increases Work Done above Critical Power, but Not Anaerobic Work. Med. Sci. Sports Exerc. 2018, 50, 131–140.
- Salvatore, C.A.; Jacobson, M.A.; Taylor, H.E.; Linden, J.; Johnson, R.G. Molecular cloning and characterization of the human A3 adenosine receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10365–10369.
- Lynge, J.; Hellsten, Y. Distribution of adenosine A1, A2A and A2B receptors in human skeletal muscle. Acta Physiol. Scand. 2000, 169, 283–290.
- Morales, A.P.; Sampaio-Jorge, F.; Barth, T.; Pierucci, A.P.T.R.; Ribeiro, B.G. Caffeine Supplementation for 4 Days Does Not Induce Tolerance to the Ergogenic Effects Promoted by Acute Intake on Physiological, Metabolic, and Performance Parameters of Cyclists: A Randomized, Double-Blind, Crossover, Placebo-Controlled Study. Nutrients 2020, 12, 2101.
- Chapman, R.F.; Mickleborough, T.D. The effects of caffeine on ventilation and pulmonary function during exercise: An often-overlooked response. Phys. Sportsmed. 2009, 37, 97–103.
- Dempsey, J.A.; Amann, M.; Romer, L.M.; Miller, J.D. Respiratory system determinants of peripheral fatigue and endurance performance. Med. Sci. Sports Exerc. 2008, 40, 457–461.
- Amann, M. Pulmonary system limitations to endurance exercise performance in humans. Exp. Physiol. 2012, 97, 311–318.
- Dempsey, J.A.; Wagner, P.D. Exercise-induced arterial hypoxemia. J. Appl. Physiol. 1999, 87, 1997–2006.
- Dempsey, J.A.; Hanson, P.G.; Henderson, K.S. Exercise-induced arterial hypoxaemia in healthy human subjects at sea level. J. Physiol. 1984, 355, 161–175.
- Harms, C.A.; McClaran, S.R.; Nickele, G.A.; Pegelow, D.F.; Nelson, W.B.; Dempsey, J.A. Exercise-induced arterial hypoxaemia in healthy young women. J. Physiol. 1998, 507, 619–628.
- Romer, L.M.; Dempsey, J.A. Effects of exercise-induced arterial hypoxaemia on limb muscle fatigue and performance. Clin. Exp. Pharmacol. Physiol. 2006, 33, 391–394.
- Dominelli, P.B.; Sheel, A.W. Exercise-induced arterial hypoxemia; some answers, more questions. Appl. Physiol. Nutr. Metab. 2019, 44, 571–579.
- Harms, C.A.; Babcock, M.A.; McClaran, S.R.; Pegelow, D.F.; Nickele, G.A.; Nelson, W.B.; Dempsey, J.A. Respiratory muscle work compromises leg blood flow during maximal exercise. J. Appl. Physiol. 1997, 82, 1573–1583.
- Harms, C.A.; Stager, J.M. Low chemoresponsiveness and inadequate hyperventilation contribute to exercise-induced hypoxemia. J. Appl. Physiol. 1995, 79, 575–580.
- Harms, C.A.; Wetter, T.J.; McClaran, S.R.; Pegelow, D.F.; Nickele, G.A.; Nelson, W.B.; Hanson, P.; Dempsey, J.A. Effects of respiratory muscle work on cardiac output and its distribution during maximal exercise. J. Appl. Physiol. 1998, 85, 609–618.
- Chapman, R.F.; Stager, J.M. Caffeine stimulates ventilation in athletes with exercise-induced hypoxemia. Med. Sci. Sports Exerc. 2008, 40, 1080–1086.
- Trippenbach, T.; Zinman, R.; Milic-Emili, J. Caffeine effect on breathing pattern and vagal reflexes in newborn rabbits. Respir. Physiol. 1980, 40, 211–225.
- Richmond, G.H. Action of Caffeine and Aminophylline as Respiratory Stimulants in Man. J. Appl. Physiol. 1949, 2, 16–23.
- D’Urzo, A.D.; Jhirad, R.; Jenne, H.; Avendano, M.A.; Rubinstein, I.; D’Costa, M.; Goldstein, R.S. Effect of caffeine on ventilatory responses to hypercapnia, hypoxia, and exercise in humans. J. Appl. Physiol. 1990, 68, 322–328.
- Duffy, P.; Phillips, Y.Y. Caffeine consumption decreases the response to bronchoprovocation challenge with dry gas hyperventilation. Chest 1991, 99, 1374–1377.
- Kivity, S.; Aharon, Y.B.; Man, A.; Topilsky, M. The Effect of Caffeine on Exercise-Induced Bronchoconstriction. Chest 1990, 97, 1083–1085.
- Merlini, M.; Beato, M.; Marcora, S.; Dickinson, J. The Effect of 1600 μg Inhaled Salbutamol Administration on 30 m SprInt. Performance Pre and Post a Yo-Yo Intermittent Running Test in Football Players. J. Sports Sci. Med. 2019, 18, 716–721.
- Molphy, J.; Dickinson, J.W.; Chester, N.J.; Loosemore, M.; Whyte, G. The Effect of 400 µg Inhaled Salbutamol on 3 km Time Trial Performance in a Low Humidity Environment. J. Sports Sci. Med. 2017, 16, 581–588.
- Supinski, G.S.; Levin, S.; Kelsen, S.G. Caffeine effect on respiratory muscle endurance and sense of effort during loaded breathing. J. Appl. Physiol. 1986, 60, 2040–2047.
- Miyamara, M.; Honda, Y. Oxygen intake and cardiac output during maximal treadmill and bicycle exercise. J. Appl. Physiol. 1972, 32, 185–188.
- Vogiatzis, I.; Louvaris, Z.; Wagner, P.D. Respiratory and locomotor muscle blood flow during exercise in health and chronic obstructive pulmonary disease. Exp. Physiol. 2020, 105, 1990–1996.
- Stark-Leyva, K.N.; Beck, K.C.; Johnson, B.D. Influence of expiratory loading and hyperinflation on cardiac output during exercise. J. Appl. Physiol. 2004, 96, 1920–1927.
- Sung, B.H.; Lovallo, W.R.; Pincomb, G.A.; Wilson, M.F. Effects of caffeine on blood pressure response during exercise in normotensive healthy young men. Am. J. Cardiol. 1990, 65, 909–913.
- Bunsawat, K.; White, D.W.; Kappus, R.M.; Baynard, T. Caffeine delays autonomic recovery following acute exercise. Eur. J. Prev. Cardiol. 2015, 22, 1473–1479.
- Olcina, G.J.; Muñoz, D.; Timón, R.; Caballero, M.J.; Maynar, J.I.; Córdova, A.; Maynar, M. Effect of caffeine on oxidative stress during maximum incremental exercise. J. Sports Sci. Med. 2006, 5, 621–628.
- Crowe, M.J.; Leicht, A.S.; Spinks, W.L. Physiological and cognitive responses to caffeine during repeated, high-intensity exercise. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 528–544.
- Vanakoski, J.; Kosunen, V.; Meririnne, E.; Seppälä, T. Creatine and caffeine in anaerobic and aerobic exercise: Effects on physical performance and pharmacokinetic considerations. Int. J. Clin. Pharmacol. Ther. 1998, 36, 258–262.
- Ruíz-Moreno, C.; Lara, B.; Brito de Souza, D.; Gutiérrez-Hellín, J.; Romero-Moraleda, B.; Cuéllar-Rayo, Á.; Del Coso, J. Acute caffeine intake increases muscle oxygen saturation during a maximal incremental exercise test. Br. J. Clin. Pharmacol. 2020, 86, 861–867.
- Zhou, B.; Conlee, R.K.; Jensen, R.; Fellingham, G.W.; George, J.D.; Fisher, A.G. Stroke volume does not plateau during graded exercise in elite male distance runners. Med. Sci. Sports Exerc. 2001, 33, 1849–1854.
- Secher, N.H.; Volianitis, S. Are the arms and legs in competition for cardiac output? Med. Sci. Sports Exerc. 2006, 38, 1797–1803.
- Klausen, K.; Secher, N.H.; Clausen, J.P.; Hartling, O.; Trap-Jensen, J. Central and regional circulatory adaptations to one-leg training. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1982, 52, 976–983.
- Arend, L.J.; Haramati, A.; Thompson, C.I.; Spielman, W.S. Adenosine-induced decrease in renin release: Dissociation from hemodynamic effects. Am. J. Physiol. 1984, 247, F447–F452.
- Daniels, J.W.; Molé, P.A.; Shaffrath, J.D.; Stebbins, C.L. Effects of caffeine on blood pressure, heart rate, and forearm blood flow during dynamic leg exercise. J. Appl. Physiol. 1998, 85, 154–159.
- Mira, J.; Floreani, M.; Savoldelli, A.; Amery, K.; Koral, J.; Oranchuk, D.J.; Messonnier, L.; Rupp, T.; Millet, G.Y. Neuromuscular Fatigue of Cycling Exercise in Hypoxia. Med. Sci. Sports Exerc. 2020.
- Meyers, B.; Cafarelli, E. Caffeine increases time to fatigue by maintaining force and not by altering firing rates during submaximal isometric contractions. J. Appl. Physiol. 2005, 99, 1056–1063.
- Pethick, J.; Winter, S.L.; Burnley, M. Caffeine ingestion attenuates fatigue-induced loss of muscle torque complexity. Med. Sci. Sports Exerc. 2018, 50, 236–245.
- Lopes, J.M.; Aubier, M.; Jardim, J.; Aranda, J.V.; Macklem, P.T. Effect of caffeine on skeletal muscle function before and after fatigue. J. Appl. Physiol. 1983, 54, 1303–1305.
- Mohr, T.; Van Soeren, M.; Graham, T.E.; Kjaer, M. Caffeine ingestion and metabolic responses of tetraplegic humans during electrical cycling. J. Appl. Physiol. 1998, 85, 979–985.
- Tarnopolsky, M.; Cupido, C. Caffeine potentiates low frequency skeletal muscle force in habitual and nonhabitual caffeine consumers. J. Appl. Physiol. 2000, 89, 1719–1724.
- Neyroud, D.; Cheng, A.J.; Donnelly, C.; Bourdillon, N.; Gassner, A.-L.; Geiser, L.; Rudaz, S.; Kayser, B.; Westerblad, H.; Place, N. Toxic doses of caffeine are needed to increase skeletal muscle contractility. Am. J. Physiol. Cell Physiol. 2019, 316, C246–C251.
- Tallis, J.; Duncan, M.J.; James, R.S. What can isolated skeletal muscle experiments tell us about the effects of caffeine on exercise performance? Br. J. Pharmacol. 2015, 172, 3703–3713.
- Tallis, J.; James, R.S.; Cox, V.M.; Duncan, M.J. The effect of physiological concentrations of caffeine on the power output of maximally and submaximally stimulated mouse EDL (fast) and soleus (slow) muscle. J. Appl. Physiol. 2012, 112, 64–71.
- Tallis, J.; Higgins, M.F.; Cox, V.M.; Duncan, M.J.; James, R.S. An exercise-induced improvement in isolated skeletal muscle contractility does not affect the performance-enhancing benefit of 70 µmol l–1 caffeine treatment. J. Exp. Biol. 2018, 221.
- Felippe, L.C.; Ferreira, G.A.; Learsi, S.K.; Boari, D.; Bertuzzi, R.; Lima-Silva, A.E. Caffeine increases both total work performed above critical power and peripheral fatigue during a 4-km cycling time trial. J. Appl. Physiol. 2018, 124, 1491–1501.
- Simmonds, M.J.; Minahan, C.L.; Sabapathy, S. Caffeine improves supramaximal cycling but not the rate of anaerobic energy release. Eur. J. Appl. Physiol. 2010, 109, 287–295.
- Mohr, M.; Nielsen, J.J.; Bangsbo, J. Caffeine intake improves intense intermittent exercise performance and reduces muscle interstitial potassium accumulation. J. Appl. Physiol. 2011, 111, 1372–1379.
- Chesley, A.; Howlett, R.A.; Heigenhauser, G.J.; Hultman, E.; Spriet, L.L. Regulation of muscle glycogenolytic flux during intense aerobic exercise after caffeine ingestion. Am. J. Physiol. 1998, 275, R596–R603.

