 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Garry Kerch | + 2732 word(s) | 2732 | 2020-06-30 10:48:01 | | | |
| 2 | Catherine Yang | Meta information modification | 2732 | 2020-08-10 05:01:15 | | |
Video Upload Options
The mechanical properties of living biological tissues change with aging and commonly can be associated with age-related diseases. Increase of tissue stiffness can be related also with thermodynamically favorable release of tightly bound to biological macromolecules water molecules.
1. Tissue Stiffness
Discher and co-authors reported in 2005 that cells respond to the rigidity of their planar substrate [1]. It has been further demonstrated that cellular processes in the three-dimensional matrix respond differently to microenvironment stiffness and, for example, in hyaluronic acid hydrogels, the differentiation behavior of human mesenchymal stem cells (hMSCs) depends on cellular traction. High degrees of cell spreading on stiffer surfaces favored osteogenesis, and lower degrees of cell spreading on softer surfaces favored adipogenesis [2]. Protein-conformational diseases and age-related diseases are associated with increasing tissue stiffness with increasing age [3][4][5]. Arterial stiffness was associated, not only with cardiovascular diseases, but also with neurodegenerative diseases, cognitive impairment, and vascular dementia [6][7][8]. Thus, the prevention of tissue stiffening is a therapeutic approach with the clinical potential to attenuate disease, which has been discussed in a recent review by Lampi and Reinhart-King [4]. It has also been suggested that antihypertensive drugs can decrease vascular stiffness and reduce the occurrence of cognitive impairment [8]. Increased extracellular matrix (ECM) stiffness was related to cancer, cardiovascular disease, diabetes, and other pathologies. ECM provides biomechanical cues that direct cell growth, migration, differentiation, and survival [9].
2. Tightly Bound Water
It is important to take into account that mechanical properties depend on the content and state of water bound to biological macromolecules [10][11]. The binding energy of water molecules to biological macromolecules decreases with aging, and tightly bound water transforms into loosely bound and free water. Tightly bound water is a plasticizer and the decrease in its content results in an increase in tissue stiffness. The decrease in bound water fraction in the extracellular matrix with advanced age was correlated with the loss of mechanical properties of the cortical bone in female and male BALB/c mice [12]. Strength and toughness of hydrated human bone decreases with decrease of bound water content, indicating the risk of fracture [13]. In bone, the quantity of water bound to collagen decreases with age [14]. Total body water (TBW) and intracellular water (ICW) decrease with age, but elderly individuals with higher ICW content have higher muscle strength and better functional performance [15]. An increase in the extracellular water (ECW) to ICW ratio in the upper legs determined using segmental bioelectrical impedance spectroscopy was associated with a decrease in muscle strength in older individuals [16]. Kim and co-authors demonstrated that high ECW/ICW ratio is a major risk indicator for all-cause mortality and cardiovascular diseases. A correlation between ECW/ICW with inflammation and pulse wave velocity was reported [17]. ECW/ICW also has been reported as a prognostic factor for survival in metastatic cancer [18]. A decrease in the ICW content with age proceeds with a higher rate than in the ECW content. Increase in ECW/ICW ratio accelerates after the age of 70 years [19].
Thus, it is very important to prevent or retard the transformation of intracellular tightly bound water into extracellular loosely bound and free water in order to mitigate an increase in tissue stiffness.
3. Advanced Glycation End Products
It has been shown that tissue stiffness increases as a result of tissue crosslinking by advanced glycation end products (AGEs). Dehydration and glycation may be considered as subsequent steps during the aging process [11][20]. A decrease in bound water content and increase in AGEs during aging were reported in the cortical bones of rats [21]. The accumulation of AGEs and reduction of water content in the ovine intervertebral disc was also observed [22]. Crosslinking of elastin and collagen by AGEs was associated with stiffening of the arterial wall with aging [5][9] and AGE cross-link breakers can be suggested as “de-stiffening” drugs [23][24].
4. De-Stiffening
4.1. Polyphenols
Polyphenols have been suggested as inhibitors of AGEs [32][33][34][35][36][37]. Diabetes leads to glycocalyx degradation, and the maintenance or restoration of the integrity of the glycocalyx is an important therapeutic target. Polyphenols, such as resveratrol, quercetin, and (−)-epicatechin, can promote the swelling of the highly hydrated endothelial glycocalyx, increasing its thickness and decreasing stiffness, as a result of cystic fibrosis transmembrane conductance regulator (CFTR) activation [35]. CFTR regulates the transport of ions and water across the epithelial barrier. Therefore, it can be concluded that polyphenols decrease stiffness and increase hydration.
Higher flavonoid consumption has been associated with a lower arterial stiffness. Isoflavones and anthocyanins have been recommended as the most efficient for the improvement of vascular health. Polyphenols activate endothelial nitric oxide synthase and increase endothelial synthesis and the production of nitric oxide [37], in such a way regulating arterial stiffness and preventing cardiovascular diseases. Anthocyanins are not only effective in the improvement of vascular health but also have antiviral properties. Inhibitory effects of anthocyanins [38] on different pathways involved in the virus life cycle have also been recently reviewed.
In an in-vitro study, it was shown that bovine coronavirus infections can be neutralized by theaflavins extracted from black tea [39]. Ueda et al. [40] reported that tannins decreased viral infectivity against 12 different viruses, which were both enveloped and non-enveloped. Extracts from persimmon (Diospyros kaki), which contains 22% of a food supplement persimmon tannin, demonstrated the highest antiviral effect. The authors demonstrated that extracts from persimmon inhibited attachment of the virus to cells. Pretreatment or posttreatment of cells with the persimmon extracts before or after virus infection did not inhibit virus replication. It was suggested that the antiviral effect of persimmon tannin can be explained by possible influenza virus protein aggregation. However, inhibited attachment of the virus to cells can also be explained by relative cell de-stiffening by persimmon extract due to changes in the ratio of virus and cell stiffnesses. Similarly, virus de-stiffening decreases platelet adhesion and prevents thrombus formation [7]. It has been shown that polyphenols can act as plasticizers; for example, tea-polyphenol-treated skin collagen demonstrated resistance to dehydration [41].
4.2. Omega-3 Fatty Acids
A more hydrated and intact intervertebral disc tissue has been observed in rats who consumed an omega-3 fatty acids diet [42]. At the same time, omega-3 fatty acids have been reported to be a de-stiffening agent; long-term omega-3 fatty acid (fish oil) supplementation in a diet decreases arterial stiffness [43]. Fish oil supplementation decreases large arterial stiffness in overweight hypertensive patients [44]. Schmidt et al. analyzed the effect of omega-3 polyunsaturated fatty acids on the cytoskeleton and concluded that n-3 PUFAs regulated cytoskeleton-associated gene expression [45]. It has also been reported that Rho GTPase inhibited the metastatic ability of human prostate cancer cell line PC-3 by omega-3 polyunsaturated fatty acids [46]. It was also recently suggested that arachidonic acid, eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA) are able to inactivate enveloped viruses and increase resistance and recovery from SARS-CoV-2, SARS, and Middle East respiratory syndrome (MERS) infections [47]. Very low blood levels of the sum of eicosapentaenoic acid and docosahexaenoic acid were observed in USA, Italy, UK, and Brazil [48]. A high number of COVID-19 patients were reported in these countries.
4.3. Drugs
Arterial wall stiffness is related to cardiovascular diseases [49]. For example, platelet adhesion and thrombus formation depend on arterial wall stiffness and hydration [50][51], Figure 1. An increase in substrate stiffness leads to endothelial monolayer disruption by increased cellular traction stresses and to increased endothelium permeability by leukocytes resulting in the development of atherosclerosis. It has been suggested that simvastatin can reduce matrix stiffness due to the decrease in RhoA activity in regulating contractility of the actin cytoskeleton and decrease in cellular traction forces. Simvastatin treatment resulted in improved endothelial barrier integrity and decreased endothelium permeability by decreasing cell–cell junction size in endothelial monolayers. It has been suggested that statins are able to maintain endothelial barrier integrity and prevent the development of atherosclerosis as a result of leukocyte penetration through endothelial monolayer damaged by tissue stiffness [52]. Similarly, it is possible that coronavirus penetration in novel COVID-19 disease can be increased for older patients due to increased age-related tissue stiffness. It was already reported that statins may decrease the fatality rate of MERS infection [53] and may be used in the treatment of COVID-19 [54]. The activities of various statins—simvastatin, atorvastatin, rusovastatin, simvastatin, lovastatin, mevastatin—and the possibility to use statins in treatment of COVID-19 was recently suggested [55].
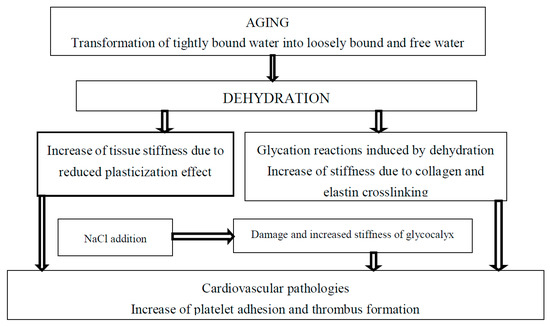
Figure 1. The effect of tightly bound water transformations on tissue stiffness and related cardiovascular diseases.
4.4. Chitosan and Its Derivatives
It can be expected that AGE content will increase in the presence of high-molecular-weight chitosan [58]. However, chitosan oligosaccharides demonstrated angiotensin-converting enzyme inhibition activity [59]. It has also been reported that substitution of the hydrogen atom at the C-6 position of pyranose residue by the aminoethyl group promotes angiotensin-converting enzyme inhibitory effects of chitooligosaccharides [60]. It is known that angiotensin-converting enzyme inhibition dilates the blood vessels and decreases high blood pressure. Angiotensin-converting enzyme inhibitors can also be used to decrease arterial stiffness. Aminoethyl-chitooligosaccharide also suppresses proliferation of human lung A549 cancer cells [61]. Chitosan oligosaccharides suppress pathogenic microorganisms’ adhesion to cells and in such a way can be considered inhibitors of initial stages of infection processes [62]. Chitosan oligosaccharides are also able to penetrate inside pathogenic organisms and prevent their reproduction.
N-(2-hydroxypropyl)-3-trimethylammonium chitosan chloride (HTCC) was proposed as an inhibitor of human coronavirus HCoV-NL63. It was suggested that HTCC blocks human coronavirus interaction with the cellular receptor, angiotensin-converting enzyme type 2 (ACE2) proteins [63].
4.5. Vitamins
Association between vitamin D deficiency and arterial stiffness determined from pulse wave velocity measurements was reported in a number of publications. Vitamin D deficiency was associated with the increase in arterial stiffness in children with type 1 and type 2 diabetes and with chronical kidney disease as well as in the elderly population [64][65][66][67][68][69][70][71].
Generally, the results from randomized controlled trials have been inconsistent and more large population studies should be conducted. It was reported that high-dose—but not low-dose—vitamin D supplementation was able to decrease arterial stiffness [70]. The decrease in arterial stiffness was related to the suggestion that 25-hydroxyvitamin D inhibits macrophage stimulation [72] and with the suppression of endothelin-induced vascular smooth muscle cell proliferation [73].
It has been recently reported that vitamin D supplementation could reduce the risk of death from the current COVID-19 epidemic. It is commonly accepted that vitamin D deficiency weakens the immune system. Vitamin D deficiency also increases tissue stiffness, and maintaining tight junctions was mentioned as one of mechanisms of vitamin D function [74]. Maintaining tight junctions prevents coronavirus infection by preventing tissue permeability.
Vitamin D intake is very low in Italy, Spain and France [75]. The most severe COVID-19 epidemic was observed in these countries, although, at present, there are not enough data confirming the link between vitamin D and COVID-19.
Vitamin E prevented an increase in femoral artery stiffness in diabetic Wistar rats [76]. It was reported that consumption of vitamin E for 2 months resulted in a decrease in carotid femoral pulse wave velocity and augmentation index in a group of 36 healthy men [77]. Arterial stiffness decreased as a result of long-term treatment with vitamin E in combination with vitamin C, coenzyme Q10, and selenium [78]. The effect of vitamin E in combination with pravastatin and hymocysteine on arterial stiffness was studied with the aim to reduce increased aortic and carotid artery stiffness in patients with end-stage renal disease [79].
It was suggested that supplementation with vitamin D and vitamin E may increase resistance to novel coronavirus SARS-CoV-2 [80].
Vitamin C decreases arterial stiffness [81][82]. A combination of vitamin C with various other drugs could be beneficial for COVID-19 patients [83].
Vitamin K2 decreases arterial stiffness by preventing and reversing calcification of arteries and inhibition of inflammation in the vascular wall [68][84][85][86].
It was reported that chitosan oligosaccharide ascorbate corrected deficiency in vitamin A, E, C and B groups [62].
4.6. Salt Restriction
It was observed that salt consumption increases arterial stiffness and the risk of cardiovascular diseases [87][88][89][90][91], and salt also increases tightly bound water mobility due to the decrease in binding energy in pork meat [92]. Therefore, it is possible to suggest a correlation between arterial stiffness and increased tightly bound water relaxation time (increased protein mobility) measured by low field nuclear magnetic resonance.
Optimal hydration retards development of age-related diseases [93]. Increase in extracellular sodium concentration results in vascular endothelial cells’ activation and leads to shrinking and increased stiffness of glycocalyx [94][95][96]. Dehydration and related increases in sodium concentration also lead to the expression of inflammation mediators vascular cell adhesion molecule 1 (VCAM-1), endothelial-leukocyte adhesion molecule 1 (E-selectin), and monocyte chemoattractant protein 1 (MCP-1) [97][98]. High sodium concentration decreases NO release and promotes endothelial release of the pro-inflammatory cytokines IL-1ß and TNFα [99]. Increased endothelial stiffening and diminished glycocalyx coverage were observed in diabetic mice if compared with control. Glycocalyx coverage decreases in advanced age with diabetes progression [100].
4.7. Dehydration, Stiffness and NF-κB Activation
Dehydration activates NF-κB [101]. Dehydration also results in enhanced tissue stiffness. Ishihara et al. reported that increased substrate stiffness leads to expression of inflammatory genes via NF-κB activation and results in actomyosin contractions, which triggers NF-κB activation [102]. The actomyosin contractions are induced by phosphorylation of myosin regulatory light chain (MRLC). The inhibition of MRLC phosphorylation by Rho kinase inhibitor Y27632 reduced the activity of NF-Κb. NF-κB activation may be associated with numerous diseases, including cancer and fibrosis development.
Matrix stiffening promotes F-actin polymerization regulated by small GTPase RhoA, Figure 2, and α smooth muscle actin (α-SMA)-containing stress-fiber formation. Inhibition of RhoA inhibits stress-fiber formation. Water molecules have an important role in F-actin polymerization, and it was shown that fewer water molecules were associated with actin when it polymerized [103]. Thus, the polymerization of actin and dehydration can be associated with an increase in tissue stiffness. The mechanical properties of actin can be regulated using various agents. For example, jasplakinolide, cytochalasin D, and latrunculin A were used to directly modulate actin polymerization of cultured rat airway smooth muscle cells [104].
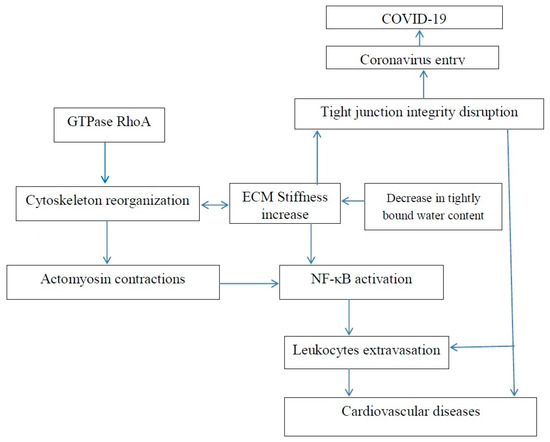
Figure 2. Increase in extracellular matrix stiffness results in both an increased risk of age-related cardiovascular disease and an increased risk of COVID-19.
Rho GTPases control cell proliferation and cytoskeleton remodeling and they are commonly hyper-expressed in tumors. Y-27632 also suppresses formation of stress fibers in cultured cells and decreases hypertension in several hypertensive rat models [105][106][107]. Pretreatment with ibuprofen was reported to prevent F-actin upregulation in articular chondrocytes from New Zealand White rabbits due to inhibition of RhoA pathway [108]. In the treatment of COVID-19, a number of doctors recommend using paracetamol rather than ibuprofen [109]. ROCK has been suggested as a potential target for the treatment of a number of diseases, such as cancer, neuronal degeneration, vascular diseases, kidney failure, asthma, acute lung injury, glaucoma, osteoporosis, erectile dysfunction, and insulin resistance [110][111][112][113][114][115]. For example, it has been recently reported that, in the treatment of acute lung injury, the Rho kinase inhibitor decreases inflammation, immune cell migration, apoptosis, coagulation, contraction, and cell adhesion in pulmonary endothelial cells. It means that all the above-mentioned diseases may be dependent on tissue stiffness related to time-dependent interaction of water molecules and tissue macromolecules. F-Actin reorganization is related to lung inflammation via increased blood neutrophil adhesion and migration. Pretreatment by cytochalasin B (CB) can block F-actin reorganization [116]. Currently, it is generally recognized that many questions remain to be investigated in order to understand Rho GTPase function, and it would be beneficial if tissue stiffness and the content of bound water were taken into account.
4.8. COVID-19 Age Dependence
It is very important that children have an extremely low rate of hospitalization presented by the U.S. Department of Health and Human Services in Figure 3. This fact demonstrates that people with soft tissues have a low chance of becoming ill with both age-related diseases and COVID-19.
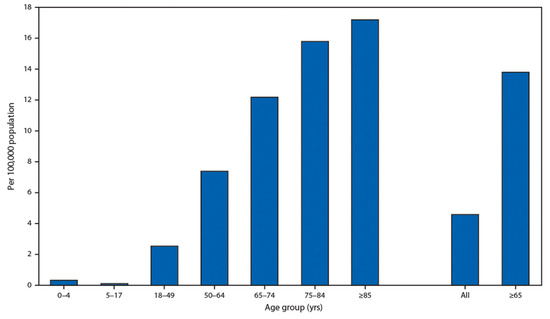
Figure 3. Coronavirus disease 2019 (COVID-19)-associated hospitalization rates, by age group, COVID-NET, 14 states, 1–28 March 2020. Cited from Garg S., Kim L., Whitaker M., et al. Hospitalization Rates and Characteristics of Patients Hospitalized with Laboratory-Confirmed Coronavirus Disease 2019, COVID-NET, 14 States, 1–30 March 2020. Morb Mortal Wkly Rep (MMWR) 2020; 69:458–464. doi:10.15585/mmwr.mm6915e3 MMWR and Morbidity and Mortality Weekly Report are service marks of the U.S. Department of Health and Human Services.
It has been also reported that obesity as an underlying condition was observed in 48.3% COVID-19 patients. It was earlier demonstrated that obesity results in increased arterial stiffness [117].
References
- Discher, D.E.; Janmey, P.; Wang, Y.-L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143.
- Khetan, S.; Guvendiren, M.; Legant, W.; Cohen, D.M.; Chen, C.S.; Burdick, J.A. Degradation-mediated cellular traction directs stem cell fate in covalently crosslinked three-dimensional hydrogels. Nat. Mater. 2013, 12, 458–465.
- Kerch, G. The potential of chitosan and its derivatives in prevention and treatment of age-related diseases. Mar. Drugs 2015, 13, 2158–2182.
- Lampi, M.C.; Reinhart-King, C.A. Targeting extracellular matrix stiffness to attenuate disease: From molecular mechanisms to clinical trials. Sci. Transl. Med. 2018, 10, eaao0475.
- Kohn, J.C.; Lampi, M.C.; Reinhart-King, C.A. Age-related vascular stiffening: Causes and consequences. Front. Genet. 2015, 6, 112.
- Iulita, M.F.; Noriega de la Colina, A.; Girouard, H. Arterial stiffness, cognitive impairment and dementia: Confounding factor or real risk? J. Neurochem. 2018, 144, 527–548.
- Pase, M.P.; Herbert, A.; Grima, N.A.; Pipingas, A.; O’Rourke, M.F. Arterial stiffness as a cause of cognitive decline and dementia: A systematic review and meta-analysis. Intern. Med. J. 2012, 42, 808–815.
- Rabkin, S.W. Arterial stiffness: Detection and consequences in cognitive impairment and dementia of the elderly. J. Alzheimers Dis. 2012, 32, 541–549.
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200.
- Kerch, G. Polymer hydration and stiffness at biointerfaces and related cellular processes. Nanomed. Nanotechnol. 2018, 14, 13–25.
- Kerch, G. Distribution of tightly and loosely bound water in biological macromolecules and age-related diseases. Int. J. Biol. Macromol. 2018, 118, 1310–1318.
- Creecy, A.; Uppuganti, S.; Girard, M.R.; Schlunk, S.G.; Amah, C.; Granke, M.; Unal, M.; Does, M.D.; Nyman, J.S. The age-related decrease in material properties of BALB/c mouse long bones involves alterations to the extracellular matrix. Bone 2020, 130, 115126.
- Granke, M.; Does, M.D.; Nyman, J.S. The role of water compartments in the material properties of cortical bone. Calcif. Tissue Int. 2015, 97, 292–307.
- Burr, D.B. Changes in bone matrix properties with aging. Bone 2019, 120, 85–93.
- Serra-Prat, M.; Lorenzo, I.; Palomera, E.; Ramírez, S.; Yébenes, J.C. Total body water and intracellular water relationships with muscle strength, frailty and functional performance in an elderly population. J. Nutr. Health Aging 2019, 23, 96–101.
- Yamada, Y.; Yoshida, T.; Yokoyama, K.; Watanabe, Y.; Miyake, M.; Yamagata, E.; Yamada, M.; Kimura, M.; Kyoto-Kameoka Study. The extracellular to intracellular water ratio in upper legs is negatively associated with skeletal muscle strength and gait speed in older people. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 293–298.
- Kim, E.J.; Choi, M.J.; Lee, J.H.; Oh, J.E.; Seo, J.W.; Lee, Y.K.; Yoon, J.W.; Kim, H.J.; Noh, J.W.; Koo, J.R. Extracellular fluid/intracellular fluid volume ratio as a novel risk indicator for all-cause mortality and cardiovascular disease in hemodialysis patients. PLoS ONE 2017, 12, e0170272.
- Lee, J.Y.; Ryu, H.S.; Yoon, S.S.; Kim, E.H.; Yoon, S.W. Extracellular-to-intracellular fluid volume ratio as a prognostic factor for survival in patients with metastatic cancer. Integr. Cancer Ther. 2019, 18, 1534735419847285.
- Ohashi, Y.; Joki, N.; Yamazaki, K.; Kawamura, T.; Tai, R.; Oguchi, H.; Yuasa, R.; Sakai, K. Changes in the fluid volume balance between intra- and extracellular water in a sample of Japanese adults aged 15–88 yr old: A cross-sectional study. Am. J. Physiol. Renal 2018, 314, F614–F622.
- Bridelli, M.G.; Stani, C.; Bedotti, R. Fourier transform infrared conformational investigation of type I collagen aged by in vitro induced dehydration and non-enzymatic glycation treatments. J. Biol. Res. Bollettino Società Italiana Biologia Sperimentale 2017, 90, 45–50.
- Uppuganti, S.; Granke, M.; Makowski, A.J.; Does, M.D.; Nyman, J.S. Age-related changes in the fracture resistance of male Fischer F344 rat bone. Bone 2016, 83, 220–232.
- Jazini, E.; Sharan, A.D.; Morse, L.J.; Dyke, J.P.; Aronowitz, E.A.; Chen, L.K.H.; Tang, S.Y. Alterations in magnetic resonance imaging T2 relaxation times of the ovine intervertebral disc due to non-enzymatic glycation. Spine 2012, 37, E209–E215.
- Chen, Y.; Shen, F.; Liu, J.; Yang, G.Y. Arterial stiffness and stroke: De-stiffening strategy, a therapeutic target for stroke. Stroke Vasc. Neurol. 2017, 2, 65–72.
- Boutouyrie, P.; Lacolley, P.; Briet, M.; Regnault, V.; Stanton, A.; Laurent, S.; Mahmud, A. Pharmacological modulation of arterial stiffness. Drugs 2011, 71, 1689–1701.
- Sell, D.R.; Monnier, V.M. Molecular basis of arterial stiffening: Role of glycation—A Mini-review. Gerontology 2012, 58, 227–237.
- Thomas, M.C.; Baynes, J.W.; Thorpe, S.R.; Cooper, M.E. The role of AGEs and AGE inhibitors in diabetic cardiovascular disease. Curr. Drug Targets 2005, 6, 453–474.
- Wu, M.S.; Liang, J.T.; Lin, Y.D.; Wu, E.T.; Tseng, Y.Z.; Chang, K.C. Aminoguanidine prevents the impairment of cardiac pumping mechanics in rats with streptozotocin and nicotinamide-induced type 2 diabetes. Br. J. Pharmacol. 2008, 154, 758–764.
- Chang, K.C.; Tseng, C.D.; Wu, M.S.; Liang, J.T.; Tsai, M.S.; Cho, Y.L.; Tseng, Y.Z. Aminoguanidine prevents arterial stiffening in a new rat model of type 2 diabetes. Eur. J. Clin. Invest. 2006, 36, 528–535.
- Sell, D.R.; Nelson, J.F.; Monnier, V.M. Effect of chronic aminoguanidine treatment on agerelated glycation, glycoxidation, and collagen cross-linking in the Fischer 344 rat. J. Gerontol. Biol. Sci. 2001, 56A, B405–B411.
- Brownlee, M.; Vlassara, H.; Kooney, T.; Ulrich, P.; Cerami, A. Aminoguanidine prevents diabetes-induced arterial wall protein crosslinking. Science 1986, 232, 1629–1632.
- Roorda, M.M. Therapeutic interventions against accumulation of advanced glycation end products (AGEs). Glycative Stress Res. 2017, 4, 132–143.
- De Bruyne, T.; Steenput, B.; Roth, L.; De Meyer, G.R.Y.; Nunes dos Santos, C.; Valentová, K.; Dambrova, M.; Hermans, N. Dietary polyphenols targeting arterial stiffness: Interplay of contributing mechanisms and gut microbiome-related metabolism. Nutrients 2019, 11, 578.
- Xie, Y.; Chen, X. Structures required of polyphenols for inhibiting advanced glycation end products formation. Curr. Drug Metab. 2013, 14, 414–431.
- Khangholi, S.; Majid, F.A.A.; Berwary, N.J.A.; Ahmad, F.; Aziz, R.B.A. The mechanisms of inhibition of advanced glycation end products formation through polyphenols in hyperglycemic condition. Planta Med. 2016, 82, 32–45.
- Peters, W.; Kusche-Vihrog, K.; Oberleithner, H.; Schillers, H. Cystic fibrosis transmembrane conductance regulator is involved in polyphenol-induced swelling of the endothelial glycocalyx. Nanomed. Nanotechnol. 2015, 11, 1521–1530.
- Lilamand, M.; Kelaiditi, E.; Guyonnet, S.; Antonelli Incalzi, R.; Raynaud-Simon, A.; Vellas, B.; Cesari, M. Flavonoids and arterial stiffness: Promising perspectives. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 698–704.
- Schmitt, C.A.; Dirsch, V.M. Modulation of endothelial nitric oxide by plant-derived products. Nitric Oxide 2009, 21, 77–91.
- Mohammadi Pour, P.; Fakhri, S.; Asgary, S.; Farzaei, M.H.; Echeverría, J. The signaling pathways, and therapeutics of antiviral agents: Focusing on the antiviral approaches and clinical perspectives of anthocyanins in the management of viral diseases. Front. Pharmacol. 2019, 10, 1207.
- Clark, K.J.; Grant, P.G.; Sarr, A.B.; Belakere, J.R.; Swaggerty, C.L.; Phillips, T.D.; Woode, G.N. An in vitro study of theaflavins extracted from black tea to neutralize bovine rotavirus and bovine coronavirus infections. Vet. Microbiol. 1998, 63, 147–157.
- Ueda, K.; Kawabata, R.; Irie, T.; Nakai, Y.; Tohya, Y.; Sakaguchi, T. Inactivation of pathogenic viruses by plant-derived tannins: Strong effects of extracts from persimmon (Diospyros kaki) on a broad range of viruses. PLoS ONE 2013, 8, e5534.
- Han, Y.; Jiang, Y.; Hu, J. Tea-polyphenol treated skin collagen owns coalesced adaptive-hydration, tensile strength and shape-memory property. Int. J. Biol. Macromol. 2020, 158, 1–8.
- NaPier, Z.; Kanim, L.; Arabi, Y.; Salehi, K.; Sears, B.; Perry, M.; Kim, S.; Sheyn, D.; Bae, H.W.; Glaeser, J.D. Omega-3 fatty acid supplementation reduces intervertebral disc degeneration. Med. Sci. Monit. 2019, 25, 9531–9537.
- Pase, M.P.; Grima, N.A.; Sarris, J. The effects of dietary and nutrient interventions on arterial stiffness: A systematic review. Am. J. Clin. Nutr. 2011, 93, 446–454.
- Wang, S.; Ma, A.Q.; Song, S.W.; Quan, Q.H.; Zhao, X.F.; Zheng, X.H. Fish oil supplementation improves large arterial elasticity in overweight hypertensive patients. Eur. J. Clin. Nutr. 2008, 62, 1426–1431.
- Schmidt, S.; Willers, J.; Riecker, S.; Möller, K.; Schuchadt, J.P.; Hahn, A. Effect of omega-3 polyunsaturated fatty acids on the cytoskeleton: An open-label intervention study. Lipids Health Dis. 2015, 14, 4.
- Yi, L.; Zhang, Q.; Mi, M. Role of Rho GTPase in inhibiting metastatic ability of human prostate cancer cell line PC-3 by omega-3 polyunsaturated fatty acid [abstract]. Ai Zheng 2007, 12, 1281–1286.
- Das, U.N. Can Bioactive Lipids Inactivate Coronavirus (COVID-19)? Arch. Med. Res. 2020, 51, 282–286.
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N., Jr. Global survey of the omega-3 fatty acids, docosahexaenoic acid and eicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 2016, 63, 132–152.
- Huveneers, S.; Daemen, M.J.A.P.; Hordijk, P.L. Between Rho(k) and a hard place: The relation between vessel wall stiffness, endothelial contractility, and cardiovascular disease. Circ. Res. 2015, 116, 895–908.
- Kerch, G.; Zicans, J.; Merijs Meri, R.; Stunda-Ramava, A.; Jakobsons, E. The use of thermal analysis in assessing the effect of bound water content and substrate rigidity on prevention of platelet adhesion. J. Therm. Anal. Calorim. 2015, 120, 533–539.
- Kerch, G.; Chausson, M.; Gautier, S.; Meri, R.M.; Zicans, J.; Jakobsons, E.; Joner, M. Heparin-like polyelectrolyte multilayer coatings based on fungal sulfated chitosan decrease platelet adhesion due to the increased hydration and reduced stiffness. Biomater. Tissue Technol. 2015, 1, 1–5.
- Lampi, M.C.; Faber, C.J.; Huynh, J.; Bordeleau, F.; Zanotelli, M.R.; Reinhart-King, C.A. Simvastatin ameliorates matrix stiffness-mediated endothelial monolayer disruption. PLoS ONE 2016, 11, e0147033.
- Yuan, S. Statins may decrease the fatality rate of Middle East respiratory syndrome infection. mBio 2015, 6, e01120-15.
- Fedson, D.S.; Opal, S.M.; Rordamc, O.M. Hiding in plain sight: An approach to treating patients with severe COVID-19 infection. mBio 2020, 11, e00398-20.
- Bifulco, M.; Gazzerro, P. Statins in coronavirus outbreak: It’s time for experimental and clinical studies. Pharmacol. Res. 2020, 156, 104803.
- Janić, M.; Lunder, M.; Sabovič, M. Arterial stiffness and cardiovascular therapy. Biomed. Res. Int. 2014, 2014, 621437.
- Shahin, Y.; Khan, J.A.; Chetter, I. Angiotensin converting enzyme inhibitors effect on arterial stiffness and wave reflections: A meta-analysis and meta-regression of randomized controlled trials. Atherosclerosis 2012, 221, 18–33.
- Rakcejeva, T.; Rusa, K.; Dukalska, L.; Kerch, G. Effect of chitosan and chitooligosaccharide lactate on free lipids and reducing sugars content and on wheat bread firming. Eur. Food Res. Technol. 2011, 232, 123–128.
- Hong, S.P.; Kim, M.H.; Oh, S.W.; Han, C.H.; Kim, Y.H. ACE inhibitory and antihypertensive effect of chitosan oligosaccharides in SHR. Korean J. Food Sci. Technol. 1998, 30, 1476–1479.
- Ngo, D.N.; Qian, Z.J.; Je, J.Y.; Kim, M.M.; Kim, S.K. Aminoethyl chitooligosaccharides inhibit the activity of angiotensin converting enzyme. Process Biochem. 2008, 43, 119–123.
- Ngo, D.H.; Ngo, D.N.; Kim, S.K.; Vo, T.S. Antiproliferative effect of aminoethyl-chitooligosaccharide on human lung A549 cancer cells. Biomolecules 2019, 9, 195.
- Kirilenko, Y.K.; Dushkova, Z.G.; Cherkasova, E.I.; Sigilietov, A.E. Chitosan oligomer and ascorbic acid salt in compensation of deficiency of some micronutrients. Adv. Chitin Sci. 2007, 10, 487–491.
- Milewska, A.; Kaminski, K.; Ciejka, J.; Kosowicz, K.; Zeglen, S.; Wojarski, J.; Nowakowska, M.; Szczubiałka, K.; Pyrc, K. HTCC: Broad range inhibitor of coronavirus entry. PLoS ONE 2016, 11, e0156552.
- Patange, A.R.; Valentini, R.P.; Du, W.; Pettersen, M.D. Vitamin D deficiency and arterial wall stiffness in children with chronic kidney disease. Pediatr. Cardiol. 2012, 33, 122–128.
- Lieberman, R.; Wadwa, R.P.; Nguyen, N.; Bishop, F.K.; Reinick, C.; Snell-Bergeon, J.K.; Maahs, D.M. The association between vitamin D and vascular stiffness in adolescents with and without type 1 diabetes. PLoS ONE 2013, 8, e77272.
- Jha, P.; Dolan, L.M.; Khoury, P.R.; Urbina, E.M.; Kimball, T.R.; Shah, A.S. Low serum vitamin D levels are associated with increased arterial stiffness in youth with type 2 diabetes. Diabetes Care 2015, 38, 1551–1557.
- Chang, J.; Ye, X.-G.; Hou, Y.-P.; Wu, J.-L.; Li, S.L.; Sun, Q.-M. Vitamin D level is associated with increased left ventricular mass and arterial stiffness in older patients with impaired renal function. Med. Sci. Monitor. 2015, 21, 3993–3999.
- Mozos, I.; Stoian, D.; Luca, C.T. Crosstalk between vitamins A, B12, D, K, C, and E status and arterial stiffness. Biomark. Cardiovasc. Dis. 2017, 2017, 14.
- Dong, Y.; Stallmann-Jorgensen, I.S.; Pollock, N.K.; Harris, R.A.; Keeton, D.; Huang, Y.; Li, K.; Bassali, R.; Guo, H.G.; Thomas, J.; et al. A 16-week randomized clinical trial of 2000 international units daily vitamin D3 supplementation in black youth: 25-Hydroxyvitamin D, adiposity, and arterial stiffness. J. Clin. Endocrinol. Metabol. 2010, 95, 4584–4591.
- Zaleski, A.; Panza, G.; Swales, H.; Arora, P.; Newton-Cheh, C.; Wang, T.; Thompson, P.G.; Taylor, B. High-dose versus low-dose vitamin D supplementation and arterial stiffness among individuals with prehypertension and vitamin D deficiency. Dis. Markers 2015, 2015, 7.
- McGreevy, C.; Barry, M.; Davenport, C.; Byrne, B.; Donaghy, C.; Colliers, G.; Tormey, W.; Smith, D.; Bennett, K.; Williams, D. The effect of vitamin D supplementation on arterial stiffness in an elderly community-based population. J. Am. Soc. Hypertens. 2015, 9, 176–183.
- Helming, L.; Bose, J.; Ehrchen, J.; Schiebe, S.; Frahm, T.; Geffers, R.; Probst-Kepper, M.; Balling, R.; Lengeling, A. 1α,25-dihydroxyvitamin D3 is a potent suppressor of interferon γ-mediated macrophage activation. Blood 2005, 106, 4351–4358.
- Chen, S.; Law, C.S.; Gardner, D.G. Vitamin D-dependent suppression of endothelin-induced vascular smooth muscle cell proliferation through inhibition of CDK2 activity. J. Steroid Biochem. Mol. Biol. 2010, 118, 135–141.
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths. Nutrients 2020, 12, 988.
- Lips, P.; Cashman, K.D.; Lamberg-Allardt, C.; Bischoff-Ferrari, H.A.; Obermayer-Pietsch, B.; Bianchi, M.L.; Stepan, J.; El-Haj Fuleihan, G.; Bouillon, R. Current vitamin D status in European and Middle East countries and strategies to prevent vitamin D deficiency: A position statement of the European Calcified Tissue Society. Eur. J. Endocrinol. 2019, 180, P23–P54.
- Wigg, S.J.; Tare, M.; Forbes, J.; Copper, M.E.; Thomas, M.C.; Coleman, H.A.; Parkington, H.C.; O’Brien, R.C. Early vitamin E supplementation attenuates diabetes-associated vascular dysfunction and the rise in protein kinase C-β in mesenteric artery and ameliorates wall stiffness in femoral artery of Wistar rats. Diabetologia 2004, 47, 1038–1046.
- Rasool, A.H.G.; Rahman, A.R.A.; Yuen, K.H.; Wong, A.R. Arterial compliance and vitamin E blood levels with a selfemulsifying preparation of tocotrienol rich vitamin E. Arch. Pharm. Res. 2008, 31, 1212–1217.
- Shargorodsky, M.; Debby, O.; Matas, Z.; Zimlichman, R. Effect of long-term treatment with antioxidants (vitamin C, vitamin E, coenzyme Q10 and selenium) on arterial compliance, humoral factors and inflammatory markers in patients with multiple cardiovascular risk factors. Nutr. Metab. 2010, 7, 55.
- Veringa, S.J.E.; Nanayakkara, P.W.B.; Van Ittersum, F.J.; Vegting, I.L.; van Guldener, C.; Smulders, Y.M.; ter Wee, P.M.; Stehouwer, C.D.A. Effect of a treatment strategy consisting of pravastatin, vitamin E, and homocysteine lowering on arterial compliance and distensibility in patients with mild-to-moderate chronic kidney disease. Clin. Nephrol. 2012, 78, 263–272.
- Wang, L.S.; Wang, Y.R.; Ye, D.W.; Liu, Q.Q. A review of the 2019 Novel Coronavirus (COVID-19) based on current evidence. Int. J. Antimicrob. Agents 2020, 105948.
- Wilkinson, I.B.; Megson, I.L.; MacCallum, H.; Sogo, N.; Cockcroft, J.R.; Webb, D.J. Oral vitamin C reduces arterial stiffness and platelet aggregation in humans. J. Cardiovasc. Pharmacol. 1999, 34, 690–693.
- Mullan, B.A.; Young, I.S.; Fee, H.; McCance, D.R. Ascorbic acid reduces blood pressure and arterial stiffness in type 2 diabetes. Hypertension 2002, 40, 804–809.
- Pawar, A.Y. Combating devastating COVID -19 by drug repurposing. Int. J. Antimicrob. Agents 2020, 17, 105984.
- Knapen, M.H.; Braam, L.A.; Drummen, N.E.; Bekers, O.; Hoeks, A.P.; Vermeer, C. Menaquinone-7 supplementation improves arterial stiffness in healthy postmenopausal women: A double-blind randomised clinical trial. Thromb. Haemost. 2015, 113, 1135–1144.
- Beulens, J.W.J.; Bots, M.L.; Atsma, F.; Bartelink, M.-L.E.L.; Prokop, M.; Geleijnse, J.M.; Witteman, J.C.M.; Grobbee, D.E.; van der Schouw, Y.T. High dietary menaquinone intake is associated with reduced coronary calcification. Atherosclerosis 2009, 203, 489–493.
- Vossen, L.M.; Schurgers, L.J.; van Varik, B.J.; Kietselaer, B.L.J.H.; Vermeer, C.; Meeder, J.G.; Rahel, B.M.; van Cauteren, Y.J.M.; Hoffland, G.A.; Rennenberg, R.J.W.M.; et al. Menaquinone-7 supplementation to reduce vascular calcification in patients with coronary artery disease: Rationale and study protocol (VitaK-CAC Trial). Nutrients 2015, 7, 8905–8915.
- Dickinson, K.M.; Clifton, P.M.; Burrell, L.M.; Barrett, P.H.R.; Keogh, J.B. Postprandial effects of a high salt meal on serum sodium, arterial stiffness, markers of nitric oxide production and markers of endothelial function. Atherosclerosis 2014, 232, 211–216.
- He, F.J.; Marciniak, M.; Visagie, E.; Markandu, N.D.; Anand, V.; Dalton, R.N.; MacGregor, G.A. Effect of modest salt reduction on blood pressure, urinary albumin, and pulse wave velocity in white, black, and Asian mild hypertensives. Hypertension 2009, 54, 482–488.
- Todd, A.S.; Macginley, R.J.; Schollum, J.B.; Johnson, R.J.; Williams, S.M.; Sutherland, W.H.; Mann, J.I.; Walker, R.J. Dietary salt loading impairs arterial vascular reactivity. Am. J. Clin. Nutr. 2010, 91, 557–564.
- Gates, P.E.; Tanaka, H.; Hiatt, W.R.; Seals, D.R. Dietary sodium restriction rapidly improves large elastic artery compliance in older adults with systolic hypertension. Hypertension 2004, 44, 35–41.
- Wu, C.F.; Liu, P.Y.; Wu, T.J.; Hung, Y.; Yang, S.P.; Lin, G.M. Therapeutic modification of arterial stiffness: An update and comprehensive review. World J. Cardiol. 2015, 7, 742–753.
- McDonnell, C.K.; Allen, P.; Duggan, E.; Arimi, J.M.; Casey, E.; Duane, G.; Lyng, J.G. The effect of salt and fibre direction on water dynamics, distribution and mobility in pork muscle: A low field NMR study. Meat Sci. 2013, 95, 51–58.
- Allen, M.D.; Springer, D.A.; Burg, M.B.; Boehm, M.; Dmitrieva, N.I. Suboptimal hydration remodels metabolism, promotes degenerative diseases, and shortens life. JCI Insight 2019, 4, e130949.
- Oberleithner, H.; Riethmüller, C.; Schillers, H.; MacGregor, G.A.; de Wardener, H.E.; Hausberg, M. Salt overload damages the glycocalyx sodium barrier of vascular endothelium. Pflugers Arch. 2011, 462, 519–528.
- Oberleithner, H.; Riethmüller, C.; Schillers, H.; MacGregor, G.A.; de Wardener, H.E.; Hausberg, M. Plasma sodium stiffens vascular endothelium and reduces nitric oxide release. Proc. Natl. Acad. Sci. USA 2007, 104, 16281–16286.
- Liew, H.; Roberts, M.A.; MacGinley, R.; McMahon, L.P. Endothelial glycocalyx in health and kidney disease: Rising star or false Dawn? Nephrology (Carlton) 2017, 22, 940–946.
- Dmitrieva, N.I.; Burg, M.B. Elevated sodium and dehydration stimulate inflammatory signaling in endothelial cells and promote atherosclerosis. PLoS ONE 2015, 10, e0128870.
- Wild, J.; Soehnlein, O.; Dietel, B.; Urschel, K.; Garlichs, C.D.; Cicha, I. Rubbing salt into wounded endothelium: Sodium potentiates proatherogenic effects of TNF-α under non-uniform shear stress. Thromb. Haemost. 2014, 112, 183–195.
- Schierke, F.; Wyrwoll, M.; Wisdorf, M.; Niedzielski, L.; Maase, M.; Ruck, T.; Meuth, S.G.; Kusche-Vihrog, K. Nanomechanics of the endothelial glycocalyx contribute to Na+-induced vascular inflammation. Sci. Rep. 2017, 7, 46476.
- Watso, J.C.; Farquhar, W.B. Hydration Status and Cardiovascular Function. Nutrients 2019, 11, 1866.
- Hao, C.M.; Yull, F.; Blackwell, T.; Kömhoff, M.; Davis, L.S.; Breyer, M.D. Dehydration activates an NF-kappaB-driven, COX2-dependent survival mechanism in renal medullary interstitial cells. J. Clin. Investig. 2000, 106, 973–982.
- Ishihara, S.; Yasuda, M.; Harada, I.; Mizutani, T.; Kawabata, K.; Haga, H. Substrate stiffness regulates temporary NF-κB activation via actomyosin contractions. Exp. Cell Res. 2013, 319, 2916–2927.
- Fuller, N.; Rand, R.P. Water in actin polymerization. Biophys. J. 1999, 76, 3261–3266.
- Laudadio, R.E.; Millet, E.J.; Fabry, B.; An, S.S.; Butler, J.P.; Fredberg, J.J. Rat airway smooth muscle cell during actin modulation: Rheology and glassy dynamics. Am. J. Physiol. Cell Physiol. 2005, 289, C1388–C1395.
- Liu, F.; Mih, J.D.; Shea, B.S.; Kho, A.T.; Sharif, A.S.; Tager, A.M.; Tschumperlin, D.J. Feedback amplification of fibrosis through matrix stiffening and cox-2 suppression. J. Cell Biol. 2010, 190, 693–706.
- Huang, X.; Yang, N.; Fiore, V.F.; Barker, T.H.; Sun, Y.; Morris, S.W.; Ding, Q.; Victor, J.; Thannickal, V.J.; Zhou, Y. Matrix stiffness–induced myofibroblast differentiation is mediated by intrinsic mechanotransduction. Am. J. Respir. Cell Mol. Biol. 2012, 47, 340–348.
- Uehata, M.; Ishizaki, T.; Satoh, H.; Ono, T.; Kawahara, T.; Morishita, T.; Tamakawa, H.; Yamagami, K.; Inui, J.; Maekawa, M.; et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature 1997, 389, 990–994.
- Li, R.; Song, X.; Li, G.; Hu, Z.; Sun, L.; Chen, C.; Yang, L. Ibuprofen attenuates interleukin-1β-induced inflammation and actin reorganization via modulation of RhoA signaling in rabbit chondrocytes. Acta Biochim. Biophys. Sin. (Shanghai) 2019, 51, 1026–1033.
- Day, M. Covid-19: Ibuprofen should not be used for managing symptoms, say doctors and scientists. BMJ 2020, 368, 1086.
- Olson, M.F. Applications for ROCK kinase inhibition. Curr. Opin. Cell Biol. 2008, 20, 242–248.
- Mulherkar, S.; Tolias, K.F. RhoA-ROCK signaling as a therapeutic target in traumatic brain injury. Cells 2020, 9, 245.
- Strassheim, D.; Gerasimovskaya, E.; Irwin, D.; Dempsey, E.C.; Stenmark, K.; Karoor, V. RhoGTPase in Vascular Disease. Cells 2019, 8, 551.
- Zhou, N.; Lee, J.J.; Stoll, S.; Ma, B.; Costa, K.D.; Qiu, H. Rho Kinase Regulates Aortic Vascular Smooth Muscle Cell Stiffness Via Actin/SRF/Myocardin in Hypertension. Cell Physiol. Biochem. 2017, 44, 701–715.
- Møller, L.L.V.; Klip, A.; Sylow, L. Rho GTPases—Emerging Regulators of Glucose Homeostasis and Metabolic Health. Cells 2019, 8, 434.
- Abedi, F.; Hayes, A.W.; Reiter, R.; Karimi, G. Acute lung injury: The therapeutic role of Rho kinase inhibitors. Pharm. Res. 2020, 155, 104736.
- Du, L.; Zhou, J.; Zhang, J.; Yan, M.; Gong, L.; Liu, X.; Chen, M.; Tao, K.; Luo, N.; Liu, J. Actin filament reorganization is a key step in lung inflammation induced by systemic inflammatory response syndrome. Am. J. Respir. Cell Mol. Biol. 2012, 47, 597–603.
- Aroor, A.R.; Jia, G.; Sowers, J.R. Cellular mechanisms underlying obesity-induced arterial stiffness. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R387–R398.


