Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Edward N. Harris | + 2669 word(s) | 2669 | 2021-07-15 10:34:08 | | | |
| 2 | Vivi Li | Meta information modification | 2669 | 2021-07-28 03:42:44 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Harris, E.N. HARE/Stabilin-2. Encyclopedia. Available online: https://encyclopedia.pub/entry/12504 (accessed on 27 July 2026).
Harris EN. HARE/Stabilin-2. Encyclopedia. Available at: https://encyclopedia.pub/entry/12504. Accessed July 27, 2026.
Harris, Edward N.. "HARE/Stabilin-2" Encyclopedia, https://encyclopedia.pub/entry/12504 (accessed July 27, 2026).
Harris, E.N. (2021, July 27). HARE/Stabilin-2. In Encyclopedia. https://encyclopedia.pub/entry/12504
Harris, Edward N.. "HARE/Stabilin-2." Encyclopedia. Web. 27 July, 2021.
Copy Citation
The Stabilin receptors are a two-member family in the type H class of scavenger receptors. These dynamic receptors bind and internalize multiple ligands from the cell surface for the purpose of clearing extracellular material including some synthetic drugs and for sensing the external environment of the cell. Stabilin-1 was the first receptor to be cloned, though the biological activity of Hyaluronic Acid Receptor for Endocytosis (HARE)/Stabilin-2 was observed about 10 years prior to the cloning of Stabilin-1. Stabilin-1 has a more diverse expression profile among the tissues than HARE/Stabilin-2.
stabilin
HARE
receptor-mediated endocytosis
scavenger receptor
hyaluronan
1. Initial Discovery
The discovery of the cloned Stabilin receptors began with Stabilin-1 in the early 1990s. A monoclonal antibody raised against whole human spleen homogenate produced a monoclonal antibody (MS-1) against an antigen that was initially identified as a large protein that is preferentially expressed in the sinusoidal endothelia of spleen, lymph node, liver, and adrenal cortex [1]. The identified MS-1 antigen is an inducible protein not found in normal skin lesions, but rather in diseased skin and some cancers such as psoriasis, melanoma, and T-cell lymphomas. Induction of MS-1 antigen is stimulated by Interleukin (IL)-4 and Dexamethasone and inhibited by tumor necrosis factor (TNF)-α, IL-1β, and IL-6 [2] in cultured human peripheral blood monocytes [3][4]. The use of the MS-1 antibody allowed for immunoprecipitation of the MS-1 antigen which was subjected to matrix-assisted laser desorption ionization—time of flight (MALDI-TOF) analysis for protein identification. The same authors used hyaluronic acid (HA) affinity chromatography to identify a similar protein that bound HA with a high degree of topology between both receptors [3]. However, several years prior to the publication of Politz et al. [3], the rat HA-binding receptor was discovered from rat liver sinusoidal endothelial cells (SEC HA receptor) [5][6] and identified as the primary HA turnover receptor named HARE (Hyaluronic Acid Receptor for Endocytosis) [7]. Due to their topological similarity, the receptors that were formerly named MS-1 antigen or protein, FEEL-1, CLEVER-1 and FEEL-2, HARE are now almost universally recognized as Stabilin-1 and Stabilin-2, respectively [3][8][9] (Figure 1 as annotated by [10][11]). Although there was a bit of confusion as to the number of protein subunits composing these receptors during their initial characterization [1][7][12], cloning and expression in stable cell lines have confirmed that Stabilin-1 is a monomer with two forms expressed as a tight doublet at about 320 kDa by sodium dodecyl sulfate—polyacrylamide gel electrophoresis (SDS-PAGE) [13] and a possible smaller isoform at around 140 kDa by SDS-PAGE which has not been confirmed by other sources [14]. Human Stabilin-2 is expressed as two monomers or isoforms at 315-kDa and 190-kDa by SDS-PAGE [15] (Figure 2) which is similar to rat Stabilin-2 expressed as 300-kDa and 175-kDa isoforms [7]. Both human and rat receptors have nearly the same number of amino acids but the higher mass of the human receptor is solely contributed by post-translational glycosylation of the receptor. In native SECs, the ratio of the large to small isoforms is close to a 1:1 ratio in liver, whereas in the spleen, it is 2:1 or 3:1 [7]. However, in stable cell lines, the isoform ratio is closer to 8–10:1, suggesting that the mechanism for the appearance of the 190HARE is more robust in primary cells [15]. It is thought that these isoforms are a product of proteolytic cleavage that occurs somewhere between the trans-Golgi network and the plasma membrane during synthesis. Using [35S] labeled tracking, the appearance of the small isoform appeared immediately after the large isoform was fully glycosylated (unpublished data). For simplicity in nomenclature, the large isoform of Stabilin-2 will be referred to as Stabilin-2 and the smaller isoform of the same receptor will be referred to as the 190HARE.
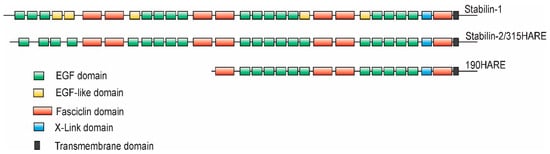
Figure 1. Topology of Stabilin-1 and Stabilin-2. The overall topologies of human Stabilin-1 and human Stabilin-2, including the 190HARE, are very similar with an overall protein sequence identity of 39% and similarity of 56% between the two large receptors. The topology algorithm was derived using the SMART program and the homology was calculated using BLAST program of the National Center for Biotechnology Information.
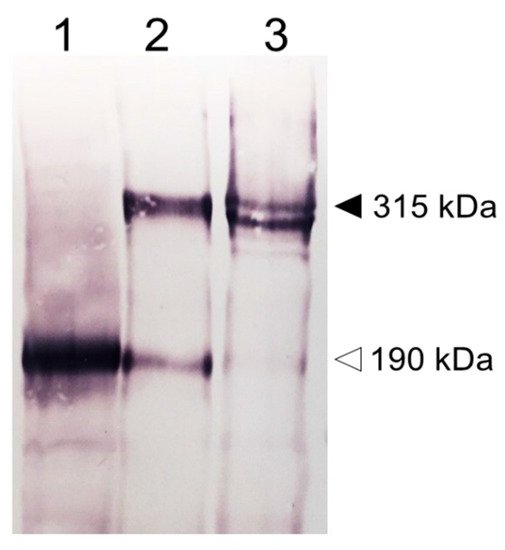
Figure 2. Protein characterization by SDS-PAGE. Stable cell lines expressing human (1) 190HARE, (2) Stabilin-2 or 315HARE, and (3) Stabilin-1 were prepared as whole cell lysates and 20 μg of lysates were separated by 5% denaturing gel electrophoresis without reduction. The dark arrowhead (315 kDa) indicates the monomer of Stabilin-2 and doublet of Stabilin-1 and the open arrowhead (190 kDa) indicates the presence of the 190HARE in cell lysates expressing a construct that only codes for the 190HARE and Stabilin-2. All plasmid constructs (pcDNA5/FRT/V5-6xHis, Invitrogen, Carlsbad, CA) used to express these proteins contained the cytomegalovirus promoter for protein expression and C-terminal tags, 6xHis and V5. Anti-V5 antibody was used to detect the recombinant protein followed by a secondary antibody for standard colorimetric detection with alkaline phosphatase.
2. Hyaluronan Binding and Endocytosis
Hyaluronan (HA) is a very long polymer containing disaccharides composed of glucuronic acid β1-3 linked to N-acetylglucosamine and these disaccharides are linked by β1-4 glycosidic bonds. HA is a necessary extracellular matrix molecule essential for a wide variety of functions including joint health and lubrication, ocular integrity, skin hydration, fertility, cancer metastasis, and many more physiological functions [16][17][18][19][20]. Likewise, turnover of HA is very dynamic in which hyaluronidases/spreading factor/mechanical forces/oxidative stress break down very large HA in the tissues which then drains into the lymphatics and finally into the blood [21]. It is the function of clearance receptors in the lymphatics and circulatory systems to maintain low levels of HA in these fluids. Early studies of HA metabolism reveal that [3H]Hyaluronan injected via the marginal ear vein of a mature rabbit is rapidly cleared in about 20 min from plasma with a half-life of 3.5–6 min [22]. Tissue accumulation of [3H]Hyaluronan was primarily in liver, spleen, lymph nodes, and bone marrow, all tissues containing high expression levels of Stabilin-2 with large overlaps in Stabilin-1 expression [23]. Furthermore, the liver SECs, and not the Kupffer or hepatocytes, have the highest accumulation of hyaluronan [24]. Both Stabilin-1 and Stabilin-2 are highly expressed in liver SECs among other tissues previously mentioned. Although both Stabilins contain X-Link domains, the Stabilin-1 domain is inactive for hyaluronan binding [3], thus Stabilin-2 is the primary scavenger receptor for systemic HA.
To investigate the HA–Stabilin-2 interaction more closely and in detail, we expressed the human 190HARE (smaller isoform) in Flp-In HEK 293 cells [25]. Using a variety of techniques including HA degradation monitoring, Scatchard analysis, and internalization vs binding assays, the disassociation rate (kD) for the 190HARE was calculated to be 7.2 nM which was similar to the kD in recombinant SK-Hep-1 cells expressing the rat 175HARE at 4.1 nM [26]. In addition, the 190HARE recycling time on the cell surface was 7–9 min which was also similar to what had been calculated for recycling time for native rat Stabilin-2 receptors in primary liver SECs [27]. These physical properties revealed that the recombinant 190HARE was a good model for the native Stabilin-2 receptors in SECs, a cell type that is difficult to culture and maintain its differentiation for more than two days. Full-length Stabilin-2 was also expressed in stable cell lines under the same promoter and cell type as the 190HARE. For reasons unknown, the expression level of Stabilin-2 was always robust, but significantly lower than the 190HARE. In addition, the 190HARE was always co-expressed with Stabilin-2 in these cell lines, suggesting that production of the small isoform is a natural byproduct of the full-length receptor. Endocytosis of HA in the Stabilin-2 cells was not significantly different from the 190HARE when accounting for the number of cells and receptor expression levels. Expressing both isoforms as soluble ecto-domains and purifying them by nickel chelate chromatography revealed that the binding of HA in an ELISA-like format for both receptors was nearly equivalent (20.7 ± 6.4 nM for 190-HARE, 9.9 ± 1.2 nM for Stabilin-2) [15], further demonstrating that the binding occurs primarily within the X-Link domain and not in other areas of the receptor. X-Link binding of HA was further confirmed in cells expressing a receptor without the X-Link domain as these cells were not able to internalize any [125]I-HA [28]. Curiously, expression of the Stabilin-2 ecto-domain did not result in producing a small isoform, indicating that membrane attachment is required for small isoform (190HARE) expression (Figure 3).
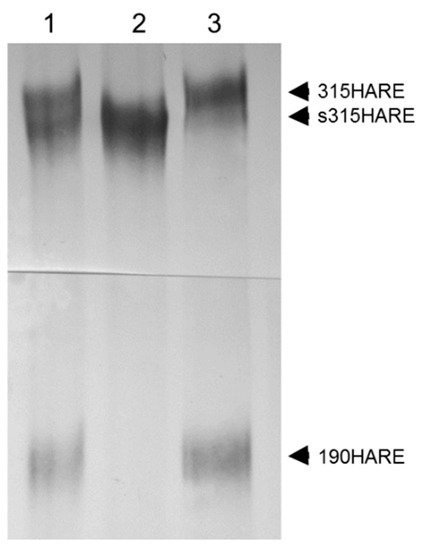
Figure 3. Membrane attachment is required for production of 190HARE. Cells expressing either 315HARE (Stabilin-2) or secreted 315HARE (lacking the transmembrane and cytoplasmic domains) were lysed in conditioned medium and immunoprecipitated with Ab30 conjugated to sepharose beads. Immunoprecipitated proteins from (1) cell lysate from both 315HARE and s315HARE were mixed 1:1, (2) s315HARE lyste, and (3) 315HARE lysate were separated by 5% SDS-PAGE on a 20 cm gel, blotted, and probed with anti-V5 antibody for protein detection.
The X-Link domain of Stabilin-2 has the highest homology to another well-known hyalectin, tumor necrosis factor inducible gene 6 protein (TSG-6), a protein whose expression is induced by inflammatory mediators, growth factors, ovulation inducers, and matrix remodeling [29][30][31]. In silico modeling based on the structural work performed with TSG-6 reveals that HA binds within a groove of the X-Link domain and interacts directly with tyrosine residues through hydrogen bonding [32]. This TSG-6 model has been upgraded more recently to reflect the more intimate interaction of HA with the X-Link domain in which binding is mediated with a combination of ring stacking interactions, hydrogen bonds, salt bridges, and hydrophobic interactions along the binding groove [33]. The similarity of HA binding with TSG-6 and the 190HARE was further corroborated by another independent study using atomic force microscopy (AFM) to determine the bond strength between HA and several hyalectins. Using biotinylated HA at the tip of the AFM probe and proteins bound to solid substratum, the mean rupture force could be calculated between a single polymer of HA and the hyalectin. Both TSG-6 and 190HARE had a mean rupture force with HA at 24 and 25 picoNewtons (pN), respectively, further suggesting that both proteins interact with HA in a similar fashion. Comparatively, HA rupture forces were 34 pN for CD44, 37 pN for versican, and more than 52 pN for aggrecan X-Link domains [34]. Although the binding affinity of HA for 190HARE is in the low nanomolar range, the bond strength with which it binds is one of the weakest among the hyalectin family. Biologically, this is logical as this is a constitutively recycling receptor that must bind HA with high specificity and then release this cargo in the endosomes. The other hyalectins with higher rupture forces are all HA-binding proteins for structural purposes, hence the more stable interactions.
Data derived from cell models suggests that the degradation of internalized HA does occur, though this is highly dependent on the cell type. For example, SK-Hep-1 cells are of endothelial origin [35], but are very active in transcytosis. It is estimated that about half of the HA that is internalized by SK-Hep-1 cells stably expressing the rat 175HARE is delivered back to the extracellular space entirely intact [36]. In 190HARE cells (HEK293 background), most of the HA is degraded and the ratio of intact versus degraded HA may be measured by cetylpryridinium chloride precipitation [36][37]. Primary SECs have a much greater capacity for endocytosis and degradation of HA than these stable recombinant cell models. HA is internalized by Stabilin-2/HARE and transported to lysosomes which contain lysosomal β-glucuronidases, β-N-acetylglucoaminidase, and hyaluronidases for polymer degradation. The HA polymer is degraded into monosaccharides that are transferred to the cytosol where glucuronic acid is converted by isomerases for metabolism. The N-acetylglucosamine is also transferred to the cytoplasm and phosphorylated then deacetylated for the eventual formation of acetate, lactate, and fructose [37][38][39].
3. Heparin Clearance by Stabilin-1 and Stabilin-2
Heparin is a complex polysaccharide made up of two base sugars, N-acetylglucosamine and glucuronic (epimer: iduronic) acid. These two sugars may be modified in the Golgi of the cell to give rise to up to 32 different disaccharide combinations resulting in more than 100 trillion unique heparin polymers [40]. The heparin core protein is Serglycin which is enriched in serine and glycine residues in which the serine is modified with the heparin polymer. As a post-translational product of Serglycin, the first modifications occur in the ER and a step-wise process of elongation and modification to the sugar chain occurs in the Golgi [41]. Heparin is produced in mast cells of the immune system which are enriched in the gut mucosa of mammals [42]. Heparin was discovered in 1916 by a medical student, Jay McLean, working in the laboratory of Professor Howell, and its use as an anticoagulant in medical applications began in the mid-1930s [43]. Three forms of heparin for use in medicine are: purified unfractionated heparin (UFH) with a mass average of 13.5 kDa, the processed UFH in the development of low-molecular-weight heparin (LMWH) with a mass average of 3.5 kDa, and the synthetic pentasaccharide, Fondaparinux [44].
Regardless of the route of administration, heparin clearance from circulation is dependent on the liver and kidneys. Longer polymers tend to be taken up by the liver for degradation and then undergo filtration by the kidney. It was discovered that the SECs of the liver are the primary site for heparin clearance from blood [45][46]. The primary, though not only, receptor for heparin clearance was discovered to be Stabilin-2 using direct binding methods of purified protein with radioactively labeled heparin [47]. Heparin has a high electronegative charge and, thus, it is a “sticky” molecule that binds to many components on the surface of cells. Cells that do not express Stabilin-2 will slowly internalize heparin during the process of normal membrane turnover. However, cells that do express either Stabilin-2 or 190HARE take up much more heparin compared to the same type of cells that do not express these receptors. Both UFH and LMWH are taken up by Stabilin-2 [44] and Stabilin-1 is also involved with heparin binding and endocytosis [48].
As previously mentioned, heparin is a very heterogeneous molecule with respect to both size and modification. Currently, therapeutic heparin is a natural product that comes from swine gut mucosa from which it is purified and refined [49][50]. The goal of making synthetic heparin has been around for decades [51][52] and is currently becoming a reality with the development and refinement of chemoenzymatic processes [53][54][55][56][57]. Using homogenous synthetic heparin polymers, we have found that the heparin polymer needs to be sulfated for binding with Stabilin-1 or Stabilin-2. The nonsulfated polymer, heparosan, does not interact with the Stabilin clearance receptors and may be a good candidate as a vehicle for drug delivery [58][59]. The minimal binding requirement is for the polymer to be at least 10 sugars long and the presence of the 3-O sulfate increases binding affinity at least fivefold [48]. To determine how important the 3-O sulfate is for overall metabolism and uptake in the liver, we subcutaneously injected mice with physiological doses of dodecamers (12mers) with or without a single 3-O sulfation residue (15 vs. 14 total sulfate groups along a single dodecamer). The 3-O sulfate routed the polymers through the liver for endocytosis and degradation and also increased the half-life of the 12mer in blood. Without the 3-O sulfation, the polymer had a far higher chance of getting immediately filtered by the kidneys [60]. From these experiments, we deducted that polymer charge is not the only defining characteristic of heparin–Stabilin interactions. The orientation of the charge is just as important as the charge itself for optimal binding with the receptor. The single 3-O modification along the polymer affected affinity to Stabilin-2 and liver, increased the half-life in blood, and, possibly, allowed interaction of heparin with other blood components. It is clear that the Stabilin-2 receptor directly affects the half-life of systemic UFH and LMWH.
The precise location of where heparin binds with the Stabilin-2 receptor is not known and the primary sequence of the amino acids does not reveal an obvious cluster of positively charged amino acids that are common for many heparin binding proteins. Since both Stabilin-2 and 190HARE bind with heparin with the same affinity [47], the putative binding site(s) are only in the 190HARE sequence and the N-terminal half of Stabilin-2 does not contribute to binding, despite similar domain organization (Figure 1). Stabilin-2 (and Stabilin-1) are both extensively folded and it is hypothesized that a heparin binding domain/groove is formed as distal amino acids are brought together in close proximity to form this site. Unfolding of the receptor through reduction of the disulfide bonds renders the receptor unable to bind any ligands.
References
- Goerdt, S. Identification of a novel high molecular weight protein preferentially expressed by sinusoidal endothelial cells in normal human tissues. J. Cell Biol. 1991, 113, 1425–1437.
- Goerdt, S.; Bhardwaj, R.; Sorg, C. Inducible expression of MS-1 high-molecular-weight protein by endothelial cells of continuous origin and by dendritic cells/macrophages in vivo and in vitro. Am. J. Pathol. 1993, 142, 1409–1422.
- Politz, O.; Gratchev, A.; McCourt, P.A.G.; Schledzewski, K.; Guillot, P.; Johansson, S.; Svineng, G.; Franke, P.; Kannicht, C.; Kzhyshkowska, J.; et al. Stabilin-1 and -2 constitute a novel family of fasciclin-like hyaluronan receptor homologues. Biochem. J. 2002, 362 Pt 1, 155–164.
- Blume-Peytavi, U.; Adler, Y.D.; Geilen, C.C.; Ahmad, W.; Christiano, A.; Goerdt, S.; Orfanos, C.E. Multiple familial cutaneous glomangioma: A pedigree of 4 generations and critical analysis of histologic and genetic differences of glomus tumors. J. Am. Acad. Dermatol. 2000, 42, 633–639.
- Yannariello-Brown, J.; Zhou, B.; Ritchie, D.; Oka, J.A.; Weigel, P.H. A novel ligand blot assay detects different hyaluronan-binding proteins in rat liver hepatocytes and sinusoidal endothelial cells. Biochem. Biophys. Res. Commun. 1996, 218, 314–319.
- Yannariello-Brown, J.; Zhou, B.; Weigel, P.H. Identification of a 175 kDa protein as the ligand-binding subunit of the rat liver sinusoidal endothelial cell hyaluronan receptor. Glycobiology 1997, 7, 15–21.
- Zhou, B.; Weigel, J.A.; Fauss, L.; Weigel, P.H. Identification of the hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2000, 275, 37733–37741.
- Tamura, Y.; Adachi, H.; Osuga, J.I.; Ohashi, K.; Yahagi, N.; Sekiya, M.; Okazaki, H.; Tomita, S.; Iizuka, Y.; Shimano, H. FEEL-1 and FEEL-2 are endocytic receptors for advanced glycation end products. J. Biol. Chem. 2003, 278, 12613–12617.
- Irjala, H.; Alanen, K.; Grénman, R.; Heikkilä, P.; Joensuu, H.; Jalkanen, S. Mannose receptor (MR) and common lymphatic endothelial and vascular endothelial receptor (CLEVER)-1 direct the binding of cancer cells to the lymph vessel endothelium. Cancer Res. 2003, 63, 4671–4676.
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, 43, D257–D260.
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496.
- Zhou, B.; Oka, J.A.; Singh, A.; Weigel, P.H. Purification and subunit characterization of the rat liver endocytic hyaluronan receptor. J. Biol. Chem. 1999, 274, 33831–33834.
- Kzhyshkowska, J.; Gratchev, A.; Martens, J.-H.; Pervushina, O.; Mamidi, S.; Johansson, S.; Schledzewski, K.; Hansen, B.; He, X.; Tang, J.; et al. Stabilin-1 localizes to endosomes and the trans-Golgi network in human macrophages and interacts with GGA adaptors. J. Leukoc. Biol. 2004, 76, 1151–1161.
- Prevo, R.; Banerji, S.; Ni, J.; Jackson, D.G. Rapid plasma membrane-endosomal trafficking of the lymph node sinus and high endothelial venule scavenger receptor/homing receptor stabilin-1 (FEEL-1/CLEVER-1). J. Biol. Chem. 2004, 279, 52580–52592.
- Harris, E.N.; Kyosseva, S.V.; Weigel, J.A.; Weigel, P.H. Expression, processing, and glycosaminoglycan binding activity of the recombinant human 315-kDa hyaluronic acid receptor for endocytosis (HARE). J. Biol. Chem. 2007, 282, 2785–2797.
- Rodriguez-Martinez, H.; Tienthai, P.; Atikuzzaman, M.; Carrillo, A.V.; Rubér, M.; Alvarez-Rodriguez, M. The ubiquitous hyaluronan: Functionally implicated in the oviduct? Theriogenology 2016, 86, 182–186.
- Euppayo, T.; Punyapornwithaya, V.; Chomdej, S.; Ongchai, S.; Nganvongpanit, K. Effects of hyaluronic acid combined with anti-inflammatory drugs compared with hyaluronic acid alone, in clinical trials and experiments in osteoarthritis: A systematic review and meta-analysis. BMC Musculoskelet. Disord. 2017, 18, 387.
- Balazs, E.A. Fine structure and function of ocular tissues. Int. Ophthalmol. Clin. 1973, 13, 169–187.
- Robert, L. Hyaluronan, a truly “youthful” polysaccharide. Its Med Appl. Pathol Biol (Paris) 2015, 63, 32–34.
- McCarthy, J.B.; El-Ashry, D.; Turley, E.A. Hyaluronan, Cancer-Associated Fibroblasts and the Tumor Microenvironment in Malignant Progression. Front. Cell Dev. Biol. 2018, 6, 48.
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33.
- Fraser, J.R.F.; Laurent, T.C.; Pertoft, H.; Baxter, E. Plasma clearance, tissue distribution and metabolism of hyaluronic acid injected intravenously in the rabbit. Biochem. J. 1981, 200, 415–424.
- Fraser, J.R.; Appelgren, L.E.; Laurent, T.C. Tissue uptake of circulating hyaluronic acid. A whole body autoradiographic study. Cell Tissue Res. 1983, 233, 285–293.
- Eriksson, S.; Fraser, J.E.; Laurent, T.C.; Pertoft, H.; Smedsrød, B. Endothelial cells are a site of uptake and degradation of hyaluronic acid in the liver. Exp. Cell Res. 1983, 144, 223–228.
- Harris, E.N.; Weigel, J.A.; Weigel, P.H. Endocytic function, glycosaminoglycan specificity, and antibody sensitivity of the recombinant human 190-kDa hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2004, 279, 36201–36209.
- Zhou, B.; Weigel, J.A.; Saxena, A.; Weigel, P.H. Molecular cloning and functional expression of the rat 175-kDa hyaluronan receptor for endocytosis. Mol. Biol. Cell 2002, 13, 2853–2868.
- McGary, C.T.; Raja, R.H.; Weigel, P.H. Endocytosis of hyaluronic acid by rat liver endothelial cells. Evidence for receptor recycling. Biochem. J. 1989, 257, 875–884.
- Kyosseva, S.V.; Harris, E.N.; Weigel, P.H. The hyaluronan receptor for endocytosis mediates hyaluronan-dependent signal transduction via extracellular signal-regulated kinases. J. Biol. Chem. 2008, 283, 15047–15055.
- Bayliss, M.; Howat, S.; Dudhia, J.; Murphy, J.; Barry, F.; Edwards, J.; Day, A. Up-regulation and differential expression of the hyaluronan-binding protein TSG-6 in cartilage and synovium in rheumatoid arthritis and osteoarthritis. Osteoarthr. Cartil. 2001, 9, 42–48.
- Glant, T.T.; Kamath, R.V.; Bardos, T.; Gal, I.; Szanto, S.; Murad, Y.M.; Sandy, J.D.; Mort, J.S.; Roughley, P.J.; Mikecz, K. Cartilage-specific constitutive expression of TSG-6 protein (product of tumor necrosis factor alpha-stimulated gene 6) provides a chondroprotective, but not antiinflammatory, effect in antigen-induced arthritis. Arthritis Rheum. 2002, 46, 2207–2218.
- Mukhopadhyay, D.; Mukhopadhyay, D.; Hascall, V.C.; Day, A.J.; Salustri, A.; Fulop, C. Two distinct populations of tumor necrosis factor-stimulated gene-6 protein in the extracellular matrix of expanded mouse cumulus cell-oocyte complexes. Arch. Biochem. Biophys. 2001, 394, 173–181.
- Blundell, C.D.; Almond, A.; Mahoney, D.J.; DeAngelis, P.L.; Campbell, I.D.; Day, A.J. Towards a structure for a TSG-6.hyaluronan complex by modeling and NMR spectroscopy: Insights into other members of the link module superfamily. J. Biol. Chem. 2005, 280, 18189–18201.
- Higman, V.A.; Briggs, D.C.; Mahoney, D.J.; Blundell, C.D.; Sattelle, B.M.; Dyer, D.P.; Green, D.E.; DeAngelis, P.L.; Almond, A.; Milner, C.M.; et al. A refined model for the TSG-6 link module in complex with hyaluronan: Use of defined oligosaccharides to probe structure and function. J. Biol. Chem. 2014, 289, 5619–5634.
- Bano, F.; Tammi, M.I.; Kang, D.W.; Harris, E.N.; Richter, R.P. Single-Molecule Unbinding Forces between the Polysaccharide Hyaluronan and Its Binding Proteins. Biophys. J. 2018, 114, 2910–2922.
- Heffelfinger, S.C.; Hawkins, H.H.; Barrish, J.; Taylor, L.; Darlington, G.J. SK HEP-1: A human cell line of endothelial origin. Vitr. Cell. Dev. Biol. 1992, 28, 136–142.
- Weigel, J.A.; Weigel, P.H. Characterization of the recombinant rat 175-kDa hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2003, 278, 42802–42811.
- McGary, C.T.; Yannariello-Brown, J.; Kim, D.W.; Stinson, T.C.; Weigel, P.H. Degradation and intracellular accumulation of a residualizing hyaluronan derivative by liver endothelial cells. Hepatology 1993, 18, 1465–1476.
- Smedsrod, B.; Pertoft, H.; Eriksson, S.; Fraser, J.R.; Laurent, T.C. Studies in vitro on the uptake and degradation of sodium hyaluronate in rat liver endothelial cells. Biochem. J. 1984, 223, 617–626.
- Smedsrod, B. Cellular Events in the Uptake and Degradation of Hyaluronan. Adv. Drug Deliv. Rev. 1991, 7, 265–278.
- Laremore, T.N.; Zhang, F.; Dordick, J.S.; Liu, J.; Linhardt, R.J. Recent progress and applications in glycosaminoglycan and heparin research. Curr. Opin. Chem. Biol. 2009, 13, 633–640.
- Rabenstein, D.L. Heparin and heparan sulfate: Structure and function. Nat. Prod. Rep. 2002, 19, 312–331.
- Kwaan, H.C.; Hatem, A. Effect of lung and gut heparin on experimental arterial thrombosis. Adv. Exp. Med. Biol. 1975, 52, 263–268.
- Barrowcliffe, T.W. History of heparin. Handb. Exp. Pharmacol. 2012, 207, 3–22.
- Harris, E.N.; Baggenstoss, B.A.; Weigel, P.H. Rat and human HARE/stabilin-2 are clearance receptors for high- and low-molecular-weight heparins. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1191–G1199.
- Praaning, D.P.P.-V.; Brouwer, A.; Knook, D.L. Clearance capacity of rat liver Kupffer, Endothelial, and parenchymal cells. Gastroenterology 1981, 81, 1036–1044.
- Oie, C.I.; Olsen, R.; Smedsrod, B.; Hansen, J.B. Liver sinusoidal endothelial cells are the principal site for elimination of unfractionated heparin from the circulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G520–G528.
- Harris, E.N.; Weigel, J.A.; Weigel, P.H. The human hyaluronan receptor for endocytosis (HARE/Stabilin-2) is a systemic clearance receptor for heparin. J. Biol. Chem. 2008, 283, 17341–17350.
- Pempe, E.H.; Xu, Y.; Gopalakrishnan, S.; Liu, J.; Harris, E.N. Probing structural selectivity of synthetic heparin binding to stabilin protein receptors. J. Biol. Chem. 2012, 287, 20774–20783.
- Mulloy, B.; Heath, A.; Shriver, Z.; Jameison, F.; Al Hakim, A.; Morris, T.S.; Szajek, A.Y. USP compendial methods for analysis of heparin: Chromatographic determination of molecular weight distributions for heparin sodium. Anal. Bioanal. Chem. 2014, 406, 4815–4823.
- Volpi, N.; Maccari, F.; Suwan, J.; Linhardt, R.J. Electrophoresis for the analysis of heparin purity and quality. Electrophoresis 2012, 33, 1531–1537.
- Petitou, M. Chemical Synthesis of Heparin. In Heparin: Chemical and Biological Properties, Clinical Applications; Lane, D.A., Lindahl, U., Eds.; CRC Press: Boca Raton, FL, USA, 1989; pp. 65–79.
- Lindahl, U. Approaches to the synthesis of heparin. Haemostasis 1990, 20 (Suppl. 1), 146–153.
- Liu, R.; Xu, Y.; Chen, M.; Weïwer, M.; Zhou, X.; Bridges, A.S.; DeAngelis, P.L.; Zhang, Q.; Linhardt, R.J.; Liu, J. Chemoenzymatic design of heparan sulfate oligosaccharides. J. Biol. Chem. 2010, 285, 34240–34249.
- Linhardt, R.J.; Liu, J. Synthetic heparin. Curr. Opin. Pharmacol. 2012, 12, 217–219.
- Xu, Y.; Cai, C.; Chandarajoti, K.; Hsieh, P.H.; Li, L.; Pham, T.Q.; Sparkenbaugh, E.M.; Sheng, J.; Key, N.S.; Pawlinski, R.; et al. Homogeneous low-molecular-weight heparins with reversible anticoagulant activity. Nat. Chem. Biol. 2014, 10, 248–250.
- Bhaskar, U.; Li, G.; Fu, L.; Onishi, A.; Suflita, M.; Dordick, J.S.; Linhardt, R.J. Combinatorial one-pot chemoenzymatic synthesis of heparin. Carbohydr. Polym. 2015, 122, 399–407.
- Joice, A.; Raman, K.; Mencio, C.; Quintero, M.V.; Brown, S.; Nguyen, T.K.; Kuberan, B. Enzymatic synthesis of heparan sulfate and heparin. Methods Mol. Biol. 2015, 1229, 11–19.
- Lane, R.S.; Haller, F.M.; Chavaroche, A.A.E.; Almond, A.; DeAngelis, P.L. Heparosan-coated liposomes for drug delivery. Glycobiology 2017, 27, 1062–1074.
- DeAngelis, P.L. Heparosan, a promising ‘naturally good’ polymeric conjugating vehicle for delivery of injectable therapeutics. Expert Opin. Drug Deliv. 2015, 12, 349–352.
- Miller, C.M.; Xu, Y.; Kudrna, K.M.; Hass, B.E.; Kellar, B.M.; Egger, A.W.; Liu, J.; Harris, E.N. 3-O sulfation of heparin leads to hepatotropism and longer circulatory half-life. Thromb. Res. 2018, 167, 80–87.
More
Information
Subjects:
Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
987
Revisions:
2 times
(View History)
Update Date:
28 Jul 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No