 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Khojasteh Malekmohammad | + 6117 word(s) | 6117 | 2021-07-15 10:50:36 | | | |
| 2 | Vivi Li | -3041 word(s) | 3076 | 2021-07-23 04:27:35 | | | | |
| 3 | Vivi Li | Meta information modification | 3076 | 2021-07-23 04:51:37 | | |
Video Upload Options
Atherosclerosis is a chronic inflammatory disease which is a major cause of coronary heart disease and stroke in humans. It is characterized by intimal plaques and cholesterol accumulation in arterial walls. The side effects of currently prescribed synthetic drugs and their high cost in the treatment of atherosclerosis has prompted the use of alternative herbal medicines, dietary supplements, and antioxidants associated with fewer adverse effects for the treatment of atherosclerosis. Natural and synthetic antioxidants have a crucial role in the prevention and treatment of atherosclerosis through different mechanisms. These include: The inhibition of low density lipoprotein (LDL) oxidation, the reduction of reactive oxygen species (ROS) generation, the inhibition of cytokine secretion, the prevention of atherosclerotic plaque formation and platelet aggregation, the preclusion of mononuclear cell infiltration, the improvement of endothelial dysfunction and vasodilation, the augmentation of nitric oxide (NO) bioavailability, the modulation of the expression of adhesion molecules such as vascular cell adhesion molecule-1 (VCAM-1) and intercellular adhesion molecule-1 (ICAM-1) on endothelial cells, and the suppression of foam cell formation.
1. Introduction
2. Oxidative Stress and the Atherosclerotic Process
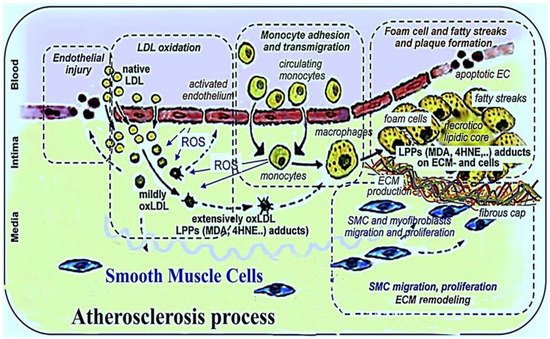
3. Endogenous Enzymatic Antioxidants
3.1. Superoxide Dismutase (SOD)
3.2. Catalase
3.3. Glutathione Peroxidase (GPx)
3.4. Thioredoxin Reductase
4. Endogenous Non-Enzymatic Antioxidants
4.1. Glutathione
4.2. Coenzyme Q (CoQ)
4.3. Bilirubin
4.4. Uric Acid
4.5. Lipoic Acid
5. Exogenous Non-Enzymatic Antioxidants (Natural Antioxidants)
5.1. Vitamin EVitamin E is the most comprehensively studied lipid soluble antioxidant in humans. It consists of eight isomeric compounds (α-, β-, γ-, and δ- tocopherol; and α-, β-, γ-, and δ-tocotrienol) [9][66]. Cooking oils, egg yolk, butter, green leafy vegetables, and some fruit (kiwi fruit, pumpkins, mangoes, papayas, and tomatoes) are rich sources of vitamin E [9].
In several animal models, vitamin E has preventative effects against atherosclerosis by: Scavenging free radicals in VSMC, diminishing the oxidation of LDL by the inhibition of Cluster Differentiating 36 (CD36) and Scavenger receptor class B type I (SR-BI) expression in VSMC, reducing VSMC proliferation via the inhibition of protein kinase C (PKC), preventing foam cell formation, lessening the secretion of cytokines and extracellular matrix in VSMC, preventing mononuclear cell infiltration, lessening inflammation, curtailing the destabilization of fibrous plaque, inhibiting the apoptosis of VSMC, modulating signal transduction and gene expression in VSMC, increasing the expression of connective tissue growth factor (CTGF) in VSMC (cell lines), preventing endothelial dysfunction related to cholesterol, modulating endothelial cells and the expression of adhesion molecules such as VCAM-1 and ICAM-1 on endothelial cells, preventing lysophosphatidylcholine (LPC)-induced endothelial dysfunction and the preservation of endothelial NO release, modulating monocytes, macrophages, T cells and mast cells, enhancing the expression of cytosolic phospholipase A2 (PLA2), cyclooxygenase, and vasodilating prostacyclin (PGI2) in endothelial cells, inhibiting thrombin formation, and reducing leukotriene synthesis [67][68][69][70][71][72][73][74][75][76].
In several clinical studies, vitamin E revealed contrasting findings. In a study examining the effect of 50 mg·day-1 synthetic vitamin E in a population with coronary heart disease, the results showed no effect on major cardiovascular events [77]. Another study showed 300 mg day-1 synthetic vitamin E had no effect on cardiovascular disease, including the rate of non-fatal myocardial infarction in patients with previous myocardial infarction [78]. Additionally, vitamin E did not significantly decrease the incidence of cardiovascular disease such as stroke [79]. In the Cambridge heart antioxidant study (CHAOS), vitamin E supplementation failed to have an impact on cardiovascular outcomes in patients at high risk of cardiovascular events [80].
5.2. Vitamin C
Vitamin C (ascorbate) is a water-soluble and ubiquitous antioxidant [7][23] with an ability to scavenge peroxyl radicals and HOCL [23][66], thus providing stability to the cell membrane. Fruit and vegetables, particularly citrus fruit, kiwi, cantaloupe, mango, strawberries, and peppers are rich sources of vitamin C [9]. It has various functions including: The improvement of nitric oxide-dependent vasomotor function [81], the enhancement of NOS activity (NO production) and the consequent augmentation of NO bioavailability, the improvement of endothelial dysfunction and vasodilation, the inhibition of cyclooxygenase, the diminishing of cell–cell adhesion [82], and the reduction of the chain-carrying α-tocopheroxyl radical to inhibit LDL peroxidation [83]. It also recycles other endogenous antioxidants, such as vitamin E [84]; discourages leukocyte aggregation and adhesion to the endothelium [9]; and scavenges ROS such as superoxide, hydroxyl radicals, peroxyl radicals, and many non-radicals, such as nitrosating agents and hydrochlorous acid [9].
A number of small-scale clinical studies have evaluated the effect of vitamin C on vascular health. The British Regional Heart Study demonstrated an inverse relation between plasma vitamin C concentration and endothelial dysfunction in men with no history of cardiovascular disease or diabetes [85]. Additionally, the European prospective investigation into cancer and nutrition (EPIC) Norfolk study showed the same results as the British Regional Heart Study in both men and women [86].
A large-scale study conducted over 20 years found that diets rich in vitamin C had no significant association with coronary heart disease [87].
5.3 . B Vitamins
B vitamins have a fundamental role in the metabolism of essential amino acids, with a specific influence on homocysteine and the antioxidant, glutathione [9]. Other significant activities of B vitamins entail scavenging hydroxyl and lipid peroxyl radicals, improving endothelial function, and ameliorating the coupling of endothelial NO synthase through the essential cofactor, tetrahydrobiopterin [88][89].
In a clinical study intake of folate, hydroxocobalamin, and pyridoxine, supplements for eight weeks decreased serum homocysteine to a normal range in patients with venous thrombosis [90].The vitamin intervention for stroke prevention (VISP) randomized controlled trial study demonstrated no significant effect of folate, hydroxocobalamin, and pyridoxine supplementation in decreasing incidence of coronary events or cardiovascular death [91]. The Cochrane systematic review reported no evidence to prevent cardiovascular events by using B vitamins [92].
5.4. Vitamin A and Carotenoids
Carotenoids are a large group of lipid soluble, colorful substances (yellow, orange, and red) such as α-carotene, β-carotene, β-cryptoxanthine, luteine and lycopene which occur extensively in fruit and vegetables [9].
They scavenge free radicals and prevent LDL peroxidation. β-Carotene can decrease plasma cholesterol levels by inhibiting HMG-CoA reductase (3-hydroxy-3-methyl-glutaryl-coenzyme A reductase). In addition, carotenoids are capable of increasing macrophage LDL receptor activity and reducing circulating LDL, inflammation, oxidative stress, and endothelial dysfunction [93][94]. A clinical study suggested an inverse relationship between the intake of β-carotene or retinol and risk of cardiovascular disease [95]. The US Preventative Task Force does not suggest β-carotene for the prevention of cardiovascular disease [96]. The Cochrane review on antioxidant consumption indicated that β-carotene and vitamin A significantly increase all-cause mortality [97].
5.5. Polyphenols
Polyphenols are the most abundant natural antioxidants possessing variable phenolic structures. They are found in fruit (especially apples), grains, vegetables, cereals, olive oil, dry legumes, chocolate and beverages, such as tea, coffee and wine [98]. These compounds are divided into several classes: Flavonoids, phenolic acids (e.g., caffeic acid and gallic acid), stilbenes (e.g., resveratrol), and lignans (e.g., secoisolariciresinol) [9]. Flavonoids, which are a major class of polyphenols, are subclassified as flavonols (e.g., quercetin), flavones (e.g., apigenin, luteolin), flavanones (naringenin, hesperetin), flavan-3-ols (catechins and their oligomers: Proanthocyanidins), isoflavones (e.g., genstein), and anthocyanins (e.g., delphinidin, cyanidin) [9][98].
The mechanistic effects of polyphenols involve: Suppressing ROS formation, scavenging ROS (both radical and non-radical oxygen), increasing the expression level of eNOS and the generation of NO or reducing NO oxidation by enhancing the intracellular free calcium concentration and by activating estrogen receptors in endothelial cells (ECs), blocking the action of xanthine oxidase and protein kinase C to prevent the production of the superoxide radical, and the protection of vascular endothelial cells and NO from oxidation. They also decrease redox-sensitive gene activation, preventing the expression of two major pro-angiogenic factors (vascular endothelial growth factor (VEGF) and matrix metalloproteinase-2 (MMP-2)) in smooth muscle cells, increase the production of major vasodilatory factors (NO, endothelium-derived hyperpolarizing factor (EDHF) as well as prostacyclin), inhibit angiogenesis (cell migration and proliferation of blood vessels), and also reduce platelet aggregation and hypertension [99][100][101][102][103][104].
Another important polyphenol that has received much attention is resveratrol (3, 5, 40-trihydroxystilbene), a stilbene polyphenol, which occurs in grapes, red wine and Polygonum cuspidatum. Resveratrol has established antioxidant properties which include the inhibition of lipid oxidation, the regulation of vasodilator and vasoconstrictor production, the inhibition of platelet aggregation, and the inhibition of the transcription factors NF-κB (Nuclear Factor kappaB) and AP-1 (Activator Protein 1) through an interaction with upstream signaling pathways and/or by decreasing pro-inflammatory mediators (TNF-α, IL) [98][105]. Clinical studies such as the Zupthen Elderly study showed a significant inverse association between flavonoid intake and coronary heart disease after 5 years of consumption [106][107]. In addition, the Rotterdam study revealed a significant inverse relationship between total flavonoid intake from the diet with myocardial infarction incidence [108]. The consumption of cocoa or chocolate is inversely associated with carotid atherosclerosis [109].
6. Synthetic Antioxidants: Probucol and Related Phenols
Probucol (2,6-di-tert-butyl-4-({2-[3,5-di-tert-butyl-4-hydroxyphenyl)sulfanyl) propan-2-yl}
sulfanyl)phenol) is a phenolic lipid-soluble antioxidant [7]. Its activities related to any antiatherosclerotic effectiveness consist of anti-inflammatory activity, the augmentation of endothelial function and repair, lessening oxidant production in vessel walls, attenuating atherosclerosis through the inhibition of LDL oxidation by blocking the production of oxidizing intermediates, inducing heme oxygenase-1 (HO-1) in arterial cells, inhibiting vasomotor dysfunction and fatty streak formation [81], reducing restenosis [7][110][111], inhibiting smooth muscle cell proliferation and adhesion molecule expression on endothelial cells, and promoting endothelium-dependent vasomotion [7].
Another important lipophilic and synthetic antioxidant is BO-653n (2, 3-dihydro-5-hydroxy-2,
2-dipentyl-4,6-di-tert-butylbenzofuran) (an analog of α-tocopherol) which inhibits the formation of atherosclerotic lesions [26], reduces α- tocopheroxyl radical and inhibits LDL oxidation in the intimal area [23][110][112][113].
7. Conclusions
Atherosclerosis is a major cause of morbidity and mortality in the developed world. Due to the factor that ROS and the generation of oxidized LDL are leading contributors to the progression of atherosclerosis, dietary supplements and antioxidants with low adverse effects may well represent a good therapeutic strategy to prevent the progression of the disease. Natural and synthetic antioxidants facilitate atherosclerosis treatment through a variety of mechanisms, including the inhibition of LDL oxidation, the reduction of generated reactive oxygen species, the inhibition of cytokine secretion, the prevention of atherosclerotic plaque formation and platelet aggregation, the prevention of mononuclear cell infiltration, the improvement of endothelial dysfunction and vasodilation, the promotion of NO bioavailability, the modulation of the expression of adhesion molecules such as VCAM-1 and ICAM-1 on endothelial cells, and the suppression of foam cell formation. It is not clear which of these different mechanisms of antioxidants action is more effective, but it seems that the use of multiple antioxidants is more effective target for antioxidant therapy.
References
- Steinberg, D. Atherogenesis in perspective: Hypercholesterolemia and inflammation as partners in crime. Nat. Med. 2002, 8, 1211–1217.
- Hennekens, C.H.; Gaziano, J.M. Antioxidants and heart disease: Epidemiology and clinical evidence. Clin. Cardiol. 1993, 16, 10–15.
- Ross, R. Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 1999, 340, 115.
- Singh, P.P.; Mahadi, F.; Roy, A.; Sharma, P. Reactive oxygen species, reactive nitrogen species and antioxidants in etiopathogenesis of diabetes mellitus type-2. Indian J. Clin. Biochem. 2009, 24, 324–342.
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143.
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422.
- Mashima, R.; Witting, P.K.; Stocker, R. Oxidants and antioxidants in atherosclerosis. Curr. Opin. Lipidol. 2001, 12, 411–418.
- Liu, Q.; Li, J.; Hartstone-Rose, A.; Wang, J.; Li, J.; Janicki, J.S.; Fan, D. Chinese herbal compounds for the prevention and treatment of atherosclerosis: Experimental evidence and mechanisms. Evid. Based Complement Alternat. Med. 2015, 2015, 1–15.
- Goszcz, K.; Deakin, S.J.; Duthie, G.G.; Stewart, D.; Leslie, S.J.; Megson, I.L. Antioxidants in cardiovascular therapy: Panacea or false hope? Front. Cardiovasc. Med. 2015, 2, 1–29.
- Niki, E. Antioxidants and atherosclerosis. Biochem. Soc. Trans. 2004, 32, 156–159.
- Brenner, C.; Pervaiz, S.; Clément, M.V.; Le Bras, M. Reactive oxygen species and the mitochondrial signaling pathway of cell death. Histol. Histopathol. 2005, 20, 205–219.
- Victor, V.M.; Apostolova, N.; Herance, R.; Hernandez-Mijares, A.; Rocha, M. Oxidative stress and mitochondrial dysfunction in atherosclerosis: Mitochondria-targeted antioxidants as potential therapy. Curr. Med. Chem. 2009, 16, 4654–4667.
- Li, H.; Horke, S.; Förstermann, U. Vascular oxidative stress, nitric oxide and atherosclerosis. Atherosclerosis 2014, 237, 208–219.
- Griendling, K.K.; FitzGerald, G.A. Oxidative stress and cardiovascular injury: Part I: Basic mechanisms and in vivo monitoring of ROS. Circulation 2003, 108, 1912–1916.
- Guzik, T.J.; West, N.E.; Black, E.; McDonald, D.; Ratnatunga, C.; Pillai, R.; Channon, K.M. Vascular superoxide production by NAD (P) H oxidase: Association with endothelial dysfunction and clinical risk factors. Circ. Res. 2000, 86, e85–e90.
- Mitra, S.; Deshmukh, A.; Sachdeva, R.; Lu, J.; Mehta, J.L. Oxidized low-density lipoprotein and atherosclerosis implications in antioxidant therapy. Am. J. Med. Sci. 2011, 342, 135–142.
- Ferri, C.; Grassi, D. Editorial Commentary Antioxidants and Beneficial Microvascular Effects. Hypertension 2010, 55, 1310–1311.
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84.
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322.
- Sies, H.; Stahl, W.; Sevanian, A. Nutritional, dietary and postprandial oxidative stress. J. Nutr. 2005, 135, 969–972.
- Jacob, R. Causal Role of Oxidants and Antioxidants in Atherosclerosis. Indian J. Cardiovasc. Dis. Women-WINCARS 2018, 3, 004–005.
- Ehara, S.; Ueda, M.; Naruko, T.; Haze, K.; Itoh, A.; Otsuka, M.; Komatsu, R.; Matsuo, T.; Itabe, H.; Takano, T. Elevated levels of oxidized low density lipoprotein show a positive relationship with the severity of acute coronary syndromes. Circulation 2001, 103, 1955–1960.
- Loenn, M.E.; Dennis, J.M.; Stocker, R. Actions of “antioxidants” in the protection against atherosclerosis. Free Radic. Biol. Med. 2012, 53, 863–884.
- Wang, T.; Palucci, D.; Law, K.; Yanagawa, B.; Yam, J.; Butany, J. Atherosclerosis: Pathogenesis and pathology. Diagn. Histopathol. 2012, 18, 461–467.
- Moradi, F.; Sewell, R.D.; Lorigooini, Z.; Rafieian-Kopaei, M. Immunosuppression-lipid metabolism interplay and medicinal plants in atherosclerosis: A review. Curr. Pharm. Des. 2018, 24, 2789–2793.
- Blake, G.J.; Ridker, P.M. Novel clinical markers of vascular wall inflammation. Circ. Res. 2001, 89, 763–771.
- Kirichenko, T.V.; Sobenin, I.A.; Nikolic, D.; Rizzo, M.; Orekhov, A.N. Anti-cytokine therapy for prevention of atherosclerosis. Phytomedicine 2016, 23, 1198–1210.
- Nikoforov, N.G.; Gratchev, A.N.; Sobenin, I.A.; Orekhov, A.N.; Kzhyhskowska, Y.G. Interaction of native and modified low density lipoprotein with intimal cells in atherosclerotic lesion. Patol. Fiziol. Eksp. Ter. 2013, 1, 109–117.
- Kishimoto, Y.; Yoshida, H.; Kondo, K. Potential anti-atherosclerotic properties of astaxanthin. Mar. Drugs 2016, 14, 35.
- Blankenberg, S.; Barbaux, S.; Tiret, L. Adhesion molecules and atherosclerosis. Atherosclerosis 2003, 170, 191–203.
- Erl, W.; Weber, P.C.; Weber, C. Monocytic cell adhesion to endothelial cells stimulated by oxidized low density lipoprotein is mediated by distinct endothelial ligands. Atherosclerosis 1998, 136, 297–303.
- Libby, P. Inflammation in atherosclerosis. Arter. Thromb. Vasc. Biol. 2012, 32, 2045–2051.
- Lind, L. Circulating markers of inflammation and atherosclerosis. Atherosclerosis 2003, 169, 203–214.
- Steinbrecher, U.P.; Parthasarathy, S.; Leake, D.S.; Witztum, J.L.; Steinberg, D. Modification of low density lipoprotein by endothelial cells involves lipid peroxidation and degradation of low density lipoprotein phospholipids. Proc. Natl. Acad. Sci. USA 1984, 81, 3883–3887.
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809.
- Patel, S.; Celermajer, D.S.; Bao, S. Atherosclerosis—Underlying inflammatory mechanisms and clinical implications. Int. J. Biochem. Cell Biol. 2008, 40, 576–580.
- Ouweneel, A.B.; Van Eck, M. Lipoproteins as modulators of atherothrombosis: From endothelial function to primary and secondary coagulation. Vascul. Pharmacol. 2016, 82, 1–10.
- Schönbeck, U.; Mach, F.; Sukhova, G.K.; Herman, M.; Graber, P.; Kehry, M.R.; Libby, P. CD40 ligation induces tissue factor expression in human vascular smooth muscle cells. Am. J. Pathol. 2000, 156, 7–14.
- Salvayre, R.; Negre-Salvayre, A.; Camaré, C. Oxidative theory of atherosclerosis and antioxidants. Biochimie 2016, 125, 281–296.
- Yang, H.; Zhou, L.; Wang, Z.; Roberts, L.J., II; Lin, X.; Zhao, Y.; Guo, Z. Overexpression of antioxidant enzymes in ApoE-deficient mice suppresses benzo (a) pyrene-accelerated atherosclerosis. Atherosclerosis 2009, 207, 51–58.
- Kirkman, H.N.; Gaetani, G.F. Mammalian catalase: A venerable enzyme with new mysteries. Trends Biochem. Sci. 2007, 32, 44–50.
- Shi, M.; Yang, H.; Motley, E.D.; Guo, Z. Overexpression of Cu/Zn-superoxide dismutase and/or catalase in mice inhibits aorta smooth muscle cell proliferation. Am. J. Hypertens. 2004, 17, 450–456.
- Wagner, A.H.; Kautz, O.; Fricke, K.; Zerr-Fouineau, M.; Demicheva, E.; Güldenzoph, B.; Bermejo, J.L.; Korff, T.; Hecker, M. Upregulation of glutathione peroxidase offsets stretch-induced proatherogenic gene expression in human endothelial cells. Arter. Thromb. Vasc. Biol. 2009, 29, 1894–1901.
- Berndt, C.; Lillig, C.H.; Holmgren, A. Thiol-based mechanisms of the thioredoxin and glutaredoxin systems: Implications for diseases in the cardiovascular system. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1227–H1236.
- Ahsan, M.K.; Lekli, I.; Ray, D.; Yodoi, J.; Das, D.K. Redox regulation of cell survival by the thioredoxin superfamily: An implication of redox gene therapy in the heart. Antioxid. Redox Signal. 2009, 11, 2741–2758.
- Zhang, H.; Luo, Y.; Zhang, W.; He, Y.; Dai, S.; Zhang, R.; Huang, Y.; Bernatchez, P.; Giordano, F.J.; Shadel, G.; et al. Endothelial-specific expression of mitochondrial thioredoxin improves endothelial cell function and reduces atherosclerotic lesions. Am. J. Pathol. 2007, 170, 1108–1120.
- Jones, C.M.; Lawrence, A.; Wardman, P.; Burkitt, M.J. Kinetics of superoxide scavenging by glutathione: An evaluation of its role in the removal of mitochondrial superoxide. Biochem. Soc. Trans. 2003, 31, 1337–1339.
- Rosenblat, M.; Coleman, R.; Aviram, M. Increased macrophage glutathione content reduces cell-mediated oxidation of LDL and atherosclerosis in apolipoprotein E-deficient mice. Atherosclerosis 2002, 163, 17–28.
- Forsmark-AndrÉe, P.; Dallner, G.; Ernster, L. Endogenous ubiquinol prevents protein modification accompanying lipid peroxidation in beef heart submitochondrial particles. Free Radic. Biol. Med. 1995, 19, 749–757.
- Tomasetti, M.; Littarru, G.P.; Stocker, R.; Alleva, R. Coenzyme Q10 enrichment decreases oxidative DNA damage in human lymphocytes. Free Radic. Biol. Med. 1999, 27, 1027–1032.
- Bowry, V.W.; Mohr, D.; Cleary, J.; Stocker, R. Prevention of tocopherol-mediated peroxidation in ubiquinol-10-free human low density lipoprotein. J. Biol. Chem. 1995, 270, 5756–5763.
- Witting, P.K.; Pettersson, K.; Stocker, R. Anti-atherogenic effect of coenzyme Q10 in apolipoprotein E gene knockout mice. Free Radic. Biol. Med. 2000, 29, 295–305.
- Thomas, S.R.; Leichtweis, S.B.; Pettersson, K.; Croft, K.D.; Mori, T.A.; Brown, A.J.; Stocker, R. Dietary cosupplementation with vitamin E and coenzyme Q10 inhibits atherosclerosis in apolipoprotein E gene knockout mice. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 585–593.
- Neužil, J.; Stocker, R. Bilirubin attenuates radical-mediated damage to serum albumin. FEBS Lett. 1993, 331, 281–284.
- Kawamura, K.; Ishikawa, K.; Wada, Y.; Kimura, S.; Matsumoto, H.; Kohro, T.; Itabe, H.; Kodama, T.; Maruyama, Y. Bilirubin from heme oxygenase-1 attenuates vascular endothelial activation and dysfunction. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 155–160.
- Nakao, A.; Murase, N.; Ho, C.; Toyokawa, H.; Billiar, T.R.; Kanno, S. Biliverdin administration prevents the formation of intimal hyperplasia induced by vascular injury. Circulation 2005, 112, 587–591.
- Öllinger, R.; Bilban, M.; Erat, A.; Froio, A.; McDaid, J.; Tyagi, S.; Csizmadia, E.; Graça-Souza, A.V.; Liloia, A.; Soares, M.P.; et al. A natural inhibitor of vascular smooth muscle cell proliferation. Circulation 2005, 112, 1030–1039.
- Kang, S.J.; Lee, C.; Kruzliak, P. Effects of serum bilirubin on atherosclerotic processes. Ann. Med. 2014, 46, 138–147.
- Hayashi, S.; Takamiya, R.; Yamaguchi, T.; Matsumoto, K.; Tojo, S.J.; Tamatani, T.; Kitajima, M.; Makino, N.; Ishimura, Y.; Suematsu, M. Induction of heme oxygenase-1 suppresses venular leukocyte adhesion elicited by oxidative stress: Role of bilirubin generated by the enzyme. Circ. Res. 1999, 85, 663–671.
- Öllinger, R.; Yamashita, K.; Bilban, M.; Erat, A.; Kogler, P.; Thomas, M.; Csizmadia, E.; Usheva, A.; Margreiter, R.; Bach, F.H. Bilirubin and biliverdin treatment of atherosclerotic diseases. Cell Cycle 2007, 6, 39–43.
- Ames, B.N.; Cathcart, R.; Schwiers, E.; Hochstein, P. Uric acid provides an antioxidant defense in humans against oxidant-and radical-caused aging and cancer: A hypothesis. Proc. Natl. Acad. Sci. USA 1981, 78, 6858–6862.
- Zhang, W.J.; Bird, K.E.; McMillen, T.S.; LeBoeuf, R.C.; Hagen, T.M.; Frei, B. Dietary alpha-lipoic acid supplementation inhibits atherosclerotic lesion development in apolipoprotein E-deficient and apolipoprotein E/low-density lipoprotein receptor-deficient mice. Circulation 2008, 117, 421–428.
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 1149–1160
- Heitzer, T.; Finckh, B.; Albers, S.; Krohn, K.; Kohlschütter, A.; Meinertz, T. Beneficial effects of α-lipoic acid and ascorbic acid on endothelium-dependent, nitric oxide-mediated vasodilation in diabetic patients: Relation to parameters of oxidative stress. Free Radic. Biol. Med. 2001, 31, 53–61.
- Ogborne, R.M.; Rushworth, S.A.; O’Connell, M.A. Mediated by Nuclear Factor Erythroid 2-Related Factor 2 and p38 Mitogen-Activated Protein Kinase in Human Monocytic Cells. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2100–2105.
- Siekmeier, R.; Steffen, C.; März, W. Role of oxidants and antioxidants in atherosclerosis: Results of in vitro and in vivo investigations. J. Cardiovasc. Pharmacol. Ther. 2007, 12, 265–282.
- Villacorta, L.; Graça-Souza, A.V.; Ricciarelli, R.; Zingg, J.M.; Azzi, A. α-Tocopherol induces expression of connective tissue growth factor and antagonizes tumor necrosis factor-α–mediated downregulation in human smooth muscle cells. Circ. Res. 2003, 92, 104–110.
- Villacorta, L.; Azzi, A.; Zingg, J.M. Regulatory role of vitamins E and C on extracellular matrix components of the vascular system. Mol. Asp. Med. 2007, 28, 507–537.
- Munteanu, A.; Zingg, J.M. Cellular, molecular and clinical aspects of vitamin E on atherosclerosis prevention. Mol. Asp. Med. 2007, 28, 538–590.
- Ricciarelli, R.; Zingg, J.M.; Azzi, A. Vitamin E reduces the uptake of oxidized LDL by inhibiting CD36 scavenger receptor expression in cultured aortic smooth muscle cells. Circulation 2000, 102, 82–87.
- Ricciarelli, R.; Zingg, J.M.; Azzi, A. Vitamin E: Protective role of a Janus molecule. FASEB J. 2001, 15, 2314–2325.
- Ricciarelli, R.; Maroni, P.; Özer, N.; Zingg, J.M.; Azzi, A. Age-dependent increase of collagenase expression can be reduced by α-tocopherol via protein kinase C inhibition. Free Radic. Biol. Med. 1999, 27, 729–737.
- Stewart-Lee, A.L.; Forster, L.A.; Nourooz-Zadeh, J.; Ferns, G.A.; Anggård, E.E. Vitamin E protects against impairment of endothelium-mediated relaxations in cholesterol-fed rabbits. Arterioscler. Thromb. Vasc. Biol. 1994, 14, 494–499. [CrossRef]
- Orbe, J.; Rodrıguez, J.A.; Arias, R.; Belzunce, M.; Nespereira, B.; Perez-Ilzarbe, M.; Roncal, C.; Paramo, J.A. Antioxidant vitamins increase the collagen content and reduce MMP-1 in a porcine model of atherosclerosis: Implications for plaque stabilization. Atherosclerosis 2003, 167, 45–53. [CrossRef]
- Devaraj, S.; Hugou, I.; Jialal, I. α-Tocopherol decreases CD36 expression in human monocyte-derived macrophages. J. Lipid Res. 2001, 42, 521–527.
- Venugopal, S.K.; Devaraj, S.; Jialal, I. RRR-α-tocopherol decreases the expression of the major scavenger receptor, CD36, in human macrophages via inhibition of tyrosine kinase (Tyk2). Atherosclerosis 2004, 175, 213–220.
- Rapola, J.M.; Virtamo, J.; Ripatti, S.; Huttunen, J.K.; Albanes, D.; Taylor, P.R.; Heinonen, O.P. Randomised trial of α-tocopherol and β-carotene supplements on incidence of major coronary events in men with previous myocardial infarction. Lancet 1997, 349, 1715–1720.
- GISSI-Prevenzione Investigators. Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: Results of the GISSI-Prevenzione trial. Lancet 1999, 354, 447–455.
- Heart Outcomes Prevention Evaluation Study Investigators. Vitamin E supplementation and cardiovascular events in high-risk patients. N. Engl. J. Med. 2000, 342, 154–160.
- Stephens, N.G.; Parsons, A.; Schofield, P.M.; Kelly, F.; Cheeseman, K.; Mitchinson, M.J. Randomised controlled trial of vitamin E in patients with coronary disease: Cambridge heart antioxidant study (CHAOS). Lancet
- 1996, 347, 781–786.
- Thomson, M.J.; Puntmann, V.; Kaski, J.C. Atherosclerosis and oxidant stress: The end of the road for antioxidant vitamin treatment? Cardiovasc. Drugs Ther. 2007, 21, 195–210.
- Lykkesfeldt, J.; Christen, S.; Wallock, L.M.; Chang, H.H.; Jacob, R.A.; Ames, B.N. Ascorbate is depleted by smoking and repleted by moderate supplementation: A study in male smokers and nonsmokers with matched dietary antioxidant intakes. Am. J. Clin. Nutr. 2000, 71, 530–536.
- Kagan, V.E.; Serbinova, E.A.; Forte, T.; Scita, G.; Packer, L. Recycling of vitamin E in human low density lipoproteins. J. Lipid Res. 1992, 33, 385–397.
- Wannamethee, S.G.; Lowe, G.D.; Rumley, A.; Bruckdorfer, K.R.; Whincup, P.H. Associations of vitamin C status, fruit and vegetable intakes, and markers of inflammation and hemostasis. Am. J. Clin. Nutr. 2006, 83, 567–574.
- Sargeant, L.A.; Khaw, K.T.; Bingham, S.; Day, N.E.; Luben, R.N.; Oakes, S.; Welch, A.; Wareham, N.J. Fruit and vegetable intake and population glycosylated haemoglobin levels: The EPIC-Norfolk Study. Eur. J. Clin. Nutr. 2001, 55, 342–348.
- Gale, C.R.; Martyn, C.N.; Winter, P.D.; Cooper, C. Vitamin C and risk of death from stroke and coronary heart disease in cohort of elderly people. BMJ 1995, 310, 1563–1566.
- Joshi, R.; Adhikari, S.; Patro, B.S.; Chattopadhyay, S.; Mukherjee, T. Free radical scavenging behavior of folic acid: Evidence for possible antioxidant activity. Free Radic. Biol. Med. 2001, 30, 1390–1399.
- Antoniades, C.; Shirodaria, C.; Crabtree, M.; Ratnatunga, C.; Pillai, R.; Neubauer, S.; Channon, K.M. Low-and high-dose treatment with folic acid, equally improve endothelial function and decrease superoxide production in human vessels via tetrahydrobiopterin-mediated eNOS coupling. Eur. Heart J. 2007, 28, 741.
- den Heijer, M.; Brouwer, I.A.; Bos, G.M.; Blom, H.J.; van der Put, N.M.; Spaans, A.P.; Rosendaal, F.R.; Thomas, C.M.; Haak, H.L.; Wijermans, P.W.; et al. Vitamin supplementation reduces blood homocysteine levels: A controlled trial in patients with venous thrombosis and healthy volunteers. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 356–361.
- Toole, J.F.; Malinow, M.R.; Chambless, L.E.; Spence, J.D.; Pettigrew, L.C.; Howard, V.J.; Sides, E.G.; Wang, C.H.; Stampfer, M. Lowering homocysteine in patients with ischemic stroke to prevent recurrent stroke, myocardial infarction, and death: The Vitamin Intervention for stroke Prevention (VISP) randomized controlled trial. JAMA 2004, 291, 565–575.
- Marti-Carvajal, A.J.; Sola, I.; Lathyris, D.; Salanti, G. Homocysteine lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2009, 4, CD006612.
- Fuhrman, B.; Elis, A.; Aviram, M. Hypocholesterolemic effect of lycopene and β-carotene is related to suppression of cholesterol synthesis and augmentation of LDL receptor activity in macrophages. Biochem. Biophys. Res. Commun. 1997, 233, 658–662.
- Rao, A.V. Lycopene, tomatoes, and the prevention of coronary heart disease. Exp. Biol. Med. 2002, 227, 908–913.
- Omenn, G.S.; Goodman, G.; Thornquist, M.; Barnhart, S.; Balmes, J.; Cherniack, M.; Cullen, M.; Glass, A.; Keogh, J.; Liu, D.; et al. Chemoprevention of lung cancer: The beta-Carotene and Retinol Efficacy Trial (CARET) in high-risk smokers and asbestos-exposed workers. IARC Sci. Publ. 1996, 136, 67–85.
- Moyer, V.A. Vitamin, mineral, and multivitamin supplements for the primary prevention of cardiovascular disease and cancer: US Preventive Services Task Force recommendation statement. Ann. Intern. Med. 2014, 160, 558–564.
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2012,3, CD007176.
- Grassi, D.; Desideri, G.; Ferri, C. Flavonoids: Antioxidants against atherosclerosis. Nutrients 2010, 2, 889–902.
- Wang, S.; Melnyk, J.P.; Tsao, R.; Marcone, M.F. How natural dietary antioxidants in fruits, vegetables and legumes promote vascular health. Food Res. Int. 2011, 44, 14–22.
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S.
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Asp. Med. 2010, 31, 435–445.
- Auger, C.; Kim, J.H.; Trinh, S.; Chataigneau, T.; Popken, A.M.; Schini-Kerth, V.B. Fruit juice-induced endothelium-dependent relaxations in isolated porcine coronary arteries: Evaluation of different fruit juices and purees and optimization of a red fruit juice blend. Food Funct. 2011, 2, 245–250.
- Han, X.; Shen, T.; Lou, H. Dietary polyphenols and their biological significance. Int. J. Mol. Sci. 2007, 8, 950–988.
- De Nigris, F.; Williams-Ignarro, S.; Lerman, L.O.; Crimi, E.; Botti, C.; Mansueto, G.; D’Armiento, F.P.; De Rosa, G.; Sica, V.; Ignarro, L.J.; et al. Beneficial effects of pomegranate juice on oxidation-sensitive genes and endothelial nitric oxide synthase activity at sites of perturbed shear stress. Proc. Natl. Acad. Sci. USA 2005, 102, 4896–4901.
- Rivera, L.; Morón, R.; Zarzuelo, A.; Galisteo, M. Long-term resveratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem. Pharmacol. 2009, 77, 1053–1063.
- Hertog, M.G.; Feskens, E.J.; Kromhout, D.; Hollman, P.C.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011.
- Hertog, M.G.; Feskens, E.J.; Kromhout, D. Antioxidant flavonols and coronary heart disease risk. Lancet 1997, 349, 699.
- Geleijnse, J.M.; Launer, L.J.; van der Kuip, D.A.; Hofman, A.; Witteman, J. Inverse association of tea and flavonoid intakes with incident myocardial infarction: The Rotterdam Study. Am. J. Clin. Nutr. 2002, 75, 880–886.
- Mursu, J.; Nurmi, T.; Tuomainen, T.P.; Ruusunen, A.; Salonen, J.T.; Voutilainen, S. The intake of flavonoids and carotid atherosclerosis: The Kuopio ischaemic heart disease risk factor study. Br. J. Nutr. 2007, 98,814–818.
- Stocker, R. Dietary and pharmacological antioxidants in atherosclerosis. Curr. Opin. Lipidol. 1999, 10, 589–597.
- Violi, F.; Micheletta, F.; Iuliano, L. Antioxidants and atherosclerosis. Eur. Heart J. 2002, 4, B17–B21.
- Noguchi, N.; Niki, E. Phenolic antioxidants: A rationale for design and evaluation of novel antioxidant drug for atherosclerosis. Free Radic. Biol. Med. 2000, 28, 1538–1546.
- Shiao, M.S.; Chiu, J.J.; Chang, B.W.; Wang, J.; Jen, W.P.; Wu, Y.J.; Chen, Y.L. In search of antioxidants and anti-atherosclerotic agents from herbal medicines. Biofactors 2008, 34, 147–157.

