Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Chunzhang Yang | + 2485 word(s) | 2485 | 2021-06-24 05:29:04 | | | |
| 2 | Vivi Li | -18 word(s) | 2467 | 2021-06-25 08:23:22 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Yang, C. Isocitrate Dehydrogenase Mutations in Glioma. Encyclopedia. Available online: https://encyclopedia.pub/entry/11292 (accessed on 08 August 2026).
Yang C. Isocitrate Dehydrogenase Mutations in Glioma. Encyclopedia. Available at: https://encyclopedia.pub/entry/11292. Accessed August 08, 2026.
Yang, Chunzhang. "Isocitrate Dehydrogenase Mutations in Glioma" Encyclopedia, https://encyclopedia.pub/entry/11292 (accessed August 08, 2026).
Yang, C. (2021, June 25). Isocitrate Dehydrogenase Mutations in Glioma. In Encyclopedia. https://encyclopedia.pub/entry/11292
Yang, Chunzhang. "Isocitrate Dehydrogenase Mutations in Glioma." Encyclopedia. Web. 25 June, 2021.
Copy Citation
Mutations in isocitrate dehydrogenase (IDH) are commonly observed in lower-grade glioma and secondary glioblastomas. IDH mutants confer a neomorphic enzyme activity that converts α-ketoglutarate to an oncometabolite D-2-hydroxyglutarate, which impacts cellular epigenetics and metabolism. IDH mutation establishes distinctive patterns in metabolism, cancer biology, and the therapeutic sensitivity of glioma. Thus, a deeper understanding of the roles of IDH mutations is of great value to improve the therapeutic efficacy of glioma and other malignancies that share similar genetic characteristics.
IDH mutation
glioma
cancer
therapy resistance
1. Introduction
In 2008, compelling research showed that mutations in isocitrate dehydrogenase (IDH1 and IDH2) are frequently identified in the World Health Organization (WHO) grade II/III gliomas and secondary glioblastomas (GBMs). In contrast, these mutations are rare in primary GBM patients [1]. In 2009, Yan et al. [2] showed that IDH1 and IDH2 mutations frequently occur in WHO grade II/III astrocytomas and oligodendrogliomas. Besides gliomas, IDH mutations also occur in other non-central nervous system (CNS) malignancies, including acute myeloid leukemia (AML) [3][4], intrahepatic cholangiocarcinoma [5][6], chondrosarcoma [7], and melanoma [8][9]. The mutations are confined to a single arginine residue (Arg132) in IDH1 or two arginine residues (Arg172 and Arg140) in IDH2 [10][11]. The mutations commonly cause amino acid substitutions, which localize at the active sites of the enzymes and alter the catalytic functions of IDH enzymes. In contrast to wild-type IDH, which transforms isocitrate into α-ketoglutarate (α-KG), the mutated IDHs convert α-KG into D-2-hydroxyglutarate (D-2-HG) [12]. The altered catalytic activity that occurs because of cancer-associated IDH mutations was later termed “neomorphic activity”. The overproduction of the oncometabolite D-2-HG leads to widespread physiological consequences, including profound effects on cellular metabolism [13][14], epigenetic shift [15][16][17][18], genomic instability [19][20][21][22][23], and redox homeostasis [24][25][26][27][28][29]. IDH mutations are considered founder events for oncogenesis, through which an ancestor glial cell commits to malignant transformation. On the other hand, the mutant IDH enzyme brings about substantial changes in cancer biology, thereby establishing novel therapeutic vulnerabilities that are not commonly identified in other neoplasms.
2. Clinical Indications Involving the Discovery of IDH-Mutated Glioma
2.1. Clinical Classification of Gliomas
The 2016 WHO classification of CNS tumors has suggested the use of integrated phenotypic and genotypic characterization, which provides an increased level of objectivity [30]. In particular, IDH mutations have become some of the most important parameters in the differential diagnosis of gliomas. For example, diffuse astrocytomas often harbor IDH mutations, followed by other mutations such as TP53 and ATRX. Oligodendrogliomas are characterized by IDH mutations along with 1p/19q co-deletion (potentially along with CIC and FUBP1 mutations). The IDH mutation status is also useful for the differential diagnosis of primary and secondary GBMs [30][31][32]. Moreover, as IDH mutations frequently induce genome-wide DNA and histone hypermethylation, the introduction of methylation profiling allows for further improving the accuracy of glioma classification. Recently, Jaunmuktane et al. [33] demonstrated a diagnostic algorithm that integrated histology, molecular signature, and methylation array, and improved the diagnostic approach. Thus, the IDH mutation status is of great value in glioma classification and the selection of appropriate therapeutic strategies.
2.2. Radiology—D-2-HG Imaging
D-2-HG is a novel metabolite that accumulates in extremely high levels in glioma cells, but is absent in normal brain cells. The drastic contrast in cellular D-2-HG levels suggests that this oncometabolite could be an ideal biomarker for clinical monitoring and diagnosis among patients with IDH-mutated cancers [34]. Several hallmark studies have developed noninvasive radiologic methods for the detection of D-2-HG, such as magnetic resonance spectroscopy (MRS). In IDH mutant gliomas, D-2-HG accumulates to sufficient levels as a brain metabolite, which renders its visibility on MRS. These levels are 2–3 orders of magnitude higher than those found in the adjacent normal brain tissues [34]. Andronesi et al. [35] reported that D-2-HG was detected unambiguously in mutant IDH1 glioma in vivo using 2D correlation spectroscopy (COSY) and J-difference spectroscopy. Several other studies have also reported that D-2-HG is detected among glioma patients or in animal models using the short echo times (TEs) method [36][37][38][39]. On the other hand, D-2-HG levels were detected in glioma patients using long TE methods and J-difference spectroscopy with 100% sensitivity [40][41]. The application of long TE methods in D-2-HG detection has been confirmed in several subsequent reports, with increased sensitivity and specificity [42][43]. Overall, the noninvasive detection of D-2-HG has been proven to be a valuable diagnostic and prognostic biomarker. D-2-HG imaging provides a useful approach to the clinical management of patients with IDH-mutated glioma. Fluctuations in D-2-HG levels may provide crucial longitudinal data for the determination of disease progression and therapy response [34].
2.3. Disease Outcomes—Prolonged Survival
In 2008, Parsons et al. [1] first reported that mutations in IDH1 occurred in most patients with secondary GBM, and were associated with better overall survival (OS). Similar trends were reported in numerous studies using various datasets [44][45][46][47][48][49]. For example, using a large clinical dataset, Yan et al. [2] reported that GBM patients harboring IDH1 or IDH2 mutations tend to have a prolonged median OS compared with patients with IDH wild-type GBM. Similar findings were also observed among patients with anaplastic astrocytoma. The median OS was 65 months for patients with IDH mutant disease, compared with 20 months for those with IDH wild-type disease. Moreover, the progression-free survival (PFS) was also improved among GBM patients with IDH mutations compared with their counterparts [45]. Secondary genetic alterations, such as TP53/ATRX mutations and 1p/19q co-deletion, predispose patients with IDH-mutated gliomas to slightly different OS and disease-free survival (DFS; Figure 1A,B). Several studies have reported that IDH mutations are associated with younger age at diagnosis and limited genome alterations among patients with WHO grade II/III gliomas and GBMs, which may bias the disease outcome (Figure 1C,D) [1][2][50][51]. However, in a multivariate analysis, Sanson et al. [45] showed that the IDH mutation status is an independent predictor of favorable outcomes among glioma patients.
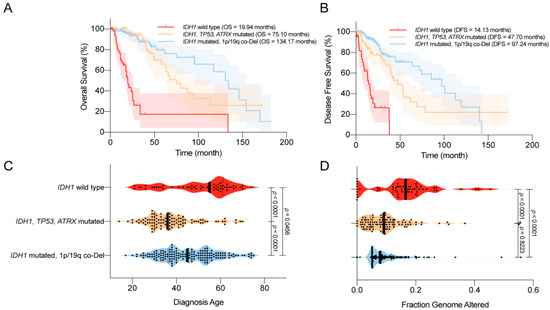
Figure 1. Clinical features of the World Health Organization (WHO) grade II/III IDH-mutated glioma. (A) Overall survival (OS) of glioma patients according to IDH1 status. IDH1 mutations are associated with prolonged OS. (B) Disease-free survival (DFS) of glioma patients according to IDH1 status. IDH1 mutations are associated with prolonged DFS. (C) Age at diagnosis among glioma patients according to IDH1 status. IDH1 mutations are associated with a younger age at diagnosis. (D) The distribution of genome alterations in glioma according to IDH1 status. IDH1 mutations are associated with fewer genome alterations. The data are visualized in cBioPortal [52][53].
2.4. Complications—Epilepsy and Secondary GBM
Epileptic seizure is one of the most common complications among patients with glioma, particularly those with LGGs (up to 90%) [54][55][56][57]. Severe seizures impair the quality of life and neurocognition function among glioma patients [58]. Considering the high incidence of IDH mutations in LGG, it is likely that the epileptic changes are relevant to the unique patterns in the tumor microenvironment, which is associated with IDH mutants. Numerous studies have indicated that mutations in IDH are associated with a high prevalence of epilepsy [59][60][61][62][63]. For example, Chen et al. [62] showed that IDH mutations are independently correlated with seizures, regardless of WHO grade. A recent study suggested that D-2-HG overproduction in the tumor microenvironment plays a major role in glioma-related epilepsy. D-2-HG is structurally similar to glutamate, which is the predominant excitatory neurotransmitter in the CNS. Thus, D-2-HG may act as an analog of glutamate, which leads to the abnormal firing of neurons through activating N-Methyl-d-aspartic acid (NMDA) receptors, and hence epileptic changes. Treating cultured rat cortical neurons with exogenous D-2-HG resulted in an elevated firing rate [62]. By mimicking the activity of glutamate, the increased level of D-2-HG mediates the abnormal neuronal activity and leads to glioma-related epilepsy [35][64][65]. However, three millimolar D-2-HG induced an elevated burst frequency in the neuronal network in vitro [62], whereas this dose is over 30 times higher than the glutamate concentration for excitotoxicity [66]. More effort is urged in order to elucidate the detailed molecular mechanism of the epileptic changes in IDH-mutated glioma. Because of the association between IDH mutations and seizures, therapies that target mutant IDH, such as mutant IDH inhibitors, could diminish D-2-HG production and potentially reduce epileptic seizures [67].
2.5. Sensitivity to Radiotherapy and Chemotherapy
Clinical data have shown that IDH mutant gliomas tend to exhibit a better disease outcome compared with wild-type IDH tumors. Several studies have explained that the favorable prognosis of IDH mutant gliomas is due to their increased sensitivity to radiotherapy and chemotherapy [68][69]. IDH mutant gliomas likely harbor defects in multiple DNA repair pathways, which render them vulnerable to radiotherapy- or chemotherapy-induced DNA damage [19][22]. These findings indicate that IDH mutation could serve as an important predictive factor for treatment response among glioma patients. For example, Houillier et al. [69] reported that IDH1 mutation is an independent predictor of temozolomide response among LGG patients. IDH1 mutations combined with 1p/19q co-deletion further improved the treatment response. Hartman et al. [70] also reported that IDH1 status is an important predictor of disease-free survival (DFS) and OS among patients undergoing adjuvant therapy. In another study conducted by van de Bent et al. [71], no correlation was found between IDH1 mutations and disease outcome in response to procarbazine (Matulane), lomustine (CCNU), and vincristine (Oncovin) chemotherapy.
3. Novel Molecular Targeting for IDH-Mutated Glioma
3.1. IDH Mutant Inhibitors
Because of the critical roles played by IDH mutations in the malignant transformation of glioma, targeting the neomorphic activity of IDH mutants has been heavily proposed as a direct therapeutic approach. In the past decade, several attempts have been made to develop small molecular compounds that directly inhibit mutant IDH enzymes. In 2012, the first-in-class mutant IDH inhibitor was discovered, which showed a specific and potent inhibitory effect on D-2-HG production in IDH mutant U87 cells and xenograft models [72]. Later, Rohle et al. [67] reported a novel synthetic inhibitor of IDH mutant, AGI-5198, which blocked D-2-HG production and subsequently reversed the malignant transformation effect of IDH mutations. Besides glioma, the inhibition of mutant IDH promotes differentiation in leukemia harboring IDH mutations [6]. With the promising findings regarding AGI-5198, second-generation mutant IDH inhibitors are under development and are undergoing evaluation in clinical studies. For example, ivosidenib (AG-120) and vorasidenib (AG-881) have been tested in AML and glioma with IDH mutations [73][74][75][76]. In a recent phase I clinical study with ivosidenib in IDH1-mutated advanced glioma conducted by Mellinghoff et al. [77], the mutant IDH inhibitor appeared to be well-tolerated throughout the experiment, which paved the way for subsequent clinical studies to evaluate its therapeutic efficacy. Although the IDH mutant enzyme inhibitors suppress malignancy, several studies have suggested that this inhibitor reduces D-2-HG production and relieves the burden on the DNA repair pathway, resulting in chemoresistance to other therapies, such as PARP inhibitors [23][78]. More effort is urged to explore the strategy of combining IDH mutant inhibitors with other glioma therapies in order to improve the clinical outcome.
3.2. Targeting Hypermethylation Phenotype
Genome-wide DNA and histone hypermethylation is a unique signature in IDH-mutated glioma, which is closely related to gliomagenesis by promoting oncogene expression and inhibiting tumor suppressors [79]. This rectification of the epigenetic shift could be a reasonable strategy for halting D-2-HG-driven oncogenesis and the malignant phenotype. DNA-demethylating agents such as 5-azacytidine or 5-aza-2′-deoxycytidine (decitabine) irreversibly bind to DNA methyltransferases (DNMTs) and inhibit the process of DNA methylation. The D-2-HG-induced hypermethylation phenotype was reversed by demethylating compounds, and cell proliferation was suppressed in vitro and in vivo [80][81][82]. Several clinical trials are evaluating the therapeutic effects of 5-azacytidine among patients with recurrent gliomas with IDH mutations (NCT03666559 and NCT03684811). On the other hand, inhibitors targeting histone methyltransferases inhibitors are also being investigated for IDH-mutated gliomas, as an alternative strategy to rectify the D-2-HG-associated hypermethylation phenotype. It is reported that H3K9 methyltransferase G9a is correlated to the development and progression of glioma, and its inhibitor BIX-01294 showed repressive effects on gliomas cells [83].
3.3. Targeting DNA Repair Pathways
As previously mentioned, IDH mutant gliomas exhibit defects in multiple DNA repair pathways. High levels of D-2-HG inhibit the activity of DNA oxidative demethylases, such as AlκB homolog 2/3 (ALKBH2/3) [19]. Several seminal studies have also indicated that D-2-HG compromises HR DNA repair, establishing a “BRCAness” in this type of malignancy [23][84]. In addition, IDH mutation-associated G-CIMP resulted in the methylation of the promoter region of O-6-methylguanine-DNA methyltransferase (MGMT), which reduced MGMT expression and led to increased sensitivity to alkylating agents [46][85][86]. Our recent study indicated that IDH mutations led to defects in NAD metabolism, which compromised PARP-associated HR, as PARP repairs DNA damage in an NAD+ dependent manner [22][87]. With the identification of the DNA repair deficiency in IDH-mutated glioma, numerous studies have attempted to evaluate DNA repair inhibitors, which may serve as a potential sensitization strategy. Several other groups and as well as ours reported that a combination of PARP inhibitors, such as olaparib, with temozolomide or radiotherapy, led to synergistic lethality in IDH mutant glioma cells [21][22][23]. Several phase I/II clinical trials are currently recruiting patients to investigate the therapeutic effect of the PARP inhibitors, pamiparib (BGB-290) or olaparib, combined with temozolomide in IDH mutant gliomas (NCT03914742, NCT03749187, and NCT03212274).
3.4. Targeting Anti-Oxidative Pathways
Redox homeostasis has been reported to be greatly impacted by IDH mutations, highlighted by profoundly elevated levels of oxidative stress [24][25][26][27][28]. As a result, ROS scavenging pathways are widely mobilized in the context of IDH mutation, so as to maintain cellular metabolism, thereby supporting cellular growth and survival. These findings suggest that the antioxidant pathway plays an essential role in IDH-mutated glioma. Targeting anti-oxidative pathways may be more effective in glioma with IDH mutations. Our recent study showed that NRF2-governed anti-oxidative pathways, such as that regarding de novo glutathione synthesis, were widespread in IDH mutant gliomas. The blockade of NRF2 using natural compound inhibitors, brusatol, or triptolide significantly increased oxidative damage and subsequently suppressed the growth of IDH mutant xenografts with prolonged OS [25][27][28][88]. The concept of targeting redox homeostasis in IDH mutant cancers has shown a potential therapeutic value. The development of pharmacological grade NRF2 inhibitors is needed urgently for potential clinical translation.
3.5. Targeting Metabolic Reprogramming
D-2-HG is a metabolite that is absent in normal cells. The production of large quantities of D-2-HG inevitably depletes a substantial amount of carbohydrate from the Krebs cycle. Several hallmark studies have demonstrated the presence of depleted metabolic pathways in IDH-mutated cells. For example, glutamate metabolism is greatly altered in IDH mutant glioma, as mentioned before. The glutamate level is significantly lower in IDH mutant cancers, which leads to an increased dependence on glutaminolysis to compensate for the metabolism [29][89][90][91]. Several studies have reported that a blockade of glutaminase activity results in the suppression of IDH mutant glioma and AML. Seltzer et al. and Emadi et al. [89][90] reported that bis-2-(5-phenylacetamido-1,2,4-thiadiazol-2-yl) ethyl sulfide (BPTES), an inhibitor of glutaminase, selectively suppresses tumor growth in IDH mutant glioma and AML by targeting the fragile glutamine metabolism. Another glutaminase inhibitor (CB-839) was also reported to induce selective radio-sensitivity in IDH mutant cancers [29] and terminal differentiation in IDH mutant AML [91]. An ongoing phase I clinical trial is investigating the side effects and the best dose of CB-839, in combination with radiation therapy and temozolomide, for treating IDH-mutated diffuse or anaplastic astrocytoma (NCT03528642). In addition, IDH mutations lead to the depletion of NAD+ because of the increased methylation of the promoter region of NAPRT1, the rate-limiting enzyme in NAD+ biosynthesis, and suppression of the expression of NAPRT1. This renders the IDH mutant glioma vulnerable to inhibition through the nicotinamide phosphoribosyltransferase (NAMPT) catalyzed NAD+ salvage pathway [92]. Moreover, Tateishi et al. [93] showed that NAMPT inhibitors further sensitized IDH mutant cancer cells to alkylating agents, such as temozolomide, as PARP activation consumes NAD+ during the base excision repair of chemotherapy-induced DNA damage. With the substantially exhausted metabolic pathways, distinctive metabolic vulnerabilities are established in IDH-mutated malignancies. Effectively targeting these metabolic pathways may induce selective cytotoxicity to cancer cells, but a lesser extent than that occurs in normal somatic cells with an intact metabolic network.
References
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.-H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.-M.; Gallia, G.L.; et al. An Integrated Genomic Analysis of Human Glioblastoma Multiforme. Science 2008, 321, 1807–1812.
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1andIDH2Mutations in Gliomas. N. Engl. J. Med. 2009, 360, 765–773.
- Paschka, P.; Schlenk, R.F.; Gaidzik, V.I.; Habdank, M.; Krönke, J.; Bullinger, L.; Späth, D.; Kayser, S.; Zucknick, M.; Götze, K.; et al. IDH1 and IDH2 Mutations Are Frequent Genetic Alterations in Acute Myeloid Leukemia and Confer Adverse Prognosis in Cytogenetically Normal Acute Myeloid Leukemia With NPM1 Mutation Without FLT3 Internal Tandem Duplication. J. Clin. Oncol. 2010, 28, 3636–3643.
- Mardis, E.R.; Ding, L.; Dooling, D.J.; Larson, D.E.; McLellan, M.D.; Chen, K.; Koboldt, D.C.; Fulton, R.S.; Delehaunty, K.D.; McGrath, S.; et al. Recurring Mutations Found by Sequencing an Acute Myeloid Leukemia Genome. N. Engl. J. Med. 2009, 361, 1058–1066.
- Borger, D.R.; Tanabe, K.K.; Fan, K.C.; Lopez, H.U.; Fantin, V.R.; Straley, K.S.; Schenkein, D.P.; Hezel, A.F.; Ancukiewicz, M.; Liebman, H.M.; et al. Frequent Mutation of Isocitrate Dehydrogenase (IDH)1 and IDH2 in Cholangiocarcinoma Identified Through Broad-Based Tumor Genotyping. Oncologist 2011, 17, 72–79.
- Wang, F.; Travins, J.; DelaBarre, B.; Lacronique, V.; Schalm, S.; Hansen, E.; Straley, K.; Kernytsky, A.; Liu, W.; Gliser, C.; et al. Targeted Inhibition of Mutant IDH2 in Leukemia Cells Induces Cellular Differentiation. Science 2013, 340, 622–626.
- Amary, M.F.; Bácsi, K.; Maggiani, F.; Damato, S.; Halai, D.; Berisha, F.; Pollock, R.; O’Donnell, P.; Grigoriadis, A.; Diss, T.; et al. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J. Pathol. 2011, 224, 334–343.
- Lopez, G.Y.; Reitman, Z.J.; Solomon, D.; Waldman, T.; Bigner, D.D.; McLendon, R.E.; A Rosenberg, S.; Samuels, Y.; Yan, H. IDH1 mutation identified in one human melanoma metastasis, but not correlated with metastases to the brain. Biochem. Biophys. Res. Commun. 2010, 398, 585–587.
- Shibata, T.; Kokubu, A.; Miyamoto, M.; Sasajima, Y.; Yamazaki, N. Mutant IDH1 Confers an in Vivo Growth in a Melanoma Cell Line with BRAF Mutation. Am. J. Pathol. 2011, 178, 1395–1402.
- Yan, H.; Bigner, D.D.; Velculescu, V.E.; Parsons, D.W. Mutant metabolic enzymes are at the origin of gliomas. Cancer Res. 2009, 69, 9157–9159.
- Ward, P.S.; Patel, J.; Wise, D.R.; Abdel-Wahab, O.; Bennett, B.D.; Coller, H.A.; Cross, J.R.; Fantin, V.R.; Hedvat, C.V.; Perl, A.E.; et al. The Common Feature of Leukemia-Associated IDH1 and IDH2 Mutations Is a Neomorphic Enzyme Activity Converting α-Ketoglutarate to 2-Hydroxyglutarate. Cancer Cell 2010, 17, 225–234.
- Dang, L.; Jin, S.; Su, S.M. IDH mutations in glioma and acute myeloid leukemia. Trends Mol. Med. 2010, 16, 387–397.
- Prensner, J.; Chinnaiyan, A.M. Metabolism unhinged: IDH mutations in cancer. Nat. Med. 2011, 17, 291–293.
- Losman, J.-A.; Kaelin, W.G. What a difference a hydroxyl makes: Mutant IDH, (R)-2-hydroxyglutarate, and cancer. Genes Dev. 2013, 27, 836–852.
- Xu, W.; Yang, H.; Liu, Y.; Yang, Y.; Wang, P.; Kim, S.-H.; Ito, S.; Yang, C.; Wang, P.; Xiao, M.-T.; et al. Oncometabolite 2-Hydroxyglutarate Is a Competitive Inhibitor of α-Ketoglutarate-Dependent Dioxygenases. Cancer Cell 2011, 19, 17–30.
- Chowdhury, R.; Yeoh, K.K.; Tian, Y.-M.; Hillringhaus, L.; Bagg, E.; Rose, N.R.; Leung, I.K.H.; Li, X.S.; Woon, E.C.Y.; Yang, M.; et al. The oncometabolite 2-hydroxyglutarate inhibits histone lysine demethylases. EMBO Rep. 2011, 12, 463–469.
- Lu, C.; Ward, P.S.; Kapoor, G.S.; Rohle, D.; Turcan, Ş.; Abdel-Wahab, O.; Edwards, C.R.; Khanin, R.; Figueroa, M.E.; Melnick, A.; et al. IDH mutation impairs histone demethylation and results in a block to cell differentiation. Nature 2012, 483, 474–478.
- Turcan, Ş.; Rohle, D.; Goenka, A.; A Walsh, L.; Fang, F.; Yilmaz, E.; Campos, C.; Fabius, A.W.M.; Lu, C.; Ward, P.S.; et al. IDH1 mutation is sufficient to establish the glioma hypermethylator phenotype. Nature 2012, 483, 479–483.
- Wang, P.; Wu, J.; Ma, S.; Zhang, L.; Yao, J.; Hoadley, K.A.; Wilkerson, M.D.; Perou, C.M.; Guan, K.-L.; Ye, D.; et al. Oncometabolite D-2-Hydroxyglutarate Inhibits ALKBH DNA Repair Enzymes and Sensitizes IDH Mutant Cells to Alkylating Agents. Cell Rep. 2015, 13, 2353–2361.
- Inoue, S.; Li, W.Y.; Tseng, A.; Beerman, I.; Elia, A.J.; Bendall, S.C.; Lemonnier, F.; Kron, K.J.; Cescon, D.W.; Hao, Z.; et al. Mutant IDH1 Downregulates ATM and Alters DNA Repair and Sensitivity to DNA Damage Independent of TET2. Cancer Cell 2016, 30, 337–348.
- Wang, Y.; Wild, A.T.; Turcan, S.; Wu, W.H.; Sigel, C.; Klimstra, D.S.; Ma, X.; Gong, Y.; Holland, E.C.; Huse, J.T.; et al. Targeting therapeutic vulnerabilities with PARP inhibition and radiation in IDH-mutant gliomas and cholangiocarcinomas. Sci. Adv. 2020, 6, eaaz3221.
- Lu, Y.; Kwintkiewicz, J.; Liu, Y.; Tech, K.; Frady, L.N.; Su, Y.-T.; Bautista, W.; Moon, S.I.; Macdonald, J.; Ewend, M.G.; et al. Chemosensitivity of IDH1-Mutated Gliomas Due to an Impairment in PARP1-Mediated DNA Repair. Cancer Res. 2017, 77, 1709–1718.
- Sulkowski, P.L.; Corso, C.D.; Robinson, N.D.; Scanlon, S.E.; Purshouse, K.; Bai, H.; Liu, Y.; Sundaram, R.K.; Hegan, D.C.; Fons, N.R.; et al. 2-Hydroxyglutarate produced by neomorphic IDH mutations suppresses homologous recombination and induces PARP inhibitor sensitivity. Sci. Transl. Med. 2017, 9, eaal2463.
- Shi, J.; Sun, B.; Shi, W.; Zuo, H.; Cui, D.; Ni, L.; Chen, J. Decreasing GSH and increasing ROS in chemosensitivity gliomas with IDH1 mutation. Tumor Boil. 2014, 36, 655–662.
- Tang, X.; Fu, X.; Liu, Y.; Yu, D.; Cai, S.J.; Yang, C. Blockade of Glutathione Metabolism in IDH1-Mutated Glioma. Mol. Cancer Ther. 2019, 19, 221–230.
- Gelman, S.J.; Naser, F.; Mahieu, N.G.; McKenzie, L.D.; Dunn, G.P.; Chheda, M.G.; Patti, G.J. Consumption of NADPH for 2-HG Synthesis Increases Pentose Phosphate Pathway Flux and Sensitizes Cells to Oxidative Stress. Cell Rep. 2018, 22, 512–522.
- Yu, D.; Liu, Y.; Zhou, Y.; Ruiz-Rodado, V.; Larion, M.; Xu, G.; Yang, C. Triptolide suppresses IDH1-mutated malignancy via Nrf2-driven glutathione metabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 9964–9972.
- Liu, Y.; Lu, Y.; Celiku, O.; Li, A.; Wu, Q.; Zhou, Y.; Yang, C. Targeting IDH1-Mutated Malignancies with NRF2 Blockade. J. Natl. Cancer Inst. 2019, 111, 1033–1041.
- McBrayer, S.K.; Mayers, J.R.; DiNatale, G.J.; Shi, D.D.; Khanal, J.; Chakraborty, A.A.; Sarosiek, K.; Briggs, K.J.; Robbins, A.K.; Sewastianik, T.; et al. Transaminase Inhibition by 2-Hydroxyglutarate Impairs Glutamate Biosynthesis and Redox Homeostasis in Glioma. Cell 2018, 175, 101–116.e25.
- Wen, P.Y.; Huse, J.T. 2016 World Health Organization Classification of Central Nervous System Tumors. Contin. Lifelong Learn. Neurol. 2017, 23, 1531–1547.
- Riemenschneider, M.J.; Jeuken, J.W.M.; Wesseling, P.; Reifenberger, G. Molecular diagnostics of gliomas: State of the art. Acta Neuropathol. 2010, 120, 567–584.
- Chen, R.; Smith-Cohn, M.; Cohen, A.L.; Colman, H. Glioma Subclassifications and Their Clinical Significance. Neurotherapeutics 2017, 14, 284–297.
- Jaunmuktane, Z.; Capper, D.; Jones, D.T.W.; Schrimpf, D.; Sill, M.; Dutt, M.; Suraweera, N.; Pfister, S.M.; Von Deimling, A.; Brandner, S. Methylation array profiling of adult brain tumours: Diagnostic outcomes in a large, single centre. Acta Neuropathol. Commun. 2019, 7, 24.
- Andronesi, O.C.; Rapalino, O.; Gerstner, E.; Chi, A.; Batchelor, T.T.; Cahill, D.P.; Sorensen, A.G.; Rosen, B.R. Detection of oncogenic IDH1 mutations using magnetic resonance spectroscopy of 2-hydroxyglutarate. J. Clin. Investig. 2013, 123, 3659–3663.
- Andronesi, O.C.; Kim, G.S.; Gerstner, E.; Batchelor, T.; Tzika, A.A.; Fantin, V.R.; Heiden, M.G.V.; Sorensen, A.G. Detection of 2-Hydroxyglutarate in IDH-Mutated Glioma Patients by In Vivo Spectral-Editing and 2D Correlation Magnetic Resonance Spectroscopy. Sci. Transl. Med. 2012, 4, 116ra4.
- Pope, W.B.; Prins, R.M.; Thomas, M.A.; Nagarajan, R.; Yen, K.E.; Bittinger, M.A.; Salamon, N.; Chou, A.P.; Yong, W.H.; Soto, H.; et al. Non-invasive detection of 2-hydroxyglutarate and other metabolites in IDH1 mutant glioma patients using magnetic resonance spectroscopy. Neuro-Oncology 2011, 107, 197–205.
- Natsumeda, M.; Igarashi, H.; Nomura, T.; Ogura, R.; Tsukamoto, Y.; Kobayashi, T.; Aoki, H.; Okamoto, K.; Kakita, A.; Takahashi, H.; et al. Accumulation of 2-hydroxyglutarate in gliomas correlates with survival: A study by 3.0-tesla magnetic resonance spectroscopy. Acta Neuropathol. Commun. 2014, 2, 158.
- Lazovic, J.; Soto, H.; Piccioni, D.E.; Ru, J.R.L.; Li, S.; Mirsadraei, L.; Yong, W.; Prins, R.M.; Liau, L.M.; Ellingson, B.M.; et al. Detection of 2-hydroxyglutaric acid in vivo by proton magnetic resonance spectroscopy in U87 glioma cells overexpressing isocitrate dehydrogenase–1 mutation. Neuro-Oncology 2012, 14, 1465–1472.
- Heo, H.; Kim, S.; Lee, H.H.; Cho, H.R.; Xu, W.J.; Lee, S.-H.; Park, C.K.; Park, S.; Choi, S.H.; Kim, H. On the Utility of Short Echo Time (TE) Single Voxel 1H–MRS in Non–Invasive Detection of 2–Hydroxyglutarate (2HG); Challenges and Potential Improvement Illustrated with Animal Models Using MRUI and LCModel. PLoS ONE 2016, 11, e0147794.
- Choi, C.; Ganji, S.K.; DeBerardinis, R.J.; Hatanpaa, K.J.; Rakheja, D.; Kovacs, Z.; Yang, X.-L.; Mashimo, T.; Raisanen, J.M.; Marin-Valencia, I.; et al. 2-hydroxyglutarate detection by magnetic resonance spectroscopy in IDH-mutated patients with gliomas. Nat. Med. 2012, 18, 624–629.
- Choi, C.; Ganji, S.; Hulsey, K.; Madan, A.; Kovacs, Z.; Dimitrov, I.; Zhang, S.; Pichumani, K.; Mendelsohn, D.; Mickey, B.; et al. A comparative study of short- and long-TE ¹H MRS at 3 T for in vivo detection of 2-hydroxyglutarate in brain tumors. NMR Biomed. 2013, 26, 1242–1250.
- De La Fuente, M.I.; Young, R.J.; Rubel, J.; Rosenblum, M.; Tisnado, J.; Briggs, S.; Pérez, J.A.; Cross, J.R.; Campos, C.; Straley, K.; et al. Integration of 2-hydroxyglutarate-proton magnetic resonance spectroscopy into clinical practice for disease monitoring in isocitrate dehydrogenase-mutant glioma. Neuro-Oncology 2015, 18, 283–290.
- Emir, U.E.; Larkin, S.J.; De Pennington, N.; Voets, N.L.; Plaha, P.; Stacey, R.; Al-Qahtani, K.; Mccullagh, J.S.; Schofield, C.J.; Clare, S.; et al. Noninvasive Quantification of 2-Hydroxyglutarate in Human Gliomas with IDH1 and IDH2 Mutations. Cancer Res. 2015, 76, 43–49.
- Killela, P.J.; Pirozzi, C.J.; Healy, P.; Reitman, Z.J.; Lipp, E.S.; Rasheed, B.A.; Yang, R.; Diplas, B.; Wang, Z.; Greer, P.K.; et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget 2014, 5, 1515–1525.
- Sanson, M.; Marie, Y.; Paris, S.; Idbaih, A.; Laffaire, J.; Ducray, F.; El Hallani, S.; Boisselier, B.; Mokhtari, K.; Hoang-Xuan, K.; et al. Isocitrate Dehydrogenase 1 Codon 132 Mutation Is an Important Prognostic Biomarker in Gliomas. J. Clin. Oncol. 2009, 27, 4150–4154.
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The Somatic Genomic Landscape of Glioblastoma. Cell 2013, 155, 462–477.
- Nobusawa, S.; Watanabe, T.; Kleihues, P.; Ohgaki, H. IDH1 Mutations as Molecular Signature and Predictive Factor of Secondary Glioblastomas. Clin. Cancer Res. 2009, 15, 6002–6007.
- Toedt, G.; Barbus, S.; Wolter, M.; Felsberg, J.; Tews, B.; Blond, F.; Sabel, M.; Hofmann, S.; Becker, N.; Hartmann, C.; et al. Molecular signatures classify astrocytic gliomas byIDH1mutation status. Int. J. Cancer 2010, 128, 1095–1103.
- Weller, M.; Felsberg, J.; Hartmann, C.; Berger, H.; Steinbach, J.P.; Schramm, J.; Westphal, M.; Schackert, G.; Simon, M.; Tonn, J.C.; et al. Molecular Predictors of Progression-Free and Overall Survival in Patients With Newly Diagnosed Glioblastoma: A Prospective Translational Study of the German Glioma Network. J. Clin. Oncol. 2009, 27, 5743–5750.
- Lai, A.; Kharbanda, S.; Pope, W.B.; Tran, A.; Solis, O.E.; Peale, F.; Forrest, W.F.; Pujara, K.; Carrillo, J.A.; Pandita, A.; et al. Evidence for Sequenced Molecular Evolution of IDH1 Mutant Glioblastoma From a Distinct Cell of Origin. J. Clin. Oncol. 2011, 29, 4482–4490.
- Balss, J.; Meyer, J.; Mueller, W.; Korshunov, A.; Hartmann, C.; Von Deimling, A. Analysis of the IDH1 codon 132 mutation in brain tumors. Acta Neuropathol. 2008, 116, 597–602.
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Skanderup, A.J.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data: Figure 1. Cancer Discov. 2012, 2, 401–404.
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Skanderup, A.J.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1.
- Chaichana, K.L.; Parker, S.L.; Olivi, A.; Quinones-Hinojosa, A. Long-term seizure outcomes in adult patients undergoing primary resection of malignant brain astrocytomas. J. Neurosurg. 2009, 111, 282–292.
- Chang, E.F.; Potts, M.B.; Keles, G.E.; Lamborn, K.R.; Chang, S.M.; Barbaro, N.M.; Berger, M.S. Seizure characteristics and control following resection in 332 patients with low-grade gliomas. J. Neurosurg. 2008, 108, 227–235.
- Kerkhof, M.; Vecht, C.J. Seizure characteristics and prognostic factors of gliomas. Epilepsia 2013, 54, 12–17.
- Van Breemen, M.S.; Wilms, E.B.; Vecht, C.J. Epilepsy in patients with brain tumours: Epidemiology, mechanisms, and management. Lancet Neurol. 2007, 6, 421–430.
- Klein, M.; Engelberts, N.H.J.; Van Der Ploeg, H.M.; Trenité, D.G.A.K.-N.; Aaronson, N.K.; Taphoorn, M.J.B.; Baaijen, H.; Vandertop, W.P.; Müller, M.; Postma, T.J.; et al. Epilepsy in low-grade gliomas: The impact on cognitive function and quality of life. Ann. Neurol. 2003, 54, 514–520.
- Stockhammer, F.; Misch, M.; Helms, H.-J.; Lengler, U.; Prall, F.; Von Deimling, A.; Hartmann, C. IDH1/2 mutations in WHO grade II astrocytomas associated with localization and seizure as the initial symptom. Seizure 2012, 21, 194–197.
- Liubinas, S.V.; D’Abaco, G.M.; Moffat, B.M.; Gonzales, M.; Feleppa, F.; Nowell, C.J.; Gorelik, A.; Drummond, K.J.; O’Brien, T.J.; Kaye, A.H.; et al. IDH1 mutation is associated with seizures and protoplasmic subtype in patients with low-grade gliomas. Epilepsia 2014, 55, 1438–1443.
- Yang, Y.; Qing, M.; Wang, X.; Liu, Y.; Mao, Y.; Zhou, Q.; Luo, J. An analysis of 170 glioma patients and systematic review to investigate the association between IDH-1 mutations and preoperative glioma-related epilepsy. J. Clin. Neurosci. 2016, 31, 56–62.
- Chen, H.; Judkins, J.; Thomas, C.; Wu, M.; Khoury, L.; Benjamin, C.; Pacione, D.; Golfinos, J.G.; Kumthekar, P.; Ghamsari, F.; et al. Mutant IDH1 and seizures in patients with glioma. Neurology 2017, 88, 1805–1813.
- Toledo, M.; Sarria-Estrada, S.; Quintana, M.; Maldonado, X.; Martínez-Ricarte, F.; Rodon, J.; Auger, C.; Aizpurua, M.; Salas-Puig, J.; Santamarina, E.; et al. Epileptic features and survival in glioblastomas presenting with seizures. Epilepsy Res. 2017, 130, 1–6.
- Moussawi, K.; Riegel, A.; Nair, S.; Kalivas, P.W. Extracellular Glutamate: Functional Compartments Operate in Different Concentration Ranges. Front. Syst. Neurosci. 2011, 5, 94.
- Elkhaled, A.; Jalbert, L.; Constantin, A.; Yoshihara, H.A.I.; Phillips, J.J.; Molinaro, A.M.; Chang, S.M.; Nelson, S.J. Characterization of metabolites in infiltrating gliomas usingex vivo1H high-resolution magic angle spinning spectroscopy. NMR Biomed. 2014, 27, 578–593.
- Vaarmann, A.; Kovac, S.; Holmstrom, K.; Gandhi, S.; Abramov, A.Y. Dopamine protects neurons against glutamate-induced excitotoxicity. Cell Death Dis. 2013, 4, e455.
- Rohle, D.; Popovici-Muller, J.; Palaskas, N.; Turcan, Ş.; Grommes, C.; Campos, C.; Tsoi, J.; Clark, O.; Oldrini, B.; Komisopoulou, E.; et al. An Inhibitor of Mutant IDH1 Delays Growth and Promotes Differentiation of Glioma Cells. Science 2013, 340, 626–630.
- Hartmann, C.; Hentschel, B.; Wick, W.; Capper, D.; Felsberg, J.; Simon, M.; Westphal, M.; Schackert, G.; Meyermann, R.; Pietsch, T.; et al. Patients with IDH1 wild type anaplastic astrocytomas exhibit worse prognosis than IDH1-mutated glioblastomas, and IDH1 mutation status accounts for the unfavorable prognostic effect of higher age: Implications for classification of gliomas. Acta Neuropathol. 2010, 120, 707–718.
- Houillier, C.; Wang, X.; Kaloshi, G.; Mokhtari, K.; Guillevin, R.; Laffaire, J.; Paris, S.; Boisselier, B.; Idbaih, A.; Laigle-Donadey, F.; et al. IDH1 or IDH2 mutations predict longer survival and response to temozolomide in low-grade gliomas. Neurology 2010, 75, 1560–1566.
- Hartmann, C.; Hentschel, B.; Tatagiba, M.; Schramm, J.; Schnell, O.; Seidel, C.; Stein, R.; Reifenberger, G.; Pietsch, T.; Von Deimling, A.; et al. Molecular Markers in Low-Grade Gliomas: Predictive or Prognostic? Clin. Cancer Res. 2011, 17, 4588–4599.
- Bent, M.J.V.D.; Dubbink, H.J.; Marie, Y.; Brandes, A.A.; Taphoorn, M.J.; Wesseling, P.; Frenay, M.; Tijssen, C.C.; Lacombe, D.; Idbaih, A.; et al. IDH1 and IDH2 Mutations Are Prognostic but not Predictive for Outcome in Anaplastic Oligodendroglial Tumors: A Report of the European Organization for Research and Treatment of Cancer Brain Tumor Group. Clin. Cancer Res. 2010, 16, 1597–1604.
- Popovici-Muller, J.; Saunders, J.O.; Salituro, F.G.; Travins, J.M.; Yan, S.; Zhao, F.; Gross, S.; Dang, L.; Yen, K.E.; Yang, H.; et al. Discovery of the First Potent Inhibitors of Mutant IDH1 That Lower Tumor 2-HG in Vivo. ACS Med. Chem. Lett. 2012, 3, 850–855.
- Dinardo, C.; Stein, E.M.; De Botton, S.; Roboz, G.J.; Altman, J.K.; Mims, A.S.; Swords, R.; Collins, R.H.; Mannis, G.N.; Pollyea, D.A.; et al. Durable Remissions with Ivosidenib inIDH1-Mutated Relapsed or Refractory AML. N. Engl. J. Med. 2018, 378, 2386–2398.
- Konteatis, Z.; Artin, E.; Nicolay, B.; Straley, K.; Padyana, A.K.; Jin, L.; Chen, Y.; Narayaraswamy, R.; Tong, S.; Wang, F.; et al. Vorasidenib (AG-881): A First-in-Class, Brain-Penetrant Dual Inhibitor of Mutant IDH1 and 2 for Treatment of Glioma. ACS Med. Chem. Lett. 2020, 11, 101–107.
- Fan, B.; Mellinghoff, I.K.; Wen, P.Y.; Lowery, M.A.; Goyal, L.; Tap, W.D.; Pandya, S.S.; Manyak, E.; Jiang, L.; Liu, G.; et al. Clinical pharmacokinetics and pharmacodynamics of ivosidenib, an oral, targeted inhibitor of mutant IDH1, in patients with advanced solid tumors. Investig. New Drugs 2019, 38, 433–444.
- Popovici-Muller, J.; Lemieux, R.M.; Artin, E.; Saunders, J.O.; Salituro, F.G.; Travins, J.; Cianchetta, G.; Cai, Z.; Zhou, D.; Cui, D.; et al. Discovery of AG-120 (Ivosidenib): A First-in-Class Mutant IDH1 Inhibitor for the Treatment of IDH1 Mutant Cancers. ACS Med. Chem. Lett. 2018, 9, 300–305.
- Mellinghoff, I.K.; Ellingson, B.M.; Touat, M.; Maher, E.; De La Fuente, M.I.; Holdhoff, M.; Cote, G.M.; Burris, H.; Janku, F.; Young, R.J.; et al. Ivosidenib in Isocitrate Dehydrogenase 1–Mutated Advanced Glioma. J. Clin. Oncol. 2020, JCO1903327.
- Molenaar, R.J.; Radivoyevitch, T.; Nagata, Y.; Khurshed, M.; Przychodzen, B.; Makishima, H.; Xu, M.; Bleeker, F.E.; Wilmink, J.W.; Carraway, H.E.; et al. IDH1/2Mutations Sensitize Acute Myeloid Leukemia to PARP Inhibition and This Is Reversed by IDH1/2-Mutant Inhibitors. Clin. Cancer Res. 2018, 24, 1705–1715.
- Flavahan, W.A.; Drier, Y.; Liau, B.B.; Gillespie, S.M.; Venteicher, A.S.; Stemmer-Rachamimov, A.O.; Suvà, M.L.; Bernstein, B.E. Insulator dysfunction and oncogene activation in IDH mutant gliomas. Nature 2015, 529, 110–114.
- Borodovsky, A.; Salmasi, V.; Turcan, Ş.; Fabius, A.W.M.; Baia, G.S.; Eberhart, C.G.; Weingart, J.D.; Gallia, G.L.; Baylin, S.B.; Chan, T.A.; et al. 5-azacytidine reduces methylation, promotes differentiation and induces tumor regression in a patient-derived IDH1 mutant glioma xenograft. Oncotarget 2013, 4, 1737–1747.
- Turcan, Ş.; Fabius, A.W.M.; Borodovsky, A.; Pedraza, A.; Brennan, C.W.; Huse, J.; Viale, A.; Riggins, G.J.; Chan, T.A. Efficient induction of differentiation and growth inhibition in IDH1 mutant glioma cells by the DNMT Inhibitor Decitabine. Oncotarget 2013, 4, 1729–1736.
- Fathi, A.T.; Abdel-Wahab, O. Mutations in Epigenetic Modifiers in Myeloid Malignancies and the Prospect of Novel Epigenetic-Targeted Therapy. Adv. Hematol. 2012, 2012, 1–12.
- Gursoy-Yuzugullu, O.; Carman, C.; Serafim, R.B.; Myronakis, M.; Valente, V.; Price, B.D. Epigenetic therapy with inhibitors of histone methylation suppresses DNA damage signaling and increases glioma cell radiosensitivity. Oncotarget 2017, 8, 24518–24532.
- Bent, M.J.V.D.; Mellinghoff, I.K.; Bindra, R.S.; Binja, R. Gray Areas in the Gray Matter: IDH1/2 Mutations in Glioma. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, 96–103.
- Esteller, M.; García-Foncillas, J.; Andion, E.; Goodman, S.N.; Hidalgo, O.F.; Vanaclocha, V.; Baylin, S.B.; Herman, J.G. Inactivation of the DNA-Repair GeneMGMTand the Clinical Response of Gliomas to Alkylating Agents. N. Engl. J. Med. 2000, 343, 1350–1354.
- Hegi, M.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; De Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMTGene Silencing and Benefit from Temozolomide in Glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003.
- Lu, Y.; Liu, Y.; Pang, Y.; Pacak, K.; Yang, C. Double-barreled gun: Combination of PARP inhibitor with conventional chemotherapy. Pharmacol. Ther. 2018, 188, 168–175.
- Cai, S.J.; Liu, Y.; Han, S.; Yang, C. Brusatol, an NRF2 inhibitor for future cancer therapeutic. Cell Biosci. 2019, 9, 45.
- Seltzer, M.J.; Bennett, B.D.; Joshi, A.D.; Gao, P.; Thomas, A.G.; Ferraris, D.; Tsukamoto, T.; Rojas, C.J.; Slusher, B.S.; Rabinowitz, J.D.; et al. Inhibition of glutaminase preferentially slows growth of glioma cells with mutant IDH1. Cancer Res. 2010, 70, 8981–8987.
- Emadi, A.; Jun, S.A.; Tsukamoto, T.; Fathi, A.T.; Minden, M.D.; Dang, C.V. Inhibition of glutaminase selectively suppresses the growth of primary acute myeloid leukemia cells with IDH mutations. Exp. Hematol. 2014, 42, 247–251.
- Matre, P.; Velez, J.; Jácamo, R.; Qi, Y.; Su, X.; Cai, T.; Chan, S.M.; Lodi, A.; Sweeney, S.R.; Ma, H.; et al. Inhibiting glutaminase in acute myeloid leukemia: Metabolic dependency of selected AML subtypes. Oncotarget 2016, 7, 79722–79735.
- Tateishi, K.; Wakimoto, H.; Iafrate, A.J.; Tanaka, S.; Loebel, F.; Lelic, N.; Wiederschain, D.; Bedel, O.; Deng, G.; Zhang, B.; et al. Extreme Vulnerability of IDH1 Mutant Cancers to NAD+ Depletion. Cancer Cell 2015, 28, 773–784.
- Tateishi, K.; Higuchi, F.; Miller, J.J.; Koerner, M.V.; Lelic, N.; Shankar, G.M.; Tanaka, S.; Fisher, D.E.; Batchelor, T.T.; Iafrate, A.J.; et al. The Alkylating Chemotherapeutic Temozolomide Induces Metabolic Stress in IDH1-Mutant Cancers and Potentiates NAD+ Depletion-Mediated Cytotoxicity. Cancer Res. 2017, 77, 4102–4115.
More
Information
Subjects:
Medicine, General & Internal
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.2K
Revisions:
2 times
(View History)
Update Date:
25 Jun 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No