 +1 credit
+1 credit
Video Upload Options
Plant enzymes are superoxide dismutase (SOD), catalase (CAT), peroxidase (POX), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione S-transferases (GST), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), and dehydroascorbate reductase (DHAR), which work as part of the antioxidant defence system. These enzymes together form a complex set of mechanisms to minimise, buffer, and scavenge the reactive oxygen species (ROS) efficiently.
1. Introduction
Plants are immobile; they cannot escape from biotic, (i.e., pathogens, parasites, grazing) and abiotic (such as drought, flooding, salinity, low-high temperatures, ultraviolet radiation, nutrient deficiency, heavy metal (HM) toxicity) stresses. Plant growth, development and productivity are influenced by a variety of environmental stresses. These stresses often perturb the homeostasis and ion distribution in plant cells and induces osmotic stress, leading to an enhancement in the accumulation of reactive oxygen species (ROS) [1]. The production and accumulation of ROS in the plants result in severe destruction of cell organelles and functions cause membrane peroxidation, leading to damage in the cell membrane, degradation of biological macromolecules and ultimately cell death. The ability of plants to scavenge the toxic effects of ROS seems to be the most important determinant for their tolerance to different stresses. Antioxidants are the first line of defence against the damages caused by free radicals and are critical for the optimum health of plant cells [2][3][4][5]. Plant antioxidants play a significant role in assisting plant development through a wide variety of mechanisms and functions.
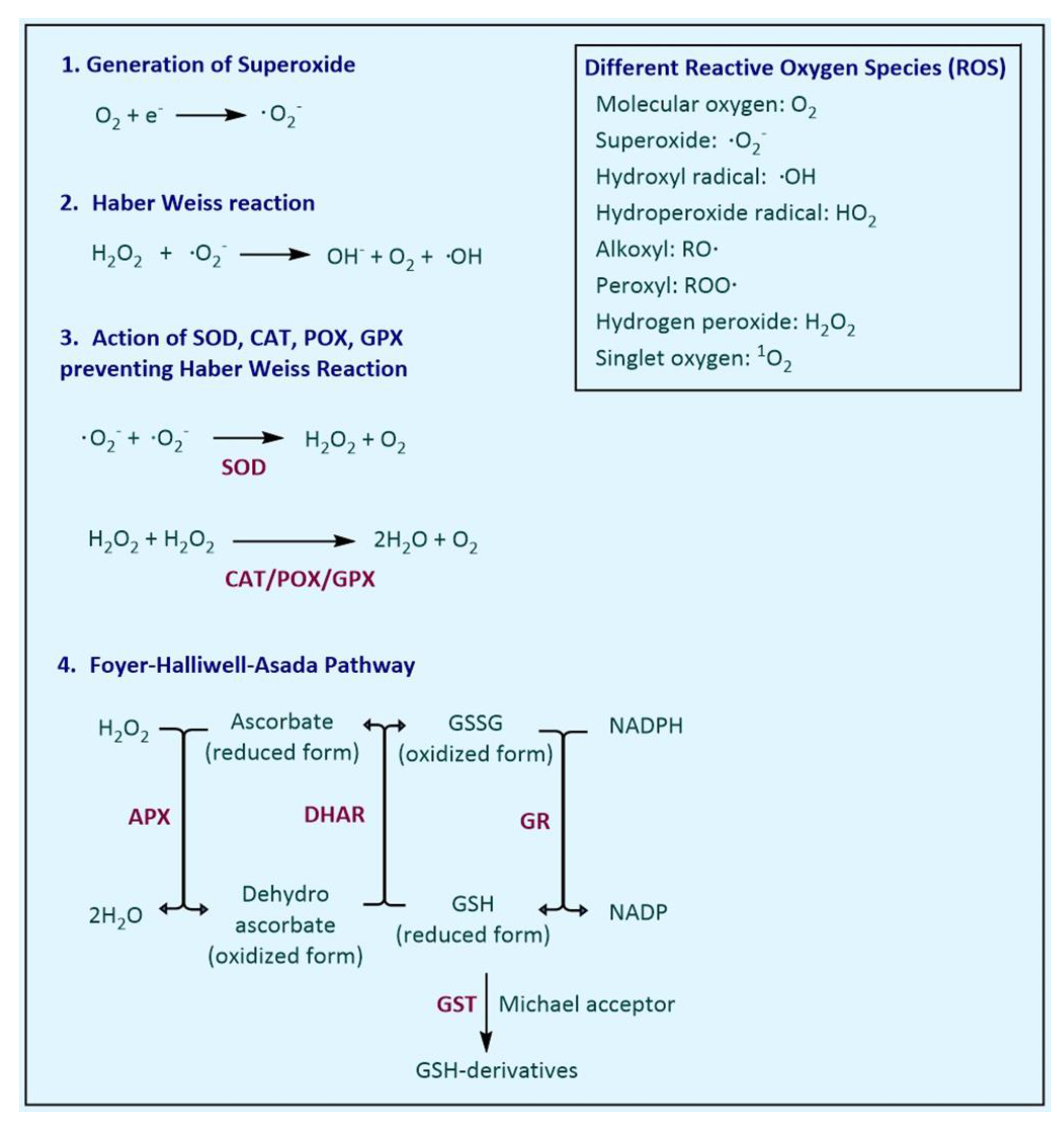
There are several antioxidant enzymes associated with ROS scavenging in plants, and the synthesis of these enzymes is known to be enhanced during the exposure to oxidative stresses [6]. The ROS comprise free radicals, such as superoxide radicals (•O2−), hydroxyl radicals (•OH), perhydroxyl radicals (HO2−) and alkoxy radicals, and non-radical forms, i.e., hydrogen peroxide (H2O2) and singlet oxygen (1O2), present in the intra- and extra-cellular locations of the plant. Superoxide radicals (•O2−) can be generated by a single electron transfer (e−) to dioxygen (O2).
Chloroplasts and mitochondria are the two main sites for the generation of ROS. The photosynthetic electron transport system (ETS) is one of the important sites for the generation of ROS, and this site has the potential to generate singlet oxygen 1O and superoxide (•O2−). Plant mitochondria differ from animal as it possesses O2 and carbohydrate-rich environment [7], and also being associated with photorespiration. The mitochondrial ETC (mtETC) is also a source of generation of ROS as it houses sufficiently energised electrons to reduce the O2. The major parts of the mtETC responsible for producing ROS are Complex I and Complex III [8]. Other sources of ROS production in the mitochondria are from the different enzymes present in the matrix. There are other sites as well for the generation of ROS, such as the endoplasmic reticulum, cell membrane, cell wall and apoplast.
Evolution has equipped plants with a wide range of defence measures, which include various enzymatic strategies to scavenge free ROS in plant cells [9][10]. The tolerance mechanisms in stressed plant include a number of physio-biochemical strategies, which includes many enzymatic components, such as superoxide dismutase (SOD), catalase (CAT), peroxidases (POX), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione S-transferases (GST), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR) and dehydroascorbate reductase (DHAR), and non-enzymatic components, such as ascorbic acid (AA), glutathione (GSH), phenolic compounds, alkaloids, flavonoids, carotenoids, free amino acids and α-tocopherols [11][12][13]. However, in the present review, we have exclusively focussed on the role and mechanisms of enzymatic components in the plant to scavenge the ROS and to cope with the stress conditions. These enzymes are selected on the basis of majority of the research reports available and with their proven utility in transgenic plants to cope with the stress conditions (Table 1). During stresses, SOD catalyses the removal of •O2− by dismutating it into O2 and H2O2, CAT converts the H2O2 into water and molecular oxygen (O2) and POX works in the extra-cellular space for scavenging H2O2. Plant GPX catalyses the reduction of H2O2 and HO2 to water and lipid alcohols, respectively, using thioredoxin as an electron donor. Glutathione reductase catalyses the reduction of oxidised glutathione (GSSG; dimeric) to reduced glutathione (GSH; monomeric) and APX utilises ascorbate as specific electron donor to scavenge H2O2 to water.
These enzymes not only protect various components of the cells from damages, but also play an important role in plant growth and development by modulating cellular–sub-cellular processes such as mitosis [14], cell elongation [15], senescence [16] and cell death [17], and are also involved in a wide range of processes, such as cell differentiation [18], cell growth/division [19], regulation of senescence and sulphate transport [20][21], detoxification of xenobiotics [22], conjugation of metabolites [23], regulation of enzymatic activities [24], synthesis of proteins and nucleotides [25][26], phytochelatins [27] and expression of stress responsive genes [28]. The antioxidant defence system protects the unsaturated membrane lipids, nucleic acids, enzymes and other cellular structures from the negative impacts of free radicals [29]. Therefore, the antioxidant defence system of plants has been attracting considerable interest of the scientific community [29][30].
2. Enzymatic Antioxidant Defence Systems in Plants
The antioxidant defence system in the plant comprises several different enzymes. They are mainly involved in either preventing the Haber-Weiss reaction (Figure 1) or the Foyer–Halliwell–Asada pathway, which reduces the H2O2 and utilises the reducing potential of NADPH.
3. Applications of Antioxidant Enzymes in Developing Stress-Tolerant Transgenic Plants
Sustainable agriculture production is a key factor in ensuring global food security. However, there are multiple stress conditions that influences the crop growth and yield. In order to overcome these stress conditions, developing stress-tolerant plants is an important step. Understanding the role of individual gene under the influence of different stress condition can be useful in developing stress-tolerant plants. The overexpression of different genes of different antioxidant enzymes has resulted in the increase in tolerance in transgenic plants to various environmental stress conditions. Several stress-tolerant genetically engineered plants have been developed in the recent past, and the significant findings of these research reports are briefly described in Table 1. The majority of these studies focussed on abiotic stress caused due to salinity, heat, chilling, drought, flood and HM, but very few reports are available on understanding the role of these enzymes to cope with biotic stress. Furthermore, in all of these studies, the gene of the antioxidant enzyme is overexpressed under a strong promoter in transgenic lines, thereby increasing the tolerance potential of the plant to stress condition. Thus, these findings are crucial for developing stress-resistant plants, and the knowledge gained will be helpful for sustained growth and productivity of various crops in variable environmental conditions.
| S.No. | Transgenic Plant(s) | Gene(s)/Source | Stress Condition | Significant Finding(s) | Reference |
|---|---|---|---|---|---|
| 1. | Transgenic S. lycopersicum | FeSOD gene from Arabidopsis | Salt stress | Overexpression of antioxidant enzymes significantly mitigates the harmful effects of salt stress on cytoskeleton structural organisation in roots of the transgenic line cells. | [172] |
| 2. | Transgenic S. tuberosum | Cu-ZnSOD (StSOD1 gene overexpressed under CaMV 35S promoter) |
Low temperature | Activity of SOD is 1.38-fold higher compared to non-transgenic lines. Furthermore, the activity of POX and CAT were also enhanced in transgenic line, signifying the fact that increasing the activity of one antioxidant enzyme can influence the activity of other defence enzymes via cross-talk. | [173] |
| 3. | Transgenic Citrus sps | CsPIF8 influencing SOD gene expression | Low temperature | Phytochrome-interacting transcription factor CsPIF8 positively regulate CsSOD expression in citrus, highlighting the cross-talk between phytochrome genes and antioxidant enzymes. In this study, it is found that CsPIF8 directly bound to the E-box (CANNTG) of CsSOD promoter and activated the promoter of CsSOD. | [50] |
| 4. | Transgenic Arabidopsis | CmSOD gene (from winter squash; Cucurbita moschata) and AtSOD gene (from Arabidopsis) under a ubiquitin promoter | Low temperature | Increased resistance to chilling and less oxidative injury in transgenic lines than wild type, indicating that the overexpression of AtSOD and CmSOD led to higher SOD activity in Arabidopsis-enhanced chilling tolerance by eliminating •O2−. Furthermore, the activity of SOD in transgenic lines is influenced by ABA, indicating the role of plant hormone in the cross-talk with enzymes of the antioxidant defence system. | [174] |
| 5. | Transgenic Arabidopsis | Cu-Zn SOD gene (SaCu/Zn SOD), from Sedum alfredii | Oxidative stress due to Cadmium | Cadmium stress induces the production of ROS, leading to oxidative stress. Cd-hyperaccumulator plant S. alfredii is used as a source of SOD gene, resulting in enhanced antioxidative defence capacity in transgenic Arabidopsis plants. The SaCu/Zn SOD is implicated as being responsible for conferring Cd tolerance. | [175] |
| 6. | Transgenic tobacco | Cu/Zn-SOD gene, SiCSD from Saussurea involucrata | Drought, cold and oxidative stress | Higher activities of SODs, CAT and APX are reported in transgenic lines, and SOD is found as a positive regulator in drought and cold stress by reducing oxidant injury. | [176] |
| 7. | Transgenic C. grandis | The basic helix-loop-helix (bHLH) family of transcription factors (PtrbHLH) from Poncirus trifoliata | Low temperature | Transgenic plant was found to exhibit lower electrolyte leakage and malondialdehyde content after chilling stress, lower ROS levels and elevated activity of antioxidant enzymes, including CAT, POX and SOD. Interestingly, PtrbHLH was found to bind to the promoter and activate the PtrCAT gene, thereby implicated as regulating the CAT gene activity. | [177] |
| 8. | Manihot esculenta | SOD (MeCu/ZnSOD) and catalase (MeCAT1) | Biotic stress (Mite Tetranychus cinnabarinus) |
The transgenic approach led to mite-resistant traits, as survival, reproduction and development of T. cinnabarinus feeding on transgenic cassava is significantly inhibited. Furthermore, the activities of SOD and CAT in transgenic cassava plants damaged by T. cinnabarinus significantly increased. This study highlights the role of antioxidant enzymes in developing pest resistant crops. | [178] |
| 9. | Transgenic Ipomoea batatas | Peroxidase gene swpa4 in I. batatas | Salt stress | Overexpressing the swpa4 gene under CaMV 35S promoter led to 3- to 13-fold higher expression in transgenic sweet potato. Transgenic plants also showed increased tolerance to salinity conditions, with 13–26% less damage than control plants. Furthermore, photosynthetic capacity and total chlorophyll contents were less severely impacted in transgenic plants. | [179] |
| 10. | Transgenic Arabidopsis | Glutathione peroxidase-like 5 gene (AtGPXL5) from Arabidopsis | Salt stress | Constitutive overexpression of AtGPXL5 led to an increase in gene expression by 17–24 times in 6-week-old plants. It results in an increase in GSH pool and more negative redox potential than wild type and increased salt tolerance. | [91] |
| 11. | Transgenic Arabidopsis | AtGR1 encoding glutathione reductase (GR) from Arabidopsis | Aluminium toxicity | The overexpression of AtGR1 led to a higher GSH pool and improved ratio of GSH/GSSG, and increased aluminium tolerance, with better root growth in comparison to the wild type under aluminium stress. Increased GSH levels were found to increase the capacity of RCS detoxification, which indicates that GR overexpression contributes to the mitigating of not only ROS, but also RCS. | [180] |
| 12. | Transgenic O. sativa | OsGSTU5 (a tau class GST in O. sativa) | Biotic stress | Overexpression of OsGSTU5 provided tolerance against sheath blight disease, caused by Rhizoctonia solani. | [181] |
| 13. | Transgenic Arabidopsis | Glutathione S-transferase from Thermosynechococcus elongatus BP-1 (TeGST) | Thiocyanate (SCN−) stress | Overexpression of TeGST in transgenic plant increased the tolerance to thiocyanate (SCN-) up to 5 mmol L−1. This approach was found to be potentially effective to enhance the phytoremediation of environmental thiocyanates. | [182] |
| 14. | Transgenic Arabidopsis | Ascorbate peroxidase (AgAPX1) from Apium graveolens | Drought tolerance | Overexpression of the AgAPX1 gene enhanced ascorbate content, antioxidant capacity and drought resistance. Furthermore, increased antioxidant capacity does not affect the growth parameters of the plant much, as a comparatively smaller decrease in the net photosynthetic rate is observed, and a high survival rate of transgenic Arabidopsis lines after drought is reported. | [43] |
| 15. | Transgenic Arabidopsis | Ascorbate peroxidase gene (DaAPX) from Dioscorea alata | Flood/Chilling stress | This study reports the effect of different types of stress on the expression of DaAPX. Yam variety Minghuai 1 (MH1), when exposed to a flood situation, showed an increase in the expression of DaAPX; however, chilling stress did not influence the expression profile of DaAPX, thereby making this variety sensitive to chilling stress. However, overexpression of DaAPX in Arabidopsis led to increased tolerance towards several abiotic stress, including flooding and chilling. | [152] |
| 16. | Transgenic Brassica juncea | Ascorbate peroxidase gene (Apx1) from Arabidopsis | Salt stress | Overexpression of cytosolic AtApx1 gene increased salinity stress tolerance in B. juncea. APX, along with higher activity of other enzymes such as GPX, CAT and POX, maintains the ROS homeostasis and provides tolerance to the cell, greater proline accumulation, increased chlorophyll stability index and lower chlorophyll a/b ratio. | [150] |
| 17. | Transgenic Nicotiana tabacum | Monodehydroascorbate reductase from S. lycopersicum (SlMDHAR) | Salt stress | Overexpression of SlMDHAR in transgenic tobacco is found to increase salt stress tolerance and NO accumulation and the S-nitrosyalted SlMDHAR levels were found to be higher in transgenic tobacco. Results suggested that SlMDHAR confers salt stress tolerance by probably involving the S-nitrosylation (post-translational modification of cysteine thiol by nitric oxide group) of MDHAR. | [183] |
| 18. | Transgenic Arabidopsis | Monodehydroascorbate reductase (BvM14-MDHAR) from B. vulgaris | Salt stress | The MDHAR gene is constitutively expressed in Arabidopsis, resulting in an enhanced salt stress tolerance phenotype, with higher AsA/DHA levels than wild-type. In addition, the overexpression seedlings showed higher activities of MDHAR and DHAR and decreased cell membrane damage. | [184] |
| 19. | Transgenic Arabidopsis | DHAR (AcDHAR1 and AcDHAR2) from Actinidia chinensis (kiwi fruit) | Salt stress | Transgenic overexpression of these two genes (separately) in Arabidopsis plants was found to significantly enhance the ascorbic acid concentration and enhance the tolerance to salinity. | [185] |


