 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Donatella Pastore | + 3019 word(s) | 3019 | 2021-06-16 10:03:08 | | | |
| 2 | Amina Yu | + 4 word(s) | 3023 | 2021-06-23 11:07:47 | | |
Video Upload Options
Stroke is now the second leading cause of death worldwide, and is defined as an impairment in blood supply to the brain, and is linked to different cardiovascular dysfunctions. Two main types of stroke exist: ischemic stroke (due to clot-mediated blood vessel occlusion), which accounts for about 85% of all stroke cases, and hemorrhagic stroke (caused by blood vessel rupture), which accounts for 15% of total cases. Following the stroke, while the core region undergoes sudden death, the surrounding injured regions (called penumbra) may be able to recover their functions.
1. Introduction: Polyphenols
Polyphenols are micronutrients present in a variety of foods, which gained interest over the last 30 years due to their antioxidant properties and their emerging role in the prevention of several diseases linked to oxidative stress such as cancer, cardiovascular and neurodegenerative disorders [1]. Polyphenols are also secondary products of plant metabolism, whose main function is to protect organisms from damage by ultraviolet radiation and pathogens [2]. The total dietary daily intake is about 1 g/d, 10 times higher the daily antioxidant intake from Vitamin C and 100 times the one from Vitamin E [3]. a cup of tea or coffee or a glass of red wine contains about 100 mg of polyphenols [3].
Figure 1 reports the most common polyphenols and their dietary sources.
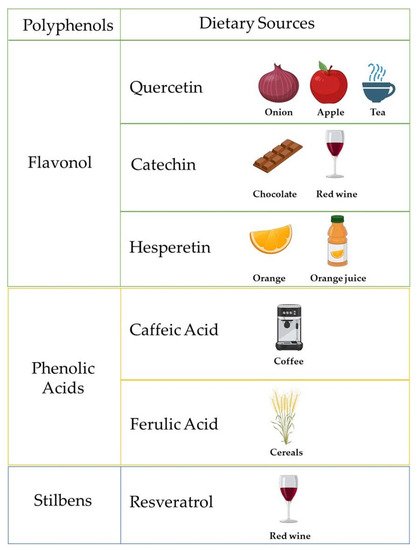
Figure 1. Schematic representation of polyphenols and their related dietary sources. Created with BioRender.com.
The most relevant scientific evidence reporting the benefic effect of polyphenols on chronic disorders refers to the so-called French paradox [4]. Although French people consume elevated levels of saturated fatty acids, which are generally associated with high mortality due to coronary heart disease, they showed low mortality [4]. The authors suggested that the high red wine consumption (and thus a high amount of resveratrol) resulted in being protective on cardiovascular risk. Accordingly, a low to moderate consumption of alcoholic drinks rich of polyphenols, such as red wine, was associated with a lower risk of cardiovascular events [5], and with a reduced mortality risk in healthy subjects, due to a decrease in coronary events, and also in patients with documented cardiovascular diseases (CVD) [6].
Several clinical and experimental studies demonstrated also that consumption of polyphenol-rich food and beverages increases plasma antioxidant capacity [7][8], and also reduces the DNA oxidative damage [9] and induces an anti-inflammatory and immune-modulating action, explaining, at least in part, their protective role on CVD [10]. In agreement with these findings, an intake of 800 g/d of fruit and vegetables was associated with a linear decrease in CVD risk [11]. Moreover, a Mediterranean dietary pattern composed of food rich in polyphenols, such as olive oil, legumes, fruits, and vegetables, is associated with a lower risk of CVD incidence and mortality [12]. A study conducted by Kokubo Y. and colleagues reported an increased consumption of soy isoflavones in post-menopausal women, which are generally at high risk of developing CVD [13] and was related to a significant reduction in the risk of myocardial and cerebral infarction [14].
Accordingly, extracts from green tea and de-alcoholated red wine inhibited platelet aggregation in in vitro studies [15][16]. Furthermore, it has been demonstrated that several polyphenols blunted endothelial dysfunction, facilitating nitric oxide-mediated vasodilation [17][18][19]. Among polyphenols, resveratrol has been reported to ameliorate endothelial function by decreasing the release of pro-inflammatory cytokines, such as interleukin (IL)-1β and tumor necrosis factor (TNF)α [20][21][22], by reducing oxidative stress in a Sirtuin1-dependent manner [23][24], and by stimulating the biosynthesis of endogenous antioxidant enzymes, such as superoxide dehydrogenase (SOD), catalase and glutathione peroxidase (GPx) [25]. Recently, we reported that Tyrosol (TR), a major polyphenol found in extra virgin olive oil (EVOO), reduced differentiation in murine 3T3-L1 preadipocytes through downregulation of adipogenic proteins, inflammation, and oxidative stress.
2. Ischemic Stroke Physiopathology
Stroke is now the second leading cause of death worldwide [26], and is defined as an impairment in blood supply to the brain [27], and is linked to different cardiovascular dysfunctions [28]. Two main types of stroke exist: ischemic stroke (due to clot-mediated blood vessel occlusion), which accounts for about 85% of all stroke cases, and hemorrhagic stroke (caused by blood vessel rupture), which accounts for 15% of total cases [29]. In particular, in the core stroke region, the blood perfusion is dramatically impaired (10–12 mL/100g/min), leading to a reduction in the oxygen and glucose supply, with a consequent decrease in energy production and neuronal death [27]. Conversely, blood perfusion in the penumbra is around 60 mL/100g/min, causing neuronal dysfunction.
Different subtypes of ischemic stroke exist. Based on the TOAST (Trial of Org 10,172 in Acute Stroke Treatment) [28] classification, in fact, five different subtypes are distinguished (Figure 2): Cardioembolic; 2. Large vessels atherosclerosis; 3.
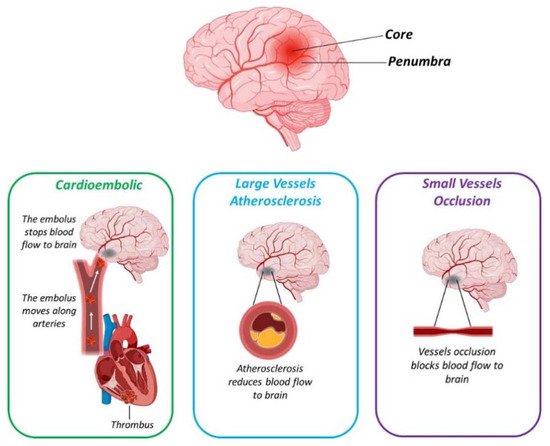
Figure 2. Schematic representation reporting the three main subtypes of ischemic stroke. Created with BioRender.com.
Cardioembolic stroke accounts for 25% to 40% of all cerebral infarction worldwide [30]. Compared to the other stroke subtypes, cardioembolic stroke is associated with the worst prognosis for patients [31]. The main risk factor is Atrial Fibrillation (AF), an alteration of the cardiac rhythm that increased with age and is also related to CVD [32].
Atherosclerosis of intracranial or extracranial arteries leads to vessel stenosis, causing a reduction in blood flow to the brain, and consequently to ischemic stroke, accounting for about 20% of all stroke cases [33][34]. The middle cerebral arteries are the main site for intracranial atherosclerosis (ICA) [33]. It has been reported that ICA is directly related to age with an increased risk ranging from 43% in 60 years old age subjects, to 80% in those older than 80 years of age [33].
Small vessel occlusion, or lacunar stroke, accounts for about 25% of stroke cases and involves the deep microcirculation of the brain [35]. Although the infarcted regions are very small compared to other subtypes of stroke, it is very harmful and patients affected by lacunar stroke showed several physical and intellectual alterations [36]. Moreover, it has been reported that about 25% of patients are prone to display a successive stroke episode within 5 years [35].
Strokes with uncommon causes account for a small group of all stroke cases [28]. Among the causes, non-atherosclerotic vasculopathies, artery dissection and prothrombotic disorder have been reported [28][29].
In some stroke cases, defined as cryptogenic or with undetermined etiology, a specific diagnosis is very difficult due to incomplete diagnostic evaluation, confounding causes, or none-specific causes assessed [28]. Thus, a brain infarcted area is recognized but none of the above-mentioned causes can be traced back to this stroke event.
Amongst the undetermined causes of stroke, we can find the embolic stroke of an undetermined source (ESUS), which accounts for an average of 17% of ischemic stroke cases, and where embolism is the leading mechanism [37]. Subjects affected by ESUS are younger compared to those with other ischemic stroke subtypes [37]. Moreover, there is an average of 4.5% annual recurrence of stroke in these patients, suggesting the relevance of a more appropriate antithrombotic prophylaxis.
Increased levels of reactive oxygen species (ROS), which contribute to enhance oxidative stress, have been reported during ischemic stroke [38]. In particular, compared to other organs, the brain resulted in being highly vulnerable to oxidative stress, since the lower presence of intracellular antioxidants levels and the presence of a large amount of intracellular lipids that can be oxidized by ROS [38]. After their release, ROS interact with several biological molecules such as protein, lipids and DNA, leading to an alteration of their structure and functions [38]. In particular, lipid peroxidation, protein denaturation, and DNA modifications are among the most common consequences of ROS accumulation [38].
Lipid peroxidation is the more harmful compared to protein oxidation, since following this initial step, a self-propagation of the oxidative process is activated, leading to an enhancement of oxidative damage [39]. An increase in proteins and lipid oxidation has been reported in both stroke patients and in mice models of stroke, such as middle cerebral artery occlusion (MCAO) [39][40][41][42]. Accordingly, a significant increase in MDA levels were found both at 24 h and 7 days post-acute ischemic stroke patients, in association with reduced levels of antioxidant enzymes [43]. Moreover, stroke-prone hypertensive rats, which developed lethal stroke, showed a dramatic increase in 4-HNE levels compared to the control group [44].
A class of protein responsible for reducing UPR was composed of the heat shock proteins (HSP) [45]. Among these, in particular, HSP70 has a pivotal role in neuronal protection following ischemic insult [45]. In an experimental model of focal ischemia, in fact, it was suggested that was an increase in HSP70 levels in the penumbra, where the neurons more resistant to ischemia are localized [46]. Accordingly, in mice, following cerebral focal ischemia, HSP70 injection of reperfusion displayed a reduction in infarct lesion and better neuronal outcomes simultaneously [47].
ROS, in fact, enhances DNA methylation levels in several animal models of ischemia and also in stroke patients [48]. Accordingly, the administration of methylation inhibitors is able to reduce brain damage in ischemic animal models Another modification observed in the animal model of IS is a decrease in several histones’ acetylation, leading to more severe neuronal damage and enhanced oxidative stress [48][49]. Moreover, HDAC1 and 2 were more active in the penumbra, according to the presence of less vulnerable neurons to stroke, while their levels were decreased in the core of lesions, where more oxidative stress and neuronal death occur [49].
Microglia activation leads to both pro- [50] and anti-inflammatory effects [51]. According to these findings, inhibition of microglia activation by using 2% isoflurane preconditioning markedly reduced inflammation and cells apoptosis in the penumbra, contributing to a decrease in MCAO-induced infarct lesion [52]. However, an impairment in microglia activation has been associated to increased infarct size and neuronal death following MCAO in mice [53]. This can be explained, at least in part, by the fact that microglia release several neurotrophic factors, such as Transforming Growth Factor Beta (TGFβ), and several anti-inflammatory cytokines, like IL-6 [54].
Astrocyte activation, after neuronal injury, enhance the release of several pro-inflammatory cytokines, such as TNFα and IL-1β, leading to an increase in oxidative stress, in neuronal death and to a reduction in neurogenesis [55][56][57]. At the same time, astrocytes produced neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), which confers neuroprotection [50]. Accordingly, the conditioned media of astrocytes following MCAO reduced the infarcted lesion, suggesting astrocytes-mediated delivery of neuroprotective factors during brain injury [58]. However, the metalloprotease released during ischemia by other cells like microglia, disrupted the connection between astrocytes and BBB, leading to its alteration and promoting the infiltration of inflammatory cells in the injured area [50][59].
Other important inflammatory cells that have harmful effects following ischemia, are leukocytes [50]. In particular, neutrophils are the first cells invading the damaged brain area, enhancing the release of pro-inflammatory cytokines, ROS production and BBB permeability, boosting the ischemic injury [60]. Moreover, leukocyte activation induced also platelet aggregation and microvasculature obstruction, leading to a reduction in blood flow to the injured brain region, dramatically potentiating the ischemic damage [50].
3. How Polyphenols Target Preventive Pathways against Ischemic Stroke
As previously reported, AF increased the risk of ischemic stroke from 4 to 5 folds [61]. In agreement, a PREDIMED (Prevención con Dieta Mediterránea) clinical trial reported the Mediterranean diet to reduce the incidence of stroke and myocardial infarction [62]. A Mediterranean diet enriched in extravergin olive oil (EVOO, which contains polyphenols) significantly reduced the risk of AF [62]. The authors hypothesized that this protection against AF may be attributable to the well established anti-inflammatory and anti-oxidant properties of polyphenols present in EVOO [62].
The authors suggested that this difference may be associated with the protective effect of the Mediterranean diet against common risk factors for IS such as diabetes mellitus and metabolic syndrome compared to those more associated with a risk of hemorrhage [63]. Moreover, it was also suggested that a Mediterranean diet is associated with a lower progression of atherosclerosis, in particular with carotid intima media thickness (IMT) [64], which is considered to be a well-established risk for diabetes mellitus and for IS rather than haemorrhagic stroke [64][65]. In a prospective study conducted on 74,961 Swedish women and men, the effects of black tea on stroke risk was evaluated [66]. The collected results highlighted that 4 or more cups of black tea daily were inversely correlated with stroke risk [66].
Taken together, all this evidence suggests that polyphenol consumption may be linked to a reduction in stroke events. Nevertheless, further studies are needed to deeply understand the molecular mechanisms underlying polyphenol-mediated protection against stroke.
Data reported are summarized in Table 1.
| Treatment | Human | Animal | Effect | References |
|---|---|---|---|---|
| Mediterranean Diet and EVOO (PREDIMED study) | X | ↓ Incidence of stroke ↓ Myocardial infarction ↓ AF |
[62] | |
| Mediterranean Diet (REGARDS study) | X | ↓ Ischemic stroke ↓ Progression of carotid intima media thickness ↓ Diabetes |
[63][64][65] | |
| Black Tea (4 or more cups) | X | ↓ Stroke risk | [66] | |
| Resveratrol | X | ↓ Infarct size ↓ Neuronal death ↑ Neurological function ↑ Shh pathway |
[67][68] | |
| Green tea extract and EGCG | X | ↓ Spatial and reference memory loss ↓ Lipid peroxidation ↓ Infarct size |
[67] | |
| EGCG | X | ↓ Infarct size ↑ Glutathione levels ↑ Neurological function ↓ Neuronal death ↑ Nrf2 levels |
[68] | |
| Pomegranate | X | ↑ Memories deficits due to MCAO | [69] | |
| Salvianolic acid B | X | ↓ Infarct size ↑ Neurological function ↓ Inflammation ↑ SIRT1 expression |
[70] |
↑ = increase and improve; ↓ = reduce.
The neuroprotective effects of polyphenols on IS have been also evaluated by several animal models. In particular, 7 days of resveratrol pretreatment before MCAO significantly reduces infarct size and improves neurological function in rats [71]. Moreover, resveratrol also decreased neuronal death by activating the neuroprotective pathway mediated by in cortical neuron culture subjected to oxygen-glucose deprivation (OGD) [71].
The results demonstrated that both these polyphenols reduced the spatial and reference memory loss induced by ischemic damage [67]. Moreover, a significant decrease in brain infarct size was observed in treated animals [67]. At the molecular level, authors found that GTx and EGCG blunted lipid peroxidation, which was enhanced following MCAO [72]. In association, increased levels of the antioxidant enzyme glutathione was reported, suggesting that polyphenols may exert a neuroprotective effect against IS through antioxidant properties.
In addition to reduced infarct size and increased glutathione levels, improved neurological scores and reduced apoptotic neuronal death have been reported for EGCG [68]. Furthermore, the authors showed a significant enhancement in nuclear factor erythroid-2 related factor 2 (Nrf2) expression following EGCG administration [68]. Since Nrf2 antioxidant activity has been previously reported to play a pivotal role against stroke injury [73], the authors suggested that EGCG neuroprotective effects may be mediated by induction of Nrf2.
Pomegranate polyphenols have been demonstrated to positively impact on several pathological conditions, such as diabetes, atherosclerosis, hyperlipidemia and cancer [74][75][76][77]. The effect of pomegranate against stroke was also evaluated [69]. In particular, pomegranate extracts were administered for two weeks before inducing MCAO in rats [69]. This study also suggested pomegranate-ameliorated behavioral deficits due to IS by crossing the BBB.
The study showed as pre-treatment with this polyphenol in rats subjected to ischemic insult significantly decreased infarct volume and improved neurological scores [70]. Moreover, an anti-inflammatory effects was also reported in association with increased expression levels of Silent information regulator 1 (SIRT1) These findings suggested that salvianolic acid B may reduce brain injury by activating the neuroprotective pathway mediated by SIRT1 as well as resveratrol.
All data reported are summarized in Table 1.
4. The Polyphenols Therapeutic Utilization against Acute Ischemic Stroke and on Stroke Rehabilitation
Following the stroke event, survivors may show cognitive impairment and functional disabilities [78]. Now, there is not an approach able to fully recover neuronal damage after stroke, impacting dramatically on individual independence, but also on public health costs. Therefore, it is mandatory to find novel theraputic targets and approaches to prevent and/or cure stroke, in order to improve neuroprotection, reducing the harmful effect induced by this disabling disease.
Particularly, pomegranate polyphenols displayed a protective role after ischemia, and chronic disorders such as hypertension, diabetes and CVD in animal models [75][79]. In a blinded-randomized clinical trial, pomegranate polyphenols or placebo pills were administered 2 weeks after a stroke event for 1 week [79]. Moreover, pomegranate administration improved cognitive and functional parameters [79]. However, few subjects were enrolled in this study and further trials involving the highest number of patients is mandatory to confirm these relevant results.
However, the treatment must be administered within 3 h from an ischemic event, otherwise fewer effects are reported in association with side effects such as intracerebral hemorrhage [80]. Twenty-four hours following administration, stroke outcomes were assessed by using the NIH stroke scale (NIHSS). Subjects belonging to the resveratrol + rtPA group showed a significant improvement in NIHSS scores in association with reduced levels of metalloproteinase (MMP)-2 and 9 (which contributes to neuronal damage following hypoxia) All these findings suggest that polyphenols may be useful to enlarge the therapeutic window for acute stroke patients leading to an improvement of stroke-related outcomes.
All data reported are summarized in Table 2.
| Treatment | Human | Animal | Effect | Reference |
|---|---|---|---|---|
| Pomegranate polyphenols | X | ↑ Cognitive and functional parameters | [79] | |
| Resveratrol, or EGCG, or Fisetin | X | ↑ Therapeutic window ↑ NIHSS scores ↓ MMP-2 and MM-9 levels |
[81][82][83][84] | |
| Salvianolic acid | X | ↑ Behavioural tests ↑ Shh pathway leading to neurogenesis ↑ Angiogenesis mediated by JAK2/STAT3 pathway |
[85] | |
| Resveratrol | X | ↓ Neurological deficits ↓ Cerebral edema ↓ Inflammation ↑ Th2 anti-inflammatory response |
[86][87] |
↑ = increase and improve; ↓ = reduce.
Here we have already reported the benefic effect of salvianolic acid pre-treatment against stroke [70]. In a subsequent study, it was evaluated whether salvianolic acid for injection (SAFI) could exert protection also following distal MCAO (dMCAO) In agreement with the benefic effect of salvianolic acid, another study, showed in mice subjected to dMCAO, post-stroke treatment with this phenolic compound induced angiogenesis in a JAK2/STAT3 All these findings suggested that salvianolic acid may be considered as a therapeutic strategy for stroke recovery.
Targeting inflammation following stroke may then be considered a strategy to confer neuroprotection. Accordingly, resveratrol administration following MCAO in rats blunted neurological deficits and cerebral edema [86]. Moreover, a significant decrease in inflammation and inflammatory mediators was also observed [86]. Interestingly, the administration of a phosphatidylinositol 3-kinase/Akt (PI3K/Akt) inhibitor abolished all these benefic effects [86], suggesting that the neuroprotection exerted by resveratrol may be, at least in part, mediated by the activation of the anti-apoptotic and antioxidant PI3K/Akt pathway.
In a recent study, it has been demonstrated that resveratrol administration following a stroke event may exert a neuroprotective effect against stroke by regulating the gut–brain axis [87]. In particular, the study reported as resveratrol induced the polarization of T lymphocyte from the Th1 pro-inflammatory to the Th2 anti-inflammatory phenotype, blunting small-intestine inflammation and reducing vascular permeability. All these regulations in the inflammatory condition ultimately led to a decrease in cytokine-mediated BBB and related brain damage. Therefore, it has been speculated that the gut–brain axis is a novel therapeutic target for ischemic stroke and shed a new light for a novel relevant action of resveratrol against stroke.
All data reported are summarized in Table 2.
References
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306.
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747.
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S.
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526.
- Rotondo, S.; Di Castelnuovo, A.; de Gaetano, G. The relationship between wine consumption and cardiovascular risk: From epidemiological evidence to biological plausibility. Ital. Heart J. 2001, 2, 1–8.
- Chiva-Blanch, G.; Arranz, S.; Lamuela-Raventos, R.M.; Estruch, R. Effects of wine, alcohol and polyphenols on cardiovascular disease risk factors: Evidences from human studies. Alcohol Alcohol. 2013, 48, 270–277.
- Nakagawa, K.; Ninomiya, M.; Okubo, T.; Aoi, N.; Juneja, L.R.; Kim, M.; Yamanaka, K.; Miyazawa, T. Tea catechin supplementation increases antioxidant capacity and prevents phospholipid hydroperoxidation in plasma of humans. J. Agric. Food Chem. 1999, 47, 3967–3973.
- Rein, D.; Lotito, S.; Holt, R.R.; Keen, C.L.; Schmitz, H.H.; Fraga, C.G. Epicatechin in human plasma: In vivo determination and effect of chocolate consumption on plasma oxidation status. J. Nutr. 2000, 130, 2109S–2114S.
- Leighton, F.; Cuevas, A.; Guasch, V.; Perez, D.D.; Strobel, P.; San Martin, A.; Urzua, U.; Diez, M.S.; Foncea, R.; Castillo, O.; et al. Plasma polyphenols and antioxidants, oxidative DNA damage and endothelial function in a diet and wine intervention study in humans. Drugs Exp. Clin. Res. 1999, 25, 133–141.
- Lotito, S.B.; Zhang, W.J.; Yang, C.S.; Crozier, A.; Frei, B. Metabolic conversion of dietary flavonoids alters their anti-inflammatory and antioxidant properties. Free Radic. Biol. Med. 2011, 51, 454–463.
- Zhan, J.; Liu, Y.J.; Cai, L.B.; Xu, F.R.; Xie, T.; He, Q.Q. Fruit and vegetable consumption and risk of cardiovascular disease: A meta-analysis of prospective cohort studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 1650–1663.
- Grosso, G.; Stepaniak, U.; Micek, A.; Stefler, D.; Bobak, M.; Pajak, A. Dietary polyphenols are inversely associated with metabolic syndrome in Polish adults of the HAPIEE study. Eur. J. Nutr. 2017, 56, 1409–1420.
- Bruckert, E.; Turpin, G. Estrogens and progestins in postmenopausal women: Influence on lipid parameters and cardiovascular risk. Horm. Res. 1995, 43, 100–103.
- Kokubo, Y.; Iso, H.; Ishihara, J.; Okada, K.; Inoue, M.; Tsugane, S.; Group, J.S. Association of dietary intake of soy, beans, and isoflavones with risk of cerebral and myocardial infarctions in Japanese populations: The Japan Public Health Center-based (JPHC) study cohort I. Circulation 2007, 116, 2553–2562.
- Russo, P.; Tedesco, I.; Russo, M.; Russo, G.L.; Venezia, A.; Cicala, C. Effects of de-alcoholated red wine and its phenolic fractions on platelet aggregation. Nutr. Metab. Cardiovasc. Dis. 2001, 11, 25–29.
- Sagesaka-Mitane, Y.; Miwa, M.; Okada, S. Platelet aggregation inhibitors in hot water extract of green tea. Chem. Pharm. Bull. 1990, 38, 790–793.
- Chen, C.K.; Pace-Asciak, C.R. Vasorelaxing activity of resveratrol and quercetin in isolated rat aorta. Gen. Pharmacol. 1996, 27, 363–366.
- Honore, E.K.; Williams, J.K.; Anthony, M.S.; Clarkson, T.B. Soy isoflavones enhance coronary vascular reactivity in atherosclerotic female macaques. Fertil. Steril. 1997, 67, 148–154.
- Andriambeloson, E.; Magnier, C.; Haan-Archipoff, G.; Lobstein, A.; Anton, R.; Beretz, A.; Stoclet, J.C.; Andriantsitohaina, R. Natural dietary polyphenolic compounds cause endothelium-dependent vasorelaxation in rat thoracic aorta. J. Nutr. 1998, 128, 2324–2333.
- Tome-Carneiro, J.; Larrosa, M.; Yanez-Gascon, M.J.; Davalos, A.; Gil-Zamorano, J.; Gonzalvez, M.; Garcia-Almagro, F.J.; Ruiz Ros, J.A.; Tomas-Barberan, F.A.; Espin, J.C.; et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013, 72, 69–82.
- Bo, S.; Ciccone, G.; Castiglione, A.; Gambino, R.; De Michieli, F.; Villois, P.; Durazzo, M.; Cavallo-Perin, P.; Cassader, M. Anti-inflammatory and antioxidant effects of resveratrol in healthy smokers a randomized, double-blind, placebo-controlled, cross-over trial. Curr. Med. Chem. 2013, 20, 1323–1331.
- Chekalina, N.I.; Kazakov, Y.M.; Mamontova, T.V.; Vesnina, L.E.; Kaidashev, I.P. Resveratrol more effectively than quercetin reduces endothelium degeneration and level of necrosis factor alpha in patients with coronary artery disease. Wiad. Lek. 2016, 69, 475–479.
- Ungvari, Z.; Labinskyy, N.; Mukhopadhyay, P.; Pinto, J.T.; Bagi, Z.; Ballabh, P.; Zhang, C.; Pacher, P.; Csiszar, A. Resveratrol attenuates mitochondrial oxidative stress in coronary arterial endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1876–H1881.
- Csiszar, A.; Labinskyy, N.; Pinto, J.T.; Ballabh, P.; Zhang, H.; Losonczy, G.; Pearson, K.; de Cabo, R.; Pacher, P.; Zhang, C.; et al. Resveratrol induces mitochondrial biogenesis in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H13–H20.
- Li, H.; Xia, N.; Forstermann, U. Cardiovascular effects and molecular targets of resveratrol. Nitric Oxide 2012, 26, 102–110.
- Gorelick, P.B. The global burden of stroke: Persistent and disabling. Lancet Neurol. 2019, 18, 417–418.
- Deb, P.; Sharma, S.; Hassan, K.M. Pathophysiologic mechanisms of acute ischemic stroke: An overview with emphasis on therapeutic significance beyond thrombolysis. Pathophysiology 2010, 17, 197–218.
- Adams, H.P., Jr.; Biller, J. Classification of subtypes of ischemic stroke: History of the trial of org 10172 in acute stroke treatment classification. Stroke 2015, 46, e114–e117.
- Parmer, P. Stroke: Classification and diagnosis. Clin. Pharm. 2018.
- Topcuoglu, M.A.; Liu, L.; Kim, D.E.; Gurol, M.E. Updates on Prevention of Cardioembolic Strokes. J. Stroke 2018, 20, 180–196.
- Kamel, H.; Healey, J.S. Cardioembolic Stroke. Circ. Res. 2017, 120, 514–526.
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.H.; McAnulty, J.H., Jr.; Zheng, Z.J.; et al. Worldwide epidemiology of atrial fibrillation: A Global Burden of Disease 2010 Study. Circulation 2014, 129, 837–847.
- Banerjee, C.; Chimowitz, M.I. Stroke Caused by Atherosclerosis of the Major Intracranial Arteries. Circ. Res. 2017, 120, 502–513.
- Marulanda-Londono, E.; Chaturvedi, S. Stroke due to large vessel atherosclerosis: Five new things. Neurol. Clin. Pract. 2016, 6, 252–258.
- Wardlaw, J.M. What causes lacunar stroke? J. Neurol. Neurosurg. Psychiatry 2005, 76, 617–619.
- Regenhardt, R.W.; Das, A.S.; Lo, E.H.; Caplan, L.R. Advances in Understanding the Pathophysiology of Lacunar Stroke: A Review. JAMA Neurol. 2018, 75, 1273–1281.
- Hart, R.G.; Catanese, L.; Perera, K.S.; Ntaios, G.; Connolly, S.J. Embolic Stroke of Undetermined Source: A Systematic Review and Clinical Update. Stroke 2017, 48, 867–872.
- Allen, C.L.; Bayraktutan, U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int. J. Stroke 2009, 4, 461–470.
- Li, S.; Zheng, J.; Carmichael, S.T. Increased oxidative protein and DNA damage but decreased stress response in the aged brain following experimental stroke. Neurobiol. Dis. 2005, 18, 432–440.
- Dominguez, C.; Delgado, P.; Vilches, A.; Martin-Gallan, P.; Ribo, M.; Santamarina, E.; Molina, C.; Corbeto, N.; Rodriguez-Sureda, V.; Rosell, A.; et al. Oxidative stress after thrombolysis-induced reperfusion in human stroke. Stroke 2010, 41, 653–660.
- Ferretti, G.; Bacchetti, T.; Masciangelo, S.; Nanetti, L.; Mazzanti, L.; Silvestrini, M.; Bartolini, M.; Provinciali, L. Lipid peroxidation in stroke patients. Clin. Chem. Lab. Med. 2008, 46, 113–117.
- Bromont, C.; Marie, C.; Bralet, J. Increased lipid peroxidation in vulnerable brain regions after transient forebrain ischemia in rats. Stroke 1989, 20, 918–924.
- Cojocaru, I.M.; Cojocaru, M.; Sapira, V.; Ionescu, A. Evaluation of oxidative stress in patients with acute ischemic stroke. Rom. J. Intern. Med. 2013, 51, 97–106.
- Guo, J.M.; Liu, A.J.; Zang, P.; Dong, W.Z.; Ying, L.; Wang, W.; Xu, P.; Song, X.R.; Cai, J.; Zhang, S.Q.; et al. ALDH2 protects against stroke by clearing 4-HNE. Cell Res. 2013, 23, 915–930.
- Kim, J.Y.; Kim, J.W.; Yenari, M.A. Heat shock protein signaling in brain ischemia and injury. Neurosci. Lett. 2020, 715, 134642.
- Sharp, F.R.; Lu, A.; Tang, Y.; Millhorn, D.E. Multiple molecular penumbras after focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2000, 20, 1011–1032.
- Doeppner, T.R.; Nagel, F.; Dietz, G.P.; Weise, J.; Tonges, L.; Schwarting, S.; Bahr, M. TAT-Hsp70-mediated neuroprotection and increased survival of neuronal precursor cells after focal cerebral ischemia in mice. J. Cereb. Blood Flow Metab. 2009, 29, 1187–1196.
- Stanzione, R.; Cotugno, M.; Bianchi, F.; Marchitti, S.; Forte, M.; Volpe, M.; Rubattu, S. Pathogenesis of Ischemic Stroke: Role of Epigenetic Mechanisms. Genes 2020, 11, 89.
- Zhao, H.; Han, Z.; Ji, X.; Luo, Y. Epigenetic Regulation of Oxidative Stress in Ischemic Stroke. Aging Dis. 2016, 7, 295–306.
- Jayaraj, R.L.; Azimullah, S.; Beiram, R.; Jalal, F.Y.; Rosenberg, G.A. Neuroinflammation: Friend and foe for ischemic stroke. J. Neuroinflamm. 2019, 16, 142.
- Parada, E.; Egea, J.; Buendia, I.; Negredo, P.; Cunha, A.C.; Cardoso, S.; Soares, M.P.; Lopez, M.G. The microglial alpha7-acetylcholine nicotinic receptor is a key element in promoting neuroprotection by inducing heme oxygenase-1 via nuclear factor erythroid-2-related factor 2. Antioxid. Redox Signal. 2013, 19, 1135–1148.
- Sun, M.; Deng, B.; Zhao, X.; Gao, C.; Yang, L.; Zhao, H.; Yu, D.; Zhang, F.; Xu, L.; Chen, L.; et al. Isoflurane preconditioning provides neuroprotection against stroke by regulating the expression of the TLR4 signalling pathway to alleviate microglial activation. Sci. Rep. 2015, 5, 11445.
- Lalancette-Hebert, M.; Swarup, V.; Beaulieu, J.M.; Bohacek, I.; Abdelhamid, E.; Weng, Y.C.; Sato, S.; Kriz, J. Galectin-3 is required for resident microglia activation and proliferation in response to ischemic injury. J. Neurosci. 2012, 32, 10383–10395.
- Nakajima, K.; Kohsaka, S. Microglia: Neuroprotective and neurotrophic cells in the central nervous system. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 65–84.
- Tuttolomondo, A.; Di Raimondo, D.; di Sciacca, R.; Pinto, A.; Licata, G. Inflammatory cytokines in acute ischemic stroke. Curr. Pharm. Des. 2008, 14, 3574–3589.
- Monje, M.L.; Toda, H.; Palmer, T.D. Inflammatory blockade restores adult hippocampal neurogenesis. Science 2003, 302, 1760–1765.
- Stoll, G.; Jander, S.; Schroeter, M. Inflammation and glial responses in ischemic brain lesions. Prog. Neurobiol. 1998, 56, 149–171.
- Kinoshita, A.; Yamada, K.; Kohmura, E.; Hayakawa, T. Effect of astrocyte-derived factors on ischemic brain edema induced by rat MCA occlusion. APMIS 1990, 98, 851–857.
- del Zoppo, G.J. The neurovascular unit in the setting of stroke. J. Intern. Med. 2010, 267, 156–171.
- Martynov, M.Y.; Gusev, E.I. Current knowledge on the neuroprotective and neuroregenerative properties of citicoline in acute ischemic stroke. J. Exp. Pharmacol. 2015, 7, 17–28.
- Saposnik, G.; Gladstone, D.; Raptis, R.; Zhou, L.; Hart, R.G. Atrial fibrillation in ischemic stroke: Predicting response to thrombolysis and clinical outcomes. Stroke 2013, 44, 99–104.
- Martinez-Gonzalez, M.A.; Toledo, E.; Aros, F.; Fiol, M.; Corella, D.; Salas-Salvado, J.; Ros, E.; Covas, M.I.; Fernandez-Crehuet, J.; Lapetra, J.; et al. Extravirgin olive oil consumption reduces risk of atrial fibrillation: The PREDIMED (Prevencion con Dieta Mediterranea) trial. Circulation 2014, 130, 18–26.
- Tsivgoulis, G.; Psaltopoulou, T.; Wadley, V.G.; Alexandrov, A.V.; Howard, G.; Unverzagt, F.W.; Moy, C.; Howard, V.J.; Kissela, B.; Judd, S.E. Adherence to a Mediterranean diet and prediction of incident stroke. Stroke 2015, 46, 780–785.
- Esposito, K.; Giugliano, D. Mediterranean diet for primary prevention of cardiovascular disease. N. Engl. J. Med. 2013, 369, 674–675.
- Tsivgoulis, G.; Vemmos, K.N.; Spengos, K.; Papamichael, C.M.; Cimboneriu, A.; Zis, V.; Zakopoulos, N.; Mavrikakis, M. Common carotid artery intima-media thickness for the risk assessment of lacunar infarction versus intracerebral haemorrhage. J. Neurol. 2005, 252, 1093–1100.
- Larsson, S.C.; Virtamo, J.; Wolk, A. Black tea consumption and risk of stroke in women and men. Ann. Epidemiol. 2013, 23, 157–160.
- Wu, K.J.; Hsieh, M.T.; Wu, C.R.; Wood, W.G.; Chen, Y.F. Green Tea Extract Ameliorates Learning and Memory Deficits in Ischemic Rats via Its Active Component Polyphenol Epigallocatechin-3-gallate by Modulation of Oxidative Stress and Neuroinflammation. Evid. Based Complement. Altern. Med. 2012, 2012, 163106.
- Han, J.; Wang, M.; Jing, X.; Shi, H.; Ren, M.; Lou, H. (-)-Epigallocatechin gallate protects against cerebral ischemia-induced oxidative stress via Nrf2/ARE signaling. Neurochem. Res. 2014, 39, 1292–1299.
- Sarkaki, A.; Rezaiei, M.; Gharib Naseri, M.; Rafieirad, M. Improving active and passive avoidance memories deficits due to permanent cerebral ischemia by pomegranate seed extract in female rats. Malays. J. Med. Sci. 2013, 20, 25–34.
- Lv, H.; Wang, L.; Shen, J.; Hao, S.; Ming, A.; Wang, X.; Su, F.; Zhang, Z. Salvianolic acid B attenuates apoptosis and inflammation via SIRT1 activation in experimental stroke rats. Brain Res. Bull. 2015, 115, 30–36.
- Yu, P.; Wang, L.; Tang, F.; Zeng, L.; Zhou, L.; Song, X.; Jia, W.; Chen, J.; Yang, Q. Resveratrol Pretreatment Decreases Ischemic Injury and Improves Neurological Function Via Sonic Hedgehog Signaling After Stroke in Rats. Mol. Neurobiol. 2017, 54, 212–226.
- Della-Morte, D.; Dave, K.R.; DeFazio, R.A.; Bao, Y.C.; Raval, A.P.; Perez-Pinzon, M.A. Resveratrol pretreatment protects rat brain from cerebral ischemic damage via a sirtuin 1-uncoupling protein 2 pathway. Neuroscience 2009, 159, 993–1002.
- Alfieri, A.; Srivastava, S.; Siow, R.C.; Modo, M.; Fraser, P.A.; Mann, G.E. Targeting the Nrf2-Keap1 antioxidant defence pathway for neurovascular protection in stroke. J. Physiol. 2011, 589, 4125–4136.
- Esmaillzadeh, A.; Tahbaz, F.; Gaieni, I.; Alavi-Majd, H.; Azadbakht, L. Concentrated pomegranate juice improves lipid profiles in diabetic patients with hyperlipidemia. J. Med. Food 2004, 7, 305–308.
- Aviram, M.; Dornfeld, L.; Rosenblat, M.; Volkova, N.; Kaplan, M.; Coleman, R.; Hayek, T.; Presser, D.; Fuhrman, B. Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: Studies in humans and in atherosclerotic apolipoprotein E-deficient mice. Am. J. Clin. Nutr. 2000, 71, 1062–1076.
- Tibullo, D.; Caporarello, N.; Giallongo, C.; Anfuso, C.D.; Genovese, C.; Arlotta, C.; Puglisi, F.; Parrinello, N.L.; Bramanti, V.; Romano, A.; et al. Antiproliferative and Antiangiogenic Effects of Punica granatum Juice (PGJ) in Multiple Myeloma (MM). Nutrients 2016, 8, 611.
- Sharma, P.; McClees, S.F.; Afaq, F. Pomegranate for Prevention and Treatment of Cancer: An Update. Molecules 2017, 22, 177.
- Cumming, T.B.; Marshall, R.S.; Lazar, R.M. Stroke, cognitive deficits, and rehabilitation: Still an incomplete picture. Int. J. Stroke 2013, 8, 38–45.
- Bellone, J.A.; Murray, J.R.; Jorge, P.; Fogel, T.G.; Kim, M.; Wallace, D.R.; Hartman, R.E. Pomegranate supplementation improves cognitive and functional recovery following ischemic stroke: A randomized trial. Nutr. Neurosci. 2019, 22, 738–743.
- Cheng, N.T.; Kim, A.S. Intravenous Thrombolysis for Acute Ischemic Stroke Within 3 Hours Versus Between 3 and 4.5 Hours of Symptom Onset. Neurohospitalist 2015, 5, 101–109.
- Chen, J.; Bai, Q.; Zhao, Z.; Sui, H.; Xie, X. Resveratrol improves delayed r-tPA treatment outcome by reducing MMPs. Acta Neurol. Scand. 2016, 134, 54–60.
- Yang, Y.; Rosenberg, G.A. Matrix metalloproteinases as therapeutic targets for stroke. Brain Res. 2015, 1623, 30–38.
- Wang, X.H.; You, Y.P. Epigallocatechin Gallate Extends Therapeutic Window of Recombinant Tissue Plasminogen Activator Treatment for Brain Ischemic Stroke: A Randomized Double-Blind and Placebo-Controlled Trial. Clin. Neuropharmacol. 2017, 40, 24–28.
- Wang, L.; Cao, D.; Wu, H.; Jia, H.; Yang, C.; Zhang, L. Fisetin Prolongs Therapy Window of Brain Ischemic Stroke Using Tissue Plasminogen Activator: A Double-Blind Randomized Placebo-Controlled Clinical Trial. Clin. Appl. Thromb. Hemost. 2019, 25.
- Zhang, Y.; Zhang, X.; Cui, L.; Chen, R.; Zhang, C.; Li, Y.; He, T.; Zhu, X.; Shen, Z.; Dong, L.; et al. Salvianolic Acids for Injection (SAFI) promotes functional recovery and neurogenesis via sonic hedgehog pathway after stroke in mice. Neurochem. Int. 2017, 110, 38–48.
- Lei, J.; Chen, Q. Resveratrol attenuates brain damage in permanent focal cerebral ischemia via activation of PI3K/Akt signaling pathway in rats. Neurol. Res. 2018, 40, 1014–1020.
- Dou, Z.; Rong, X.; Zhao, E.; Zhang, L.; Lv, Y. Neuroprotection of Resveratrol Against Focal Cerebral Ischemia/Reperfusion Injury in Mice Through a Mechanism Targeting Gut-Brain Axis. Cell. Mol. Neurobiol. 2019, 39, 883–898.

