Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hugo Fernandes-Silva | + 5103 word(s) | 5103 | 2021-06-03 10:43:44 | | | |
| 2 | Vivi Li | -36 word(s) | 5067 | 2021-06-23 04:43:53 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Fernandes-Silva, H. Retinoic Acid and Lung Development. Encyclopedia. Available online: https://encyclopedia.pub/entry/11160 (accessed on 27 July 2026).
Fernandes-Silva H. Retinoic Acid and Lung Development. Encyclopedia. Available at: https://encyclopedia.pub/entry/11160. Accessed July 27, 2026.
Fernandes-Silva, Hugo. "Retinoic Acid and Lung Development" Encyclopedia, https://encyclopedia.pub/entry/11160 (accessed July 27, 2026).
Fernandes-Silva, H. (2021, June 22). Retinoic Acid and Lung Development. In Encyclopedia. https://encyclopedia.pub/entry/11160
Fernandes-Silva, Hugo. "Retinoic Acid and Lung Development." Encyclopedia. Web. 22 June, 2021.
Copy Citation
Retinoic acid (RA) is a key molecular player in embryogenesis and adult tissue homeostasis. In embryo development, RA plays a crucial role in the formation of different organ systems, namely, the respiratory system. During lung development, there is a spatiotemporal regulation of RA levels that assures the formation of a fully functional organ. RA signaling influences lung specification, branching morphogenesis, and alveolarization by regulating the expression of particular target genes. Moreover, cooperation with other developmental pathways is essential to shape lung organogenesis.
vitamin A
retinol
respiratory system
lung specification
branching morphogenesis
alveologenesis
1. General Introduction
Retinoic acid (RA), the active metabolite of retinol (Vitamin A), plays countless roles in different organs and tissues. In the adult, RA is crucial for immune, nervous and reproductive systems, as well as for vision and skin renewal. Throughout embryogenesis, RA acts as a potent morphogen that mediates cellular signaling and transcriptional regulation, thus modulating numerous aspects of embryo development [1][2][3]. RA levels must be tightly regulated, if not, RA can act as a teratogen, rather than a morphogen. In fact, deficiency or excess of RA causes severe congenital malformations that may affect somite, skeletal, spinal cord, limb, heart, pancreas, eye, diaphragm, and lung development [2].
Regarding lung development, RA regulates cellular events such as proliferation, differentiation, patterning, and maturation [4]. Evidently, this signaling pathway “does not walk alone”, and the interplay with other well-known regulators is critical for proper lung formation.
2. Lung Development and Retinoic Acid
Lung development is a highly coordinated and multistage process governed by mechanical, anatomical, and biochemical events. In humans, it begins around the fourth week after conception and continues into the post-natal period approximately until 22 years of age [5][6][7]. The mature lung displays a tree-like tubular system connected to the trachea and a highly branched vascular system. The conducting (proximal) airways differ from the respiratory (distal) airways. In fact, towards the distal portion of the lung, the airways become progressively smaller/narrower, ending in the alveolar region where gas exchange occurs [8][9].
In humans, lung organogenesis initiates with the appearance of an endodermal bud on the ventral side of the anterior foregut around the fourth week of gestation. The endodermal evagination leads to the formation of the two lung buds and trachea, surrounded by mesoderm and a vascular network [9]. Lung epithelium is endoderm-derived and lines the airways and the alveoli. The conducting airways are generated by a highly regulated dichotomous branching morphogenesis commencing on the 5th week of gestation, giving rise to a complex network with numerous terminal branches. The continuous bifurcation of the epithelial bud tips relies on the close interaction between epithelium and mesenchyme. The mesenchyme at the tips is particularly important since it contains precursor cells that will differentiate into smooth muscle that stabilize the cleft between the newly formed buds. At the same time, diffusible signals act in both compartments, orchestrating lung organogenesis processes [10][11][12]. The alveolar cellular differentiation begins between the 16th and the 24th week of gestation. During this phase, bronchial epithelial cells differentiate into alveolar epithelial cells type 1 (AEC1) and type 2 (AEC2), also known as type 1 and type 2 pneumocytes, respectively. AEC2 act as AEC1 progenitor cells and are responsible for surfactant production, whereas AEC1 are responsible for gas exchange. From the 24th week onwards, tissue projection into the distal airspaces gives rise to structures resembling sacs, the primitive alveoli, lined with AEC1 and AEC2. Although the alveoli are formed at birth, their maturation begins approximately five weeks after birth, with the formation of a fine air-blood barrier composed by a thin epithelial sheet, an endothelial layer, and a basement membrane. Even though in the first years of life, the alveolar size remains constant, in adolescence, with the enlargement of the thoracic cage, it increases [13][14][15].
The crosstalk between the epithelial and mesenchymal compartment is conveyed by several diffusible signals from multiple signaling pathways including Sonic Hedgehog (SHH) [16], Wingless-related Integration Site (WNT) [17], Transforming Growth Factor β (TGFβ), Bone Morphogenetic Protein (BMP) [18], Fibroblast Growth Factor (FGF) [19], Hippo [20], and Retinoic Acid [4], just to name a few. In the next sections, we will dissect the role of RA throughout the five lung developmental phases, namely, embryonic, pseudoglandular, canalicular, saccular, and alveolar (Figure 1), and also in lung vascular development.
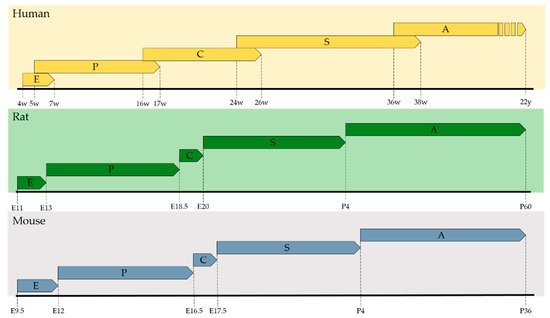
Figure 1. Lung developmental phases. Time scale of human, rat and mouse lung developmental phases (E = Embryonic; P = Pseudoglandular; C = Canalicular; S = Saccular; A = Alveolar). The phases are defined largely by morphological standards and may overlap in time. Adapted from [14].
2.1. Embryonic Phase
The respiratory system is specified from the anterior foregut endoderm approximately at embryonic day 9.5 (E9.5) in mouse and around the 4th week of gestation in humans [14]. Initially, the anterior foregut begins as a sole epithelial tube surrounded by mesoderm. Lung and trachea specification begins with the establishment of a localized expression domain of the transcription factor NKX2.1 (also known as TTF1) in the ventral wall of the anterior foregut [21]. The expression of nkx2.1 on the ventral side, distally to the rudimentary trachea, leads to the evagination of the two primary lung buds [9]. The epithelial outgrowth of the primary buds is supported by diffusible signals from the mesoderm tightly regulated both in time, dosage, and space. Briefly, WNT2 and WNT2b expression specify NKX2.1+ respiratory endoderm progenitors in the ventral anterior mesoderm that surrounds the anterior foregut endoderm [22]. The expression of NKX2.1 also depends on BMP signaling activation. Endodermal SHH regulates BMP4 expression in the ventral mesenchyme surrounding the ventral foregut, mainly through the transcription factors FOXF1, GLI1, and GLI3. BMP signaling represses the expression of the transcription factor SOX2 (that promotes esophageal fate), thus enabling NKX2.1 endodermal expression [23]. After respiratory lineage specification, FGF10 mesodermal expression requires TGFβ inhibition, mediated by RA signaling, to conduct outgrowth and extension of the primary lung buds [24].
RA signaling machinery is present in the foregut at the beginning of lung organogenesis. At E9.5, raldh2 is highly and ubiquitously expressed in the anterior foregut, specifically in the mesenchyme neighboring the prospective trachea and lung primordia, suggesting that RA is being synthesized. Moreover, at this gestational age, a high local RAR activation is also detected in all layers of the foregut where trachea and primordial lung are emerging. This expression pattern is consistent with the hypothesis that RA acts on its receptors present in the epithelium in early lung development. At E10, raldh2 levels are preserved in the tracheal mesenchyme and proximal lung but decrease towards the distal lung. Conversely, cyp26 is not detected between E9.5–10; consequently, at these stages, RA synthesis is not compensated by its degradation, pointing to a crucial role for RA in the formation of the lung bud primordium. It is worth noting that, as lung morphogenesis proceeds, cyp26 is progressively expressed, therefore controlling RA levels [25].
Disruption of RA signaling in in vitro mouse foregut cultures impairs lung bud initiation [26]. Likewise, raldh2−/− knockout mice and dams exposed to severe vitamin A deficiency during gestation display lung agenesis [27][28]. In vitro studies revealed that RA induces fgf10 expression in the foregut mesoderm, where lung initiates, which then activates FGFR2 signaling in the endoderm and induces primary bud morphogenesis [26]. By further dissecting raldh2−/− knockout mice, it was uncovered that RARβ mediates fgf10 induction by RA. Differential activation of RARα and RARβ has opposite effects in fgf10 mesodermal expression; nonetheless, both receptors are needed for proper lung development [28]. Furthermore, it has been shown that endogenous RA acts via WNT and TGFβ signaling to control fgf10 expression. RA downregulates TGFβ activity in the foregut mesoderm, thus allowing local expression of fgf10, which is crucial for primary lung bud induction [24]. Conversely, RA facilitates the activation of WNT signaling, crucial for the appearance/maintenance of the respiratory field, by inhibiting the expression of a WNT/β-catenin antagonist (Dickkopf1) [29]. Hence, an RA-dependent fine-tuned equilibrium of the WNT/TGFβ/FGF10 axis is mandatory to permit both respiratory progenitors’ expansion and lung primordium formation.
RA acts in both endoderm and mesoderm to provide a niche for lung induction by regulating the expression of signals that will induce the lung and the competence to respond. In early somite stages, RA pre-patterns the lateral plate mesoderm and establishes the anterior foregut region that will form the respiratory niche. Subsequently, RA signaling promotes endodermal Hedgehog (HH) expression, which, mediated by mesenchymal GLI transcription factors, is indispensable for wnt2/2b and bmp4 expression; subsequently, wnt2/2b and bmp4 act on the endoderm to promote nkx2.1 expression. The RA-HH-WNT signaling cascade that coordinates respiratory lineage specification is conserved between frog, mouse, and human [30]. Moreover, RA/RAR activity regulates the competence of the endoderm to activate the NKX2.1+ respiratory program in response to mesodermal WNT and BMP, independently of HH, during early somite stages of development.
Recently, it has been shown that RA signaling has different roles during endoderm organogenesis, acting in a distinct temporal and spatial pattern. In early gastrula stages (mouse E6–7.5), during endoderm formation, RA induces hindgut and pancreatic stages but inhibits foregut fate. Conversely, in early somitogenesis (mouse E7.5–8.5), during endoderm patterning, RA suppresses pharyngeal and promotes respiratory fate in the anterior endoderm. Finally, lung induction (mouse E9.5) relies on the RA/WNT/BMP axis previously described [30]. However, RA actions are only possible because, during the gastrula stage, WNT/BMP signaling specifies endodermal competence domains and, consequently, how cells respond to signals during the subsequent phases [31].
2.2. Pseudoglandular Phase
Lung branching morphogenesis occurs during the pseudoglandular phase (E12–16.5 mouse; E13–E18.5 rat; 5–17 weeks, human), and it is characterized by reiterative splitting of the airway epithelium into the surrounding mesenchyme [32]. The development of new generations of terminal buds contributes to defining the airway conducting system. This process is tightly regulated by a network of signaling cascades that operate via epithelial-mesenchymal interactions [14]. The RA pathway has been recognized as a critical regulator of pulmonary branching [4][25][33][34]. In fact, in rdh10−/− mutants, primary lung bud growth is arrested and branching morphogenesis impaired, thus preventing the formation of the lung [35]. Likewise, raldh2−/− mouse embryos display defective growth and branching, resulting in hypoplastic lungs [27].
RA signaling machinery is dynamically expressed during branching morphogenesis. For instance, stra6 mouse transcript is present in the mesenchyme surrounding the bronchi [36]. Conversely, as lung branching initiates, crbp transcripts are present in both epithelial and mesenchymal compartments, and later its expression is restricted to the mesenchyme [37]. In the embryonic mouse lung, raldh2 is expressed along the mesothelium surface at sites of low branching activity. By comparing fgf10 and raldh2 expression patterns, it is possible to observe that regions, where fgf10 is expressed, are low in raldh2 [25][38]. Conversely, raldh1 mRNA is present in the proximal region of the bronchi [25][33], while raldh3 is weakly expressed in the epithelium of the main bronchus [39]. Importantly, RA bioavailability must be balanced to avoid detrimental concentrations. As secondary buds start to emerge, cyp26 expression displays a proximal-distal epithelial gradient with high expression at distal sites. In the mesenchymal compartment, cyp26 is present between the secondary buds contributing to the regulation of RA levels in this region [25].
Retinoic acid signaling response is, ultimately, transduced through RAR and RXR nuclear receptors that recognize RARE sequences in the promoter region of RA-target genes. The importance of RAR is particularly evident in rarαβ2 mouse mutants, which exhibit lung agenesis and hypoplasia due to the absence of left lung budding and altered branching morphogenesis [40]. In the developing lung, rarα1 and rarα2 are ubiquitously expressed [33][41], whereas rarβ expression is restricted to the proximal bronchi epithelium and the proximal subepithelial and subpleural mesenchyme [25][33][37]. In opposition, rarγ transcripts are faintly and ubiquitously expressed in the embryonic lung; indeed, RARγ knockouts do not affect lung branching [25][33][37][41][42][43]. Throughout branching, RARE-lacZ reporter activity is detected in the proximal bronchial region with a pattern that strongly overlaps rarβ expression. Moreover, RARE-lacZ lacks from both the distal bud epithelium and the mesenchymal compartment [25]. Furthermore, RXR can be sequestered by Chicken Ovalbumin upstream promoter-transcription factors (COUP-TFs), resulting in RA signaling inhibition. coup-tfII expression is found in the lung mesenchyme, particularly higher in the mesenchyme between the secondary buds, but absent from the pulmonary epithelium [25]. Recently, RA signaling was characterized in the embryonic chicken lung [34]. For instance, stra6 is expressed in the periepithelial mesenchyme surrounding the epithelium as it occurs in the mouse lung [34][36]. raldh2, raldh3, cyp26a1, and rarα expression patterns are comparable to their mammalian counterparts. Curiously, rarβ has a stage-dependent expression that is present in the proximal region of the mesenchymal compartment [34]. It is worth mentioning that while in the mammalian lung RA signaling members are distributed between the mesenchymal and epithelial compartments, in the chick lung, all members seem to be restricted to the mesenchyme [34]. Nonetheless, transcripts like stra6 and raldh2 display highly conserved expression patterns. Moreover, the complementary expression pattern between raldh2 and fgf10 is conserved, with raldh2 being expressed in the ventral region, and fgf10 in the dorsal pulmonary region [34][44].
RA signaling must be finely tuned so that mammalian lung branching morphogenesis may occur. In the mesenchymal compartment, RA is produced in the mesothelial region, at low branching sites, by RALDH2. Conversely, RA synthesis is regulated by CYP26 and COUP-TFs in the mesenchyme between the secondary buds [25]. A gradient of RA is produced from the pleura to the periepithelial mesenchyme that surrounds the distal region of the growing bud. RA availability regulates fgf10 that is typically expressed in the mesenchyme surrounding the distal bud tips [25][38]; for this reason, raldh2−/− mouse embryos phenotype is rescued by RA and FGF10 supplementation [27]. Concurrently, in the epithelial compartment, RA is produced in the proximal bronchi by RALDH1 [25][33] and may act through RARβ, which is detected in the same region [25][33][37]. RA epithelial levels are regulated by CYP26, which displays a gradient from the distal to the proximal region of the epithelial bronchi [25]. Intriguingly, stra6 expression in the periepithelial mesenchyme, surrounding the epithelial bronchi, may play a role in retinol trafficking between epithelium and mesenchyme [36].
The role of RA signaling during pulmonary branching was uncovered by performing RA stimulation/inhibition studies. Mouse lung explants exposed to RA display an increase in lung branching in a dose-dependent manner [45]. Furthermore, RA supplementation can increase pulmonary branching in nitrofen-induced hypoplastic lungs, which display decreased RALDH2 activity [46]. Similarly, rat lung explants treated with exogenous RA exhibit an increase in the number of peripheral airway buds, epithelial perimeter, and the total area of the lung [47]. Likewise, RA supplementation, in the chicken model, revealed a dose-dependent increase in lung branching [34]. Furthermore, RA-exposed lung explants can display morphological alterations that resemble a more immature branching configuration; specifically, the formation of typical distal bud-like structures is impaired, promoting a more proximal-like phenotype [25][48][49]. Nevertheless, in some studies, a reduced number of terminal buds is observed in RA-treated lungs [25][41][48][49].
RA pathway modulation studies revealed a series of feedback control mechanisms underlying pulmonary branching morphogenesis. For instance, crbpI and raldh1 expression are susceptible to be modulated by activation/inhibition of the RA cascade [33][41]. This feedback loop mechanism may regulate RA synthesis in the proximal epithelial buds [33]. On the other hand, RA supplementation increases both rarβ and RARβ but decreases RARα and RARγ [41][47].
RA downstream targets are not only signaling machinery components, but also members of other pathways. For instance, RA can directly downregulate fgf10 expression or indirectly by stimulating the SHH pathway (through increasing shh expression) and, consequently, decrease fgf10 transcript levels in the distal region of the lung [25][33][38][49][50]. Moreover, bmp4 is also negatively modulated by RA signaling stimulation [25]. The interaction between RA, FGF, SHH, and BMP4 is crucial for branching and proximal-distal patterning [25][33][38][49][50][51][52]. In the proximal epithelium, RA modulates the expression of foxa2, a known regulator of epithelial differentiation and pulmonary branching [33][53]. RA pathway stimulation promotes tgfβ3 expression in the lobar bronchial mesenchyme, pleura region, and bronchial epithelium [33], and downregulates cftr epithelial expression in the lobar bronchial and bronchial epithelium [33]. In fact, mammalian lung branching morphogenesis can be divided into two RA responsive centers: the distal epithelial region where shh, and bmp4 are expressed and interact with fgf10 mesenchymal expression; and the proximal epithelial region where RA regulates tgfβ3, foxa2 and cftr [25][33][38][50].
There are other RA downstream targets described in the embryonic mammalian lung, namely hoxa2, hoxa5, hoxb5, and hoxb6, which are related to patterning. Moreover, RA regulates HOXB5 protein levels, and hoxa2 and hoxb6 proximal-distal patterns [49][54][55]. In addition, disruption of the RA pathway results in altered murine smooth muscle phenotype and a consequent increase in the expression of acta2 and myh11 [56]. During chick lung branching, RA supplementation does not affect sox2 but decreases sox9 expression (patterning-related genes); however, it affects sox9 spatial distribution, which is progressively excluded from the distal regions of the lung. Still, the new distal branches do not exhibit proximal appearance [34][57][58]. Furthermore, RA signaling stimulation, of the embryonic chick lung, increases the expression of rarβ, hoxb5, and meis2; alters the spatial distribution of cyp26a1, id2, tgfβ2, and sox9; and has a very modest effect on fgf10, fgfr2, and sox2. Even though RA machinery is confined to the mesenchymal compartment, it can influence the expression of epithelial genes such as shh, sox9, and fgfr2 [34]. In the chicken model, RA signaling stimulates early pulmonary branching and contributes to pulmonary branching proximal-distal patterning [34].
2.3. Canalicular Phase
The canalicular phase occurs between post-conception weeks 16 and 26 in human, E18.5–E20 in rat, and around E16.5–E17.5 in mouse [14]. During this phase, the epithelial airways continue to increase in size, and the terminal buds extend into the airspaces, forming the primitive pulmonary acini (terminal sacs) [59]. Simultaneously, distal mesenchyme, although still abundant at this phase, becomes progressively thinner. Additionally, early differentiation of AEC1 and AEC2 occurs, establishing an elementary gas exchange surface [9][60]; this respiratory region is morphologically different from the more proximal conducting airways [61][62][63]. Moreover, the bronchioalveolar duct junctions appear when epithelial differentiation happens and correspond to a stem cell niche; the location of this junction stays constant throughout life [64][65][66]. Finally, in this phase, vascularization begins. The continuous angiogenic process and juxtaposition of capillaries in close contact with the distal respiratory epithelium create the first air-blood barrier in the future alveolar ducts and saccules [67][68].
The role of RA in the canalicular phase has not been particularly dissected so far. However, in this phase, with the occurrence of cytodifferentiation of the columnar epithelium, the level of rarα transcripts decreases [69]. RA is a key regulator of cellular growth, proliferation, differentiation, and embryogenesis per se, and one cannot discard that it may contribute to this phase of lung morphogenesis. For instance, retinoic acid is a major regulator of angiogenesis and vasculogenesis [70], and the organization of the pulmonary vascular bed initiates during the canalicular period. Therefore, RA may be involved in the development of the lung vascular system.
2.4. Saccular Phase
The pulmonary saccular phase is characterized by the widening and growing of the terminal airways, which contribute to forming clusters of larger airspaces called saccules. During this phase (weeks 24–38 in human; E20–P4 in rat; E17.5–P4 in mouse), the enlargement of the future gas-exchange region causes condensation of the mesenchyme in-between airspaces, establishing the primary septa. Primary septa are composed of a double layer of capillaries separated by a central layer of mesenchyme. The majority of the primary septa surface is covered by AEC1, and the remaining surface is occupied by AEC2. Throughout this phase, lamellar bodies appear, and surfactant secretion initiates. Smooth muscle cell precursors start to form a network of elastic fiber and collagen fibrils that prepare the lung for alveolarization [14][71][72].
rar transcripts are present in the embryonic rat and mouse lung during the canalicular phase [69][73]. The expression levels of rarα transcript decrease in this phase [69], which seems to be required to allow sacculation and the differentiation program that contributes to forming mature AEC1 [73]. On the other hand, rarβ expression increases significantly in the late saccular period. This increase in rarβ matches AEC1 and AEC2 induction, suggesting a role for rarβ in preparing the lung for the alveolar phase. Low expression levels of rarγ were detected in this period [69]. Such findings point to a crucial role of the RA signaling pathway in preparing the lung for sacculation, and later, during the saccular phase, to prepare the lung for alveolarization.
2.5. Alveolar Phase
Alveologenesis or alveolarization is the process by which the gas exchange functional units, the alveoli, are generated. Alveoli formation is mainly a postnatal event, and it is time and species-dependent. For instance, mammals like deer, guinea pigs, or sheep are born with functional alveoli; mother-dependent mammals like mice, rats, and humans are born with primitive alveoli and, so, alveologenesis continues postnatally [74]. In mouse and rat, alveologenesis occurs entirely in the postnatal period, approximately from P4 to P36 and P4 to P60, respectively [7][9][14]. In humans, part of the alveoli is formed before birth, but their genesis continues postnatally. Alveologenesis is divided into three phases: classical alveolarization (week 36 until ∼3 years), microvascular maturation (week 36 until young adulthood), and continued alveolarization (∼2 years until young adulthood) [14].
Throughout the canalicular and saccular phases, the terminal tubes are transformed into small saccules separated by the primary septum. During classical alveologenesis, the sacs are subdivided by the ingrowth of secondary septum, the outline of the future alveoli. Myofibroblast progenitors, endothelial cells, fibroblasts, and lipofibroblasts (LIFs) cover the secondary septa, and matrix proteins such elastin are deposited at the tip [7][9][75][76]. At first, alveolarization is quite fast (known as “bulk alveolarization”), but then, this process is slowed down, giving rise to the continued alveolarization. As the alveoli mature, the surrounding capillaries are remodeled, forming the capillary unit, and the endothelial cells are located in the proximity of the AEC1 cells permitting an efficient gas exchange. This process is known as microvascular maturation and occurs simultaneously with alveolarization [7][9][14][75][76][77][78].
Secondary septation is intimately associated with several signaling events. For example, platelet-derived growth factor α (PDGFα) regulates myofibroblast differentiation and elastin production; TGFβ regulates elastin expression [75][76][79], whereas ephrin B2 plays an essential role in endothelial cells. Furthermore, fgfr3/fgfr4 double null mice fail to generate the secondary septa pointing to a role for FGF signaling in controlling this event [80]. Finally, RA is crucial for proper alveolar formation [7][9].
Alveologenesis in the developing lung is partially regulated by vitamin A [81], and endogenous RA is present in the postnatal lung [82][83]. Retinoids have been recognized over the years as alveolar morphogens and possible therapeutic mediators [84]. Different studies have demonstrated that vitamin A deficiency induces major histological changes in the lung, namely: airspace enlargement, thinner alveolar walls, more alveolar breaks, and an increase in the mean free distance in the air spaces [85]. Furthermore, vitamin A deficiency impairs lung epithelial function in rodents and promotes bronchopulmonary dysplasia (BPD) in humans [86][87]. The precise mechanism by which RA regulates alveolar formation remains poorly understood, although RA regulation of myofibroblast proliferation was shown to rely on intact FGF signaling [9]. Conversely, RA supplementation in explant culture induces septation, and retinol treatment of premature lambs promotes alveologenesis [88].
RA signaling machinery is differentially expressed during alveologenesis. For instance, in mice, CRBPI is present in the alveolar walls and exhibits higher expression levels around P9, whereas CRBPII is detected during alveolar formation at P4 [82]. Furthermore, in the rat model, CRABP levels rise shortly after birth, reach its maximum values at P10, and at P21 it is barely detected. These genes were upregulated during alveologenesis in whole rat lung, and isolated LIFs [89], and the alterations in the expression levels coincide with the morphological transformation of primitive saccules into differentiated alveoli [90][91]. Likewise, RALDH1 and RALDH2 are expressed in a temporal and spatial regulated manner in the lung and are associated with different patterns of alveolar cell wall proliferation. For example, in mice, RALDH1 levels are higher at P4, and it is found in LIFs populating alveolar walls, like CRBPI, and in the bronchial epithelium. Conversely, RALDH2 peaks just before birth, it slowly declines during alveologenesis, and it is expressed in the bronchial epithelium and pleural mesothelial cells but not in the alveolar walls [92]. Secondary septation is RALDH/RA-dependent. Epithelium-derived vascular endothelial growth factor A (VEGFA) regulates RALDH levels and, consequently, RA production by endothelial cells. RA acts as a paracrine factor to induce the expression of FGF18 by lung fibroblasts that, on its turn, regulates elastin deposition [93].
RA and RAR receptors are crucial for septation and secondary crest formation [79]. RARα, RARβ, and RARγ were identified in rat lung isolated LIFs and in postnatal mice lung, namely RARα1, RARβ2, RARβ4, and RARγ2 [92]. RARs are present in the alveolar walls, bronchial epithelium, pleura, and bronchial vascular smooth muscle. Their expression changes with time and, in mouse, increases significantly at P4 [82]. Lungs from rarα−/− mice are similar to wild type at P14 (end of septation), but, at P50, the alveoli number and surface area are decreased. These results hint that RARα is not vital for alveolar septation, but it may have a role in later phases of alveologenesis [94]. Conversely, increased RA signaling of a dominant active RARα receptor in the epithelium leads to lung immaturity [73]. In rarβ−/− mice (exon 10 deletion), septation occurs earlier and faster; also, the administration of RARβ specific agonists in the rat model impairs septation. Overall, it seems that RARβ negatively controls the septation process [95]. However, RARβ seems to be important for neonatal alveolar growth, as disclosed by studying a different rarβ−/− mutant (exon 6 deletion) that displayed a decreased surface area of gas exchange [96]. RARγ is required for alveoli formation during the first 28 postnatal days. rarγ−/− display a decrease in the elastic tissue of the whole lung, and the alveoli number. In contrast, the distance between the alveolar walls and the mean chord length increased. Additionally, in lipofibroblasts, the trophoelastin mRNA decreased at P12 [79]. Moreover, RARγ agonist’s administration promotes alveolar repair in an animal model of emphysema [97]. Altogether, it seems that RARγ positively regulates pulmonary septation. Furthermore, in cultured LIFs, it was demonstrated that RARβ and RARγ increase at birth, which corresponds to the peak of RA storage. The increase in RARγ is temporary since it decreases after P2 (RA in the LIFs also decreases after postnatal day 2). In summary, RA may act at RARγ level in the early postnatal period [92]. RAR heterodimerizes with RXR before binding to DNA. Homozygous null RXR-α mice die in utero. Additionally, null for RARγ and heterozygote for RXR-α present decreased elastin expression in the LIFs at P10; and a decrease in pulmonary elastic tissue in the alveolar septa and not at the airway or vascular walls at P28 [79].
After the administration of retinyl palmitate to pregnant rats, the levels of retinyl esters increase in the fetal and postnatal lung [98], which implies that fetal lungs store retinol or retinyl esters from the progenitor. The lung is the second biggest storing place of retinoids following the liver [99]. LIFs are the main retinoid reservoir of the lung and contain all the components of the pathway [89][99][100]; consequently, they can uptake circulating retinol or hydrolyze stored retinyl esters and convert them into retinol or RA [101][102]. LIFs are localized in the septating tissue adjacent to AEC2 [103]. LIFs generate all-trans retinoic acid (ATRA) that acts autocrinally to upregulate elastin gene expression and synthesis, thus playing a crucial role in the induction of septal eruption [101][104][105]. Moreover, ATRA acts paracrinally to induce proliferation of the adjacent AEC2 [106][107], gene expression [100], and angiogenesis in microvascular endothelial cells [108]. After birth, esters start to disappear from the lung and are converted into active forms, like retinol and RA [83][89][109]. Retinyl esters are abundant in the last three prenatal days (similar amount comparing to the liver) [102], and its depletion correlates with lung maturation [83]. Retinoid clearance is associated with changes in the epithelium of the conducting airways. Specifically, cell phenotype alters from stratified columnar epithelium with goblet cells to keratinizing squamous epithelium [110].
RA regulates positively or negatively the expression of many genes. Among them, RA is important for the synthesis of the surfactant proteins SPA, SPB, and SPC. Administration of retinyl palmitate at high doses to pregnant rats elevates both total phospholipid and the desaturated portion of phosphatidylcholine of the pulmonary surfactant. However, SPA concentration remains unchanged. Additionally, repetitive RA administration increases surfactant phospholipid content [111]. On the other hand, RA also has inhibitory actions. In some studies, it was shown that RA decreases the expression of both mRNA and protein levels of SPA [112].
Regenerative processes are intimately connected with the reawakening of developmental programs [113], and RA has been described as important in alveolar regeneration. Exogenous ATRA, RARα and RARγ agonists and 4-oxo RA can induce alveolar regeneration in adult rats displaying emphysema and partially rescue their phenotype [95][97][114]. On the contrary, pan RXR agonist, 13-cis RA, and retinol were not regeneration inducers, and RARβ was not required for regeneration [115]. In some studies, ATRA was shown to induce apoptosis, a normal process in wound healing, in rats previously treated with elastase, rescuing the alveolar surface area whereas, in others, RA did not affect lung function [116][117]. Recently, using lung organoid models, it was shown that RA pathway stimulation led to a decrease in the organoid size and inhibited epithelial proliferation. In contrast, RA pathway inhibition promoted epithelial proliferation in mice lung organoids and human organoids from chronic obstructive pulmonary disease (COPD) patients. In the mouse model, increased proliferation happened with concomitant suppression of epithelial differentiation from the airway and alveolar epithelium. Furthermore, cell proliferation was intermediated by YAP (yes-associated protein) activation and FGF signaling. Lastly, the inhibition of Histone deacetylase in combination with ATRA was proposed as a potential method to restore adult lung epithelial cell differentiation [118]. An in silico study provided the first clues regarding the possible role of ATRA in minimizing inflammation in elastase-induced emphysema in rat lungs and, therefore, in alveolar epithelium regeneration. It was proposed that ATRA can bind to receptors and ligands of both ERK and JAK-STAT signaling pathways, namely, TNF-α, IL6ST, TNFR1, and IL6. Furthermore, ATRA showed more binding efficiency for TNF-α and IL6ST and can potentially regulate both ERK and JAK-STAT pathways by acting at its first steps. ATRA administration restored lung histology, the proteases/antiproteases balance (imbalance is characteristic of emphysema condition), the levels of inflammatory markers and antioxidants, and the expression of candidate genes of ERK and JAK-STAT. In conclusion, ATRA reduces inflammation and improves alveolar epithelium regeneration in rat lung with emphysema [119].
2.6. Vascular Development
The pulmonary vasculature forms synchronously with the airways by a process called distal angiogenesis [120]. Actually, disruption of lung vascular development impairs airway and alveolar formation, a feature that characterizes some congenital lung diseases [121]. VEGFA plays a crucial role in the interaction between the epithelial and endothelial compartment [122]. On its turn, new endothelial cells are bordered by pericytes that require PDGFβ signaling [123]. As it occurs with airway formation, fine-tuning of multiple signaling pathways is essential for vascular development [121].
RA signaling regulates the proliferation of endothelial cells during vasculogenesis in mice embryos and yolk sacs [124]. In the particular case of the lung, Schmidt et al. [125] suggested that prenatal administration of RA improves lung vascularization and VEGF expression in a rat model of congenital diaphragmatic hernia (CDH). Later in development, during alveologenesis, endothelial cells produce RA that promotes FGF18 expression in adjacent mesenchymal cells, thus regulating elastin production. Simultaneously, RA acts autocrinally controlling endothelial cell proliferation and tube formation [93]. More recently, it has been demonstrated that RA signaling is involved in pericyte migration, angiogenesis, and collagen IV expression [126].
References
- Cunningham, T.J.; Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nat. Rev. Mol. Cell Biol. 2015, 16, 110–123.
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146.
- Timoneda, J.; Rodríguez-Fernández, L.; Zaragozá, R.; Marín, M.P.; Cabezuelo, M.T.; Torres, L.; Viña, J.R.; Barber, T. Vitamin A Deficiency and the Lung. Nutrients 2018, 10, 1132.
- Cardoso, W.V.; Chen, F. Retinoic Acid in the Developing Lung and Other Foregut Derivatives. In The Retinoids: Biology, Biochemistry, and Disease; Dollé, P., Neiderreither, K., Eds.; John Wiley & Sons: IncHoboken, NJ, USA, 2015.
- Quanjer, P.H.; Stanojevic, S.; Cole, T.J.; Baur, X.; Hall, G.L.; Culver, B.H.; Enright, P.L.; Hankinson, J.L.; Ip, M.S.M.; Zheng, J.; et al. Multi-ethnic reference values for spirometry for the 3-95-yr age range: The global lung function 2012 equations. Eur. Respir. J. 2012, 40, 1324–1343.
- Stocks, J.; Hislop, A.; Sonnappa, S. Early lung development: Lifelong effect on respiratory health and disease. Lancet Respir. Med. 2013, 1, 728–742.
- Burri, P.H. Structural Aspects of Postnatal Lung Development–Alveolar Formation and Growth. Biol. Neonate 2006, 89, 313–322.
- Hogan, B.L.M.; Barkauskas, C.E.; Chapman, H.A.; Epstein, J.A.; Jain, R.; Hsia, C.C.W.; Niklason, L.; Calle, E.; Le, A.; Randell, S.H.; et al. Repair and regeneration of the respiratory system: Complexity, plasticity, and mechanisms of lung stem cell function. Cell Stem Cell 2014, 15, 123–138.
- Morrisey, E.E.; Hogan, B.L.M. Preparing for the First Breath: Genetic and Cellular Mechanisms in Lung Development. Dev. Cell 2010, 1, 8–23.
- McCulley, D.; Wienhold, M.; Sun, X. The pulmonary mesenchyme directs lung development. Curr. Opin. Genet. Dev. 2015, 32, 98–105.
- Warburton, D.; Perin, L.; Defilippo, R.; Bellusci, S.; Shi, W.; Driscoll, B. Stem/progenitor cells in lung development, injury repair, and regeneration. Proc. Am. Thorac. Soc. 2008, 5, 703–706.
- Kim, H.Y.; Pang, M.F.; Varner, V.D.; Kojima, L.; Miller, E.; Radisky, D.C.; Nelson, C.M. Localized Smooth Muscle Differentiation Is Essential for Epithelial Bifurcation during Branching Morphogenesis of the Mammalian Lung. Dev. Cell 2015, 34, 719–726.
- Warburton, D.; El-Hashash, A.; Carraro, G.; Tiozzo, C.; Sala, F.; Rogers, O.; De Langhe, S.; Kemp, P.J.; Riccardi, D.; Torday, J.; et al. Chapter Three-Lung Organogenesis. In Current Topics in Developmental Biology; Koopman, P., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 90, pp. 73–158.
- Schittny, J.C. Development of the lung. Cell Tissue Res. 2017, 367, 427–444.
- Herriges, M.; Morrisey, E.E. Lung development: Orchestrating the generation and regeneration of a complex organ. Development 2014, 141, 502–513.
- Fernandes-Silva, H.; Correia-Pinto, J.; Moura, R.S. Canonical Sonic Hedgehog Signaling in Early Lung Development. J. Dev. Biol. 2017, 5, 3.
- Hussain, M.; Xu, C.; Lu, M.; Wu, X.; Tang, L.; Wu, X. Wnt/β-catenin signaling links embryonic lung development and asthmatic airway remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3226–3242.
- Ning, J.; Zhao, Y.; Ye, Y.; Yu, J. Opposing roles and potential antagonistic mechanism between TGF-β and BMP pathways: Implications for cancer progression. EBioMedicine 2019, 41, 702–710.
- Danopoulos, S.; Shiosaki, J.; Al Alam, D. FGF Signaling in Lung Development and Disease: Human Versus Mouse. Front. Genet. 2019, 10, 170.
- Yeung, B.; Yu, J.; Yang, X. Roles of the Hippo pathway in lung development and tumorigenesis. Int. J. Cancer 2016, 138, 533–539.
- Lazzaro, D.; Price, M.; Felice, M.D.E.; Lauro, R.D.I. The transcription factor TTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restricted regions of the foetal brain. Development 1991, 113, 1093–1104.
- Goss, A.M.; Tian, Y.; Tsukiyama, T.; Cohen, E.D.; Zhou, D.; Lu, M.M.; Yamaguchi, T.P.; Morrisey, E.E. Wnt2/2b and β-Catenin Signaling Are Necessary and Sufficient to Specify Lung Progenitors in the Foregut. Dev. Cell 2009, 17, 290–298.
- Domyan, E.T.; Ferretti, E.; Throckmorton, K.; Mishina, Y.; Nicolis, S.K.; Sun, X. Signaling through BMP receptors promotes respiratory identity in the foregut via repression of Sox2. Development 2011, 138, 971–981.
- Chen, F.; Desai, T.J.; Qian, J.; Niederreither, K.; Lü, J.; Cardoso, W.V. Inhibition of Tgfβ signaling by endogenous retinoic acid is essential for primary lung bud induction. Development 2007, 134, 2969–2979.
- Malpel, S.; Mendelsohn, C.; Cardoso, W.V. Regulation of retinoic acid signaling during lung morphogenesis. Development 2000, 127, 3057–3067.
- Desai, T.J.; Malpel, S.; Flentke, G.R.; Smith, S.M.; Cardoso, W.V. Retinoic acid selectively regulates Fgf10 expression and maintains cell identity in the prospective lung field of the developing foregut. Dev. Biol. 2004, 273, 402–415.
- Wang, Z.; Dollé, P.; Cardoso, W.V.; Niederreither, K. Retinoic acid regulates morphogenesis and patterning of posterior foregut derivatives. Dev. Biol. 2006, 297, 433–445.
- Desai, T.J.; Chen, F.; Lü, J.; Qian, J.; Niederreither, K.; Dollé, P.; Chambon, P.; Cardoso, W.V. Distinct roles for retinoic acid receptors alpha and beta in early lung morphogenesis. Dev. Biol. 2006, 291, 12–24.
- Chen, F.; Cao, Y.; Qian, J.; Shao, F.; Niederreither, K.; Cardoso, W.V. A retinoic acid–dependent network in the foregut controls formation of the mouse lung primordium. J. Clin. Investig. 2010, 120, 2040–2048.
- Rankin, S.A.; Han, L.; Kyle, W.; Wells, J.M.; John, M.; Zorn, A.M.; Rankin, S.A.; Han, L.; Mccracken, K.W.; Kenny, A.P.; et al. Article A Retinoic Acid-Hedgehog Cascade Coordinates Mesoderm-Inducing Signals and Endoderm Competence during Lung Specification. Cell Rep. 2016, 16, 66–78.
- Rankin, S.A.; McCracken, K.W.; Luedeke, D.M.; Han, L.; Wells, J.M.; Shannon, J.M.; Zorn, A.M. Timing is everything: Reiterative Wnt, BMP and RA signaling regulate developmental competence during endoderm organogenesis. Dev. Biol. 2018, 434, 121–132.
- Metzger, R.J.; Klein, O.D.; Martin, G.R.; Krasnow, M.A. The branching programme of mouse lung development. Nature 2008, 453, 745–750.
- Chazaud, C.; Dollé, P.; Rossant, J.; Mollard, R. Retinoic acid signaling regulates murine bronchial tubule formation. Mech. Dev. 2003, 120, 691–700.
- Fernandes-Silva, H.; Vaz-Cunha, P.; Barbosa, V.B.; Silva-Gonçalves, C.; Correia-Pinto, J.; Moura, R.S. Retinoic acid regulates avian lung branching through a molecular network. Cell Mol. Life Sci. 2017, 74, 4599–4619.
- Rhinn, M.; Schuhbaur, B.; Niederreither, K.; Dollé, P. Involvement of retinol dehydrogenase 10 in embryonic patterning and rescue of its loss of function by maternal retinaldehyde treatment. Proc. Natl. Acad. Sci. USA 2011, 108, 16687–16692.
- Bouillet, P.; Sapin, V.; Chazaud, C.; Messaddeq, N.; Décimo, D.; Dollé, P.; Chambon, P. Developmental expression pattern of Stra6, a retinoic acid-responsive gene encoding a new type of membrane protein. Mech. Dev. 1997, 63, 173–186.
- Dollé, P.; Ruberte, E.; Leroy, P.; Morriss-Kay, G.; Chambon, P. Retinoic acid receptors and cellular retinoid binding proteins. I. A systematic study of their differential pattern of transcription during mouse organogenesis. Development 1990, 110, 1133–1151.
- Bellusci, S.; Grindley, J.; Emoto, H.; Itoh, N.; Hogan, B.L. Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 1997, 124, 4867–4878.
- Niederreither, K.; Fraulob, V.; Garnier, J.M.; Chambon, P.; Dollé, P. Differential expression of retinoic acid-synthesizing (RALDH) enzymes during fetal development and organ differentiation in the mouse. Mech. Dev. 2002, 110, 165–171.
- Mendelsohn, C.; Lohnes, D.; Décimo, D.; Lufkin, T.; LeMeur, M.; Chambon, P.; Mark, M. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994, 120, 2749–2771.
- Mollard, R.; Ghyselinck, N.B.; Wendling, O.; Chambon, P.; Mark, M. Stage-dependent responses of the developing lung to retinoic acid signaling. Int. J. Dev. Biol. 2000, 44, 457–462.
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dollé, P.; Dierich, A.; Gorry, P.; Gansmuller, A.; Chambon, P. Function of the retinoic acid receptors (RARs) during development (I). Craniofacial and skeletal abnormalities in RAR double mutants. Development 1994, 120, 2723–2748.
- Lü, J.; Qian, J.; Izvolsky, K.I.; Cardoso, W.V. Global analysis of genes differentially expressed in branching and non-branching regions of the mouse embryonic lung. Dev. Biol. 2004, 273, 418–435.
- Moura, R.S.; Coutinho-Borges, J.P.; Pacheco, A.P.; Damota, P.O.; Correia-Pinto, J. FGF signaling pathway in the developing chick lung: Expression and inhibition studies. PLoS ONE 2011, 6, e17660.
- Schuger, L.; Varani, J.; Mitra, R., Jr.; Gilbride, K. Retinoic acid stimulates mouse lung development by a mechanism involving epithelial-mesenchymal interaction and regulation of epidermal growth factor receptors. Dev. Biol. 1993, 159, 462–473.
- Montedonico, S.; Nakazawa, N.; Puri, P. Retinoic acid rescues lung hypoplasia in nitrofen-induced hypoplastic foetal rat lung explants. Pediatr. Surg. Int. 2006, 22, 2–8.
- Pereira-Terra, P.; Moura, R.S.; Nogueira-Silva, C.; Correia-Pinto, J. Neuroendocrine factors regulate retinoic acid receptors in normal and hypoplastic lung development. J. Physiol. 2015, 593, 3301–3311.
- Cardoso, W.V.; Williams, M.C.; Mitsialis, S.A.; Joyce-Brady, M.; Rishi, A.K.; Brody, J.S. Retinoic acid induces changes in the pattern of airway branching and alters epithelial cell differentiation in the developing lung in vitro. Am. J. Respir. Cell Mol. Biol. 1995, 12, 464–476.
- Cardoso, W.V.; Mitsialis, S.A.; Brody, J.S.; Williams, M.C. Retinoic acid alters the expression of pattern-related genes in the developing rat lung. Dev. Dyn. 1996, 207, 47–59.
- Urase, K.; Mukasa, T.; Igarashi, H.; Ishii, Y.; Yasugi, S.; Momoi, M.Y.; Momoi, T. Spatial expression of Sonic hedgehog in the lung epithelium during branching morphogenesis. Biochem. Biophys. Res. Commun. 1996, 225, 161–166.
- Lebeche, D.; Malpel, S.; Cardoso, W.V. Fibroblast growth factor interactions in the developing lung. Mech. Dev. 1999, 86, 125–136.
- Weaver, M.; Yingling, J.M.; Dunn, N.R.; Bellusci, S.; Hogan, B.L. Bmp signaling regulates proximal-distal differentiation of endoderm in mouse lung development. Development 1999, 126, 4005–4015.
- Zhou, L.; Dey, C.R.; Wert, S.E.; Yan, C.; Costa, R.H.; Whitsett, J.A. Hepatocyte nuclear factor-3beta limits cellular diversity in the developing respiratory epithelium and alters lung morphogenesis in vivo. Dev. Dyn. 1997, 210, 305–314.
- Bogue, C.W.; Gross, I.; Vasavada, H.; Dynia, D.W.; Wilson, C.M.; Jacobs, H.C. Identification of Hox genes in newborn lung and effects of gestational age and retinoic acid on their expression. Am. J. Physiol. 1994, 266, L448–L454.
- Volpe, M.V.; Vosatka, R.J.; Nielsen, H.C. Hoxb-5 control of early airway formation during branching morphogenesis in the developing mouse lung. Biochim. Biophys. Acta 2000, 1475, 337–345.
- Chen, F.; Marquez, H.; Kim, Y.K.; Qian, J.; Shao, F.; Fine, A.; Cruikshank, W.W.; Quadro, L.; Cardoso, W.V. Prenatal retinoid deficiency leads to airway hyperresponsiveness in adult mice. J. Clin. Investig. 2014, 124, 801–811.
- Gontan, C.; de Munck, A.; Vermeij, M.; Grosveld, F.; Tibboel, D.; Rottier, R. Sox2 is important for two crucial processes in lung development: Branching morphogenesis and epithelial cell differentiation. Dev. Biol. 2008, 317, 296–309.
- Rockich, B.; Hrycaj, S.M.; Shih, H.P.; Nagy, M.S.; Ferguson, M.A.; Kopp, J.L.; Sander, M.; Wellik, D.M.; Spence, J.R. Sox9 plays multiple roles in the lung epithelium during branching morphogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E4456–E4464.
- deMello, D.E.; Reid, L.M. Embryonic and early fetal development of human lung vasculature and its functional implications. Pediatr. Dev. Pathol. 2000, 3, 439–449.
- Ornitz, D.M.; Yin, Y. Signaling networks regulating development of the lower respiratory tract. Cold Spring Harb. Perspect. Biol. 2012, 4.
- Boyden, E.A. The structure of the pulmonary acinus in a child of six years and eight months. Am. J. Anat. 1971, 132, 275–299.
- Kitaoka, H.; Burri, P.H.; Weibel, E.R. Development of the human fetal airway tree: Analysis of the numerical density of airway endtips. Anat. Rec. 1996, 244, 207–213.
- DiMaio, M.; Gil, J.; Ciurea, D.; Kattan, M. Structural maturation of the human fetal lung: A morphometric study of the development of air-blood barriers. Pediatr. Res. 1989, 26, 88–93.
- McQualter, J.L.; Yuen, K.; Williams, B.; Bertoncello, I. Evidence of an epithelial stem/progenitor cell hierarchy in the adult mouse lung. Proc. Natl. Acad. Sci. USA 2010, 107, 1414–1419.
- Nolen-Walston, R.D.; Kim, C.F.; Mazan, M.R.; Ingenito, E.P.; Gruntman, A.M.; Tsai, L.; Boston, R.; Woolfenden, A.E.; Jacks, T.; Hoffman, A.M. Cellular kinetics and modeling of bronchioalveolar stem cell response during lung regeneration. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, 1158–1166.
- Barré, S.F.; Haberthür, D.; Cremona, T.P.; Stampanoni, M.; Schittny, J.C. The total number of acini remains constant throughout postnatal rat lung development. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L1082–L1089.
- DeMello, D.E.; Sawyer, D.; Galvin, N.; Reid, L.M. Early Fetal Development of Lung Vasculature. Am. J. Respir. Cell Mol. Biol. 1997, 16, 568–581.
- Vu, T.H.; Alemayehu, Y.; Werb, Z. New insights into saccular development and vascular formation in lung allografts under the renal capsule. Mech. Dev. 2003, 120, 305–313.
- Grummer, M.A.; Thet, L.A.; Zachman, R.D. Expression of retinoic acid receptor genes in fetal and newborn rat lung. Pediatr. Pulmonol. 1994, 17, 234–238.
- Pawlikowski, B.; Wragge, J.; Siegenthaler, J.A. Retinoic acid signaling in vascular development. Genesis 2019, 57, e23287.
- Kumar, V.H.; Lakshminrusimha, S.; El, Abiad, M.T.; Chess, P.R.; Ryan, R.M. Growth factors in lung development. Adv. Clin. Chem. 2005, 40, 261–316.
- Nikolić, M.Z.; Sun, D.; Rawlins, E.L. Human lung development: Recent progress and new challenges. Development 2018, 145.
- Wongtrakool, C.; Malpel, S.; Gorenstein, J.; Sedita, J.; Ramirez, M.I.; Underhill, T.M.; Cardoso, W.V. Down-regulation of retinoic acid receptor alpha signaling is required for sacculation and type I cell formation in the developing lung. J. Biol. Chem. 2003, 278, 46911–46928.
- Massaro, G.D. Formation of Pulmonary Alveoli and Gas-Exchange Surface Area: Quantitation and Regulation. Annu. Rev. Physiol. 1996, 58, 73–92.
- Boström, H.; Willetts, K.; Pekny, M.; Levéen, P.; Lindahl, P.; Hedstrand, H.; Pekna, M.; Hellström, M.; Gebre-Medhin, S.; Schalling, M.; et al. PDGF-A signaling is a critical event in lung alveolar myofibroblast development and alveogenesis. Cell 1996, 85, 863–873.
- Lindahl, P.; Karlsson, L.; Hellström, M.; Gebre-Medhin, S.; Willetts, K.; Heath, J.K.; Betsholtz, C. Alveogenesis failure in PDGF-A-deficient mice is coupled to lack of distal spreading of alveolar smooth muscle cell progenitors during lung development. Development 1997, 124, 3943–3953.
- Burri, P.H. Postnatal growth and maturation of the lung. Chest 1975, 67, 2S–3S.
- Caduff, J.H.; Fischer, L.C.; Burri, P.H. Scanning electron microscope study of the developing microvasculature in the postnatal rat lung. Anat. Rec. 1986, 216, 154–164.
- McGowan, S.; Jackson, S.K.; Jenkins-Moore, M.; Dai, H.H.; Chambon, P.; Snyder, J.M. Mice bearing deletions of retinoic acid receptors demonstrate reduced lung elastin and alveolar numbers. Am. J. Respir. Cell Mol. Biol. 2000, 23, 162–167.
- Weinstein, M.; Xu, X.; Ohyama, K.; Deng, C.X. FGFR-3 and FGFR-4 function cooperatively to direct alveogenesis in the murine lung. Development 1998, 125, 3615–3623.
- Massaro, G.D.; Massaro, D. Treatment with retinoic acid increases the number of pulmonary alveoli. FASEB J. 1996, 270, L305–L310.
- Hind., M.; Corcoran., J.; Maden., M. Temporal/spatial expression of retinoid binding proteins and RAR isoforms in the postnatal lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L468–L476.
- Geevarghese, S.K.; Chytil, F. Depletion of retinyl esters in the lungs coincides with lung prenatal morphological maturation. Biochem. Biophys. Res. Commun. 1994, 200, 529–535.
- McGowan, S.E. Contributions of retinoids to the generation and repair of the pulmonary alveolus. Chest 2002, 121, 206S–208S.
- Baybutt, R.C.; Hu, L.; Molteni, A. Vitamin A Deficiency Injures Lung and Liver Parenchyma and Impairs Function of Rat Type II Pneumocytes. J. Nutr. 2000, 130, 1159–1165.
- Biesalski, H.K.; Nohr, D. Importance of vitamin-A for lung function and development. Mol. Asp. Med. 2003, 24, 431–440.
- Verma, R.P.; Mcculloch, K.M.; Worrell, L.; Vidyasagar, D. Vitamin A deficiency and severe bronchopulmonary dysplasia in very low birthweight infants. Am. J. Perinatol. 1996, 13, 389–393.
- Albertine, K.H.; Jones, G.P.; Starcher, B.C.; Bohnsack, J.F.; Davis, P.L.; Cho, S.C.; Carlton, D.P.; Bland, R.D. Chronic lung injury in preterm lambs: Disordered respiratory tract development. Am. J. Respir. Crit. Care Med. 1999, 159, 945–958.
- McGowan, S.E.; Harvey, C.S.; Jackson, S.K. Retinoids, retinoic acid receptors, and cytoplasmic retinoid binding proteins in perinatal rat lung fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 1995, 269, L463–L472.
- Powell, J.T.; Whitney, P.L. Postnatal development of rat lung. Changes in lung lectin, elastin, acetylcholinesterase and other enzymes. Biochem. J. 1980, 188, 1–8.
- Ong, D.E.; Chytil, F. Changes in levels of cellular retinol and retinoic acid binding proteins of liver and lung during perinatal development of rat. Proc. Natl. Acad. Sci. USA 1976, 73, 3976–3978.
- Hind, M.; Corcoran, J.; Maden, M. Alveolar proliferation, retinoid synthesizing enzymes, and endogenous retinoids in the postnatal mouse lung different roles for Aldh-1 and Raldh-2. Am. J. Respir Cell Mol. Biol. 2002, 26, 67–73.
- Yun, E.J.; Lorizio, W.; Seedorf, G.; Abman, S.H.; Vu, T.H. VEGF and endothelium-derived retinoic acid regulate lung vascular and alveolar development. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L287–L298.
- Massaro, G.D.; Massaro, D.; Chambon, P. Retinoic acid receptor-α regulates pulmonary alveolus formation in mice after, but not during, perinatal period. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 284, 2002–2004.
- Massaro, G.D.; Massaro, D.; Chan, W.Y.; Clerch, L.B.; Ghyselinck, N.; Chambon, P.; Chandraratna, R.A.S. Retinoic acid receptor-β: An endogenous inhibitor of the perinatal formation of pulmonary alveoli. Physiol. Genom. 2001, 4, 51–57.
- Snyder, J.M.; Jenkins-Moore, M.; Jackson, S.K.; Goss, K.L.; Dai, H.H.; Bangsund, P.J.; Giguere, V.; Mcgowan, S.E. Alveolarization in retinoic acid receptor-β-deficient mice. Pediatr. Res. 2005, 57, 384–391.
- Massaro, G.D.; Massaro, D. Retinoic acid treatment abrogates elastase-induced pulmonary emphysema in rats. Nat. Med. 1997, 3, 675–677.
- Shenai, J.P.; Chytil, F. Effect of maternal vitamin-A administration on fetal lung vitamin-A stores in the perinatal rat. Biol. Neonate 1990, 58, 318–325.
- Okabe, T.; Yorifuji, H.; Yamada, E.; Takaku, F. Isolation and characterization of vitamin-A-storing lung cells. Exp. Cell Res. 1984, 154, 125–135.
- Dirami, G.; Massaro, G.D.; Clerch, L.B.; Ryan, U.S.; Reczek, P.R.; Massaro, D. Lung retinol storing cells synthesize and secrete retinoic acid, an inducer of alveolus formation. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 286, 249–256.
- McGowan, S.E.; Torday, J.S. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu. Rev. Physiol. 1997, 59, 43–62.
- Shenai, J.P.; Chytil, F. Vitamin A storage in lungs during perinatal development in the rat. Biol. Neonate 1990, 57, 126–132.
- Burri, P.H. The postnatal growth of the rat lung III. Morphology. Anat. Rec. 1974, 180, 77–98.
- Mcgowan, S.E.; Doro, M.M.; Jackson, S.K. Endogenous retinoids increase perinatal elastin gene expression in rat lung fibroblasts and fetal explants. Am. J. Physiol. Lung Cell Mol. Physiol. 1997, 273, L410–L416.
- Liu, R.; Harvey, C.S.; McGowan, S.E. Retinoic acid increases elastin in neonatal rat lung fibroblast cultures. Am. J. Physiol. Lung Cell Mol. Physiol. 1993, 265, L430–L437.
- Nabeyrat, E.; Besnard, V.; Corroyer, S.; Cazals, V.; Clement, A. Retinoic acid-induced proliferation of lung alveolar epithelial cells: Relation with the IGF system. Am. J. Physiol. Lung Cell Mol. Physiol. 1998, 275, 71–79.
- Nabeyrat, E.; Corroyer, S.; Epaud, R.; Besnard, R.I.E.; Corroyer, S.; Epaud, R. Retinoic acid-induced proliferation of lung alveolar epithelial cells is linked to p21(CIP1) downregulation. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 278, L42–L50.
- Ng-Blichfeldt, J.P.; Alçada, J.; Montero, M.A.; Dean, C.H.; Griesenbach, U.; Griffiths, M.J.; Hind, M. Deficient retinoid-driven angiogenesis may contribute to failure of adult human lung regeneration in emphysema. Thorax 2017, 72, 510–521.
- Zachman, R.D.; Kakkad, B.; Chytil, F. Perinatal rat lung retinol (vitamin A) and retinyl palmitate. Pediatr. Res. 1984, 18, 1297–1299.
- Wolbach, S.B.; Howe, P.R. Tissue Changes Following Deprivation of Fat-Soluble A Vitamin. J. Exp. Med. 1925, 42, 753–791.
- Fraslon, C.; Bourbon, J.R. Retinoids control surfactant phospholipid biosynthesis in fetal rat lung. Am. J. Physiol. Lung Cell Mol. Physiol. 1994, 266, L705–L712.
- Metzler, M.D.; Snyder, J.M. Retinoic acid differentially regulates expression of surfactant-associated proteins in human fetal lung. Endocrinology 1993, 133, 1990–1998.
- Maden, M.; Hind, M. Retinoic acid in alveolar development, maintenance and regeneration. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 799–808.
- Hind, M.; Maden, M. Retinoic acid induces alveolar regeneration in the adult mouse lung. Eur. Respir J. 2004, 23, 20–27.
- Maden, M. Retinoids have differing efficacies on alveolar regeneration in a dexamethasone-treated mouse. Am. J. Respir Cell Mol. Biol. 2006, 35, 260–267.
- Belloni, P.N.; Garvin, L.; Mao, C.P.; Bailey-Healy, I.; Leaffer, D. Effects of all-trans-retinoic acid in promoting alveolar repair. Chest 2000, 117, 235S–241S.
- Srinivasan, G.; Bruce, E.N.; Houtz, P.K.; Bruce, M.C. Dexamethasone-induced changes in lung function are not prevented by concomitant treatment with retinoic acid. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, 275–287.
- Ng-Blichfeldt, J.P.; Gosens, R.; Dean, C.; Griffiths, M.; Hind, M. Regenerative pharmacology for COPD: Breathing new life into old lungs. Thorax 2019, 74, 890–897.
- Uniyal, S.; Dhasmana, A.; Tyagi, A.; Muyal, J.P. ATRA reduces inflammation and improves alveolar epithelium regeneration in emphysematous rat lung. Biomed. Pharmacother. 2018, 108, 1435–1450.
- Parera, M.C.; Van Dooren, M.; Van Kempen, M.; De Krijger, R.; Grosveld, F.; Tibboel, D.; Rottier, R. Distal angiogenesis: A new concept for lung vascular morphogenesis. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, 141–149.
- Kool, H.; Mous, D.; Tibboel, D.; de Klein, A.; Rottier, R.J. Pulmonary vascular development goes awry in congenital lung abnormalities. Birth Defects Res. Part. C Embryo Today Rev. 2014, 102, 343–358.
- Yamamoto, H.; Jun Yun, E.; Gerber, H.P.; Ferrara, N.; Whitsett, J.A.; Vu, T.H. Epithelial-vascular cross talk mediated by VEGF-A and HGF signaling directs primary septae formation during distal lung morphogenesis. Dev. Biol. 2007, 308, 44–53.
- Armulik, A.; Abramsson, A.; Betsholtz, C. Endothelial/pericyte interactions. Circ. Res. 2005, 97, 512–523.
- Lai, L.; Bohnsack, B.L.; Niederreither, K.; Hirschi, K.K. Retinoic acid regulates endothelial cell proliferation during vasculogenesis. Development 2003, 130, 6465–6474.
- Schmidt, A.F.; Gonçalves, F.L.L.; Regis, A.C.; Gallindo, R.M.; Sbragia, L. Prenatal retinoic acid improves lung vascularization and VEGF expression in CDH rat. Am. J. Obs. Gynecol. 2012, 207, 76.
- Kool, H.M.; Bürgisser, P.E.; Edel, G.G.; de Kleer, I.; Boerema-de Munck, A.; de Laat, I.; Chrifi, I.; Cheng, C.; van Cappellen, W.A.; Kremers, G.J.; et al. Inhibition of retinoic acid signaling induces aberrant pericyte coverage and differentiation resulting in vascular defects in congenital diaphragmatic hernia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 317, 317–331.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
903
Revisions:
2 times
(View History)
Update Date:
23 Jun 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No