Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Helen H.W. Chen | + 2858 word(s) | 2858 | 2021-06-16 06:24:01 |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Chen, H.H.; Kuo, M. Copper Chelators and cDDP Resistance. Encyclopedia. Available online: https://encyclopedia.pub/entry/11046 (accessed on 26 June 2026).
Chen HH, Kuo M. Copper Chelators and cDDP Resistance. Encyclopedia. Available at: https://encyclopedia.pub/entry/11046. Accessed June 26, 2026.
Chen, Helen H.w., Macus Kuo. "Copper Chelators and cDDP Resistance" Encyclopedia, https://encyclopedia.pub/entry/11046 (accessed June 26, 2026).
Chen, H.H., & Kuo, M. (2021, June 21). Copper Chelators and cDDP Resistance. In Encyclopedia. https://encyclopedia.pub/entry/11046
Chen, Helen H.w. and Macus Kuo. "Copper Chelators and cDDP Resistance." Encyclopedia. Web. 21 June, 2021.
Copy Citation
The platinum (Pt)-containing antitumor drugs including cisplatin (cis-diamminedichloroplatinum II, cDDP), carboplatin, and oxaliplatin, have been the mainstay of cancer chemotherapy. These drugs are effective in treating many human malignancies. The major cell-killing target of Pt drugs is DNA. Recent findings underscored the important roles of Pt drug transport system in cancer therapy. It was demonstrated that by reducing cellular Cu bioavailable levels by Cu chelators, hCtr1 is transcriptionally upregulated by transcription factor Sp1, which binds the promoters of Sp1 and hCtr1. In contrast, elevated Cu poisons Sp1, resulting in suppression of hCtr1 and Sp1, constituting the Cu-Sp1-hCtr1 mutually regulatory loop.
cisplatin
carboplatin
oxaliplatin
copper transporter
Ctr1
Atox1
ATP7A. ATP7B. Sp1
copper chelator
1. Introduction
Platinum (Pt)-based antitumor agents, including cisplatin (cis-diamminedichloroplatinum II, cDDP), carboplatin, and oxaliplatin, are active against many tumor types. cDDP is commonly used for treating metastatic testicular cancer, carboplatin for advanced ovarian cancer, and oxaliplatin for advanced colorectal cancer. cDDP has also been used for treating other cancers, including cancers of lung, bladder, head, and neck, and uterine cervix [1][2]. The use of cDDP is limited by its adverse side effects including nephrotoxicity [3][4], ototoxicity [4], and peripheral neurotoxicity. These toxicities are not associated with carboplatin, which mainly induces myelosuppression; whereas the most common toxicity associated with oxaliplatin is peripheral neuropathy [5].
The cytotoxic target of these drugs is DNA, by forming primarily intra-stranded crosslinks d(GpG) adducts. If not repaired, these lesions will damage DNA replication, transcription, and associated cellular functions, resulting in cell death or apoptosis. Previous work demonstrated that the overall lethality of drugs is positively correlated with the cellular contents of Pt drugs [6]. Moreover, it has been reported that reduced Pt levels are the hallmark of Pt-drug resistance [7]. These findings strongly suggest that Pt drug transport mechanisms play important roles in cell-killing of Pt-based antitumor agents.
2. The Transport Mechanisms of Pt Drugs
Several mechanisms have been proposed for Pt drug delivery into cancer cells. Early studies suggested that cDDP enters cells by simple diffusion and independent of membrane protein carrier [8]. Later, using genome-wide loss-of-function screening of a haploid cell line, it was identified that loss of subunits leucine-rich repeat-containing protein (LRRC8A) and LRRC8D of the heteromeric LRRC8 volume-regulated anion channels (VRACs) increased resistance to cDDP and carboplatin but not oxaliplatin [9], perhaps due to structural differences among these drugs. Analyses of The Cancer Genome Atlas (TCGA) database showed that low LRRC8D expression correlates with reduced survival of Pt drug-treated ovarian cancer patients. VRACs are important regulators that control cellular volume-associated chloride and organic osmolytes movements across the cell membrane [10]. Moreover, incorporation of LRRC8D subunit into VRAC enhanced permeability for cDDP. Furthermore, it was reported that cDDP accumulation correlates with LRRC8A protein level and channel activity [11]. However, a recent study reported that knockdown or overexpression of LRRC8A and LRRC8D in a lung adenocarcinoma cell model did not affect cDDP resistance or sensitivity [12]. Elucidation of detailed mechanistic basis underlying how VRAC-mediated cDDP permeation into cells may resolve the discrepancies.
Multiple transporters have been reported for oxaliplatin, i.e., the organic cation transporter system (SLC22A), including OCT2 (SLC22A2) [13] and OCT3 (SLC22A3) [14], organic cation/carnitine transporters OCTNs (OCTN1/OCTN2) [15], and multidrug resistance protein 4 (MRP4) [16]. OCTs are a group of poly-specific transporters for subtracts typically of positively charged or zwitterions at physiological pH conditions, e.g., organic amines choline, neurotransmitters dopamine and serotonin, and vitamin B (thiamine) [17]. In one study, it was reported that long progression-free survival (PFS) in metastatic colon cancer treated with oxaliplatin is linked to high expression of OCT2 [18], implicating the role of this transporter in oxaliplatin treatment.
3. Connecting the Essential Trace Element Cu to Pt-Based Antitumor Drugs
3.1. Identification of the High-Affinity Copper Transporter (Ctr1) as cDDP Importer
Ishida et al. [19] using mutagenesis screening identified that the high-affinity copper transporter (Ctr1) is a cDDP uptake transporter in the yeast. These authors also demonstrated that disruption of yCtr1 showed reduced accumulation of cDDP and increased cDDP resistance. Similar results were found in murine cell line with deletion of the mCtr1 alleles [19].
Ctr1 is an evolutionarily conserved Cu(I) ion transporter from yeast to humans. The hCtr1 (SLC31A1) encodes a polypeptide of 190 amino acids, organized in three transmembrane domains with the N-terminus extracellularly located. Natural Cu exists in oxidized form i.e., Cu(II). It has to be reduced by reductases to Cu(I) for hCtr1 transport and the subsequent intracellular distributions and final export. Unlike yeast Ctr1, which uses iron-copper reductase Fre1p/2p [20], human membrane-associated reductase is not known. However, using model peptides, it was suggested that hCtr1 ATCUN (Amino Terminal Cu(II)- and Ni(II))-binding site located at the extracellular N-terminus and its adjacent bis-His sequences capture extracellular Cu(II) and converts it into Cu(I) in the presence of ascorbate [21]. However, whether ATCUN motif is involved in cellular hCtr1-mediated Cu(I) transport is unknown.
Biochemical analyses identified three highly conserved methionine (Met or M)-rich motifs (7MxMxxM, 40MMMMxM, and 150Mxxx154M) in hCtr1. The first two Met-rich motifs are located at the extracellular N-terminus, whereas the last Met-rich motif is located at the end of the second transmembrane domain (TM2) [22]. Site-directed mutagenesis revealed that these amino acids are also important for cDDP transport [23]. Two dimensional electron crystallographic data [24] revealed that a functional hCtr1 consists of three monomers in an ion channel-like structure with an ~8 Å distance in the central pore. Recently, X-ray crystal structure determination of a genetically engineered fish (Salom salar) sCtr1 revealed two residues in TM2 (M146 and M150), which are critical for Cu(I) binding through Cu-S coordination. These two Met-residuals are also critical for the metal specificity, i.e., Cu(I) vs. Cu(II), Na+, and K+, and for ion (substrate) passing [25].
3.2. Cu Chaperones in Intracellular cDDP Trafficking
Intracellular cDDP trafficking to different subcellular compartments generally follows the paths of Cu(I) distributions carried out by Cu chaperons, i.e., by cytochrome C oxidase copper chaperone Cox17 to mitochondrion, by superoxide dismutase 1 (SOD1) carrier (CCS) to cytoplasmic SOD1, and by antioxidant protein 1 (Atox1) to Cu-efflux pumps ATP7A and ATP7B, the two P-type ATPases located at trans-Golgi network (TGN) (Figure 1). Transcriptome analysis revealed that many of these Cu transporter proteins are coordinately regulated in cancer cells [26]. Atox1 is particularly relevant because it shuffles cDDP to the ATP7A/ATP7B efflux pumps. Atox1 is a soluble protein of 68 amino acids, which captures Cu(I) by directly interacting with the C-terminal 188HCH end of hCtr1. Atox1 coordinates one Cu(I) ion with the cysteine (Cys15) residues of the conserved 12CXX15C motif in Atox1 dimerization [27][28][29].
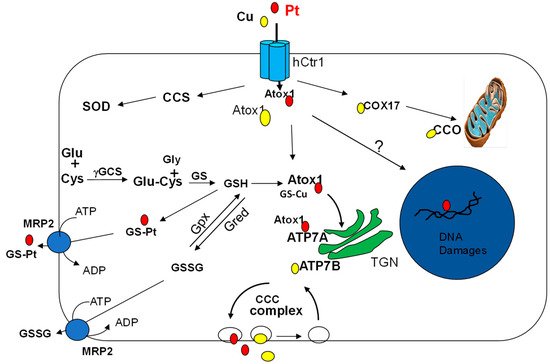
Figure 1. Schematic illustration of copper and platinum entrance, distribution, and elimination in tumor cell. Copper (Cu) and cDDP (Pt) entering cell via Ctr1 are transferred to Chaperones Atox1 and then to ATP7A/ATP7B at the Trans-Golgi Network (TGN) in the cytosol via the help of glutathione (GSH). GSH is synthesized via two sequential enzymatic reactions: ligation of glutamine (Glu) and cysteine (Cys) by γ-glutamylcysteine synthetase (γ-GCS), followed by the addition of glycine (Gly) by glutathione synthetase (GS). Interaction of ATP7A with CCC complex, which consists of COMMD/CCDC22/CCDC93 nodule to eliminate Cu or Pt out of the cell. Cu(I) and Pt can be transferred to mitochondrial cytochrome oxidase (CCO) or to Cu-Zn-superoxidase dismutase (SOD) via chaperones Cox17 and CCS, respectively. GSH and cDDP can form Pt(GS)2, which is eliminated by multidrug protein (MRP2) efflux pump. GSH is oxidized to GSSG by GSH peroxidase (Gpx) and GSSG is reduced back to GSH by GSH reductase (Gred). GSSG can be eliminated by MRP2 using ATP hydrolysis as an energy source.
3.3. Cu-ATPases in Pt Drugs Efflux
The two P-type ATPases ATP7A and ATP7B are well-known cellular Cu(I) exporters. ATP7A is mainly expressed in the intestinal epithelium for Cu absorption from food. ATP7A deletion results in systemic Cu deficiency that causes the Menkes’ disease. ATP7B is mainly located in the livers. Mutations of ATP7B result in massive Cu accumulation in the livers, resulting in Wilson’s disease [30]. Cell culture model revealed that ATP7A deletion enhances ATP7B expression, suggesting a compensatory expression between ATP7A and ATP7B [31].
The human ATP7A and ATP7B contain 1500 and 1465 amino acids, respectively, each organizing in eight transmembrane domains. The N-termini of ATP7A and ATP7B contain six metal-binding domains (MBD), each contains a CXXC motif that receives Cu(I) or cDDP delivered by Atox1. The folding of these CXXC MBD shares similarity with that in Atox1 that is believed to facilitate rapid intermolecular metal transfer between Atox1 and ATPases [32]. Moreover, the conserved amino acid 60lysine in Atox1 is critical for hetero-protein interactions between Atox1 and ATPases for processing Cu(I) transfer [32].
3.4. Redox Regulation of the Cu Transport System in Pt Drug Pharmacology
Copper is a redox-sensitive element, therefore, its physiology is greatly affected by cellular redox conditions. Copper is also an essential element for cell survival and growth, but excess Cu (normally exceeding 10 µM [33]) is detrimental. Therefore, cellular Cu levels must be exquisitely regulated. Cu catalyzes the Fenton reactions, which involve iron-mediated conversion of H2O2 to hydroxyl radicals [34]; both are important reactive oxygen species (ROS). ROS induce oxidative damage of nucleic acids, proteins and lipids. Excessive ROS is lethal, therefore, Cu homeostasis has to be constantly under the control of redox conditions.
One of the most abundant physiological antioxidants that regulates intracellular Cu redox conditions is glutathione (GSH). GSH is a tripeptide synthesized by sequential reactions involving glutamine (Glu) and cysteine (Cys) conjugation catalyzed by γ-glutamylcysteine synthesis (γGCS) followed by the addition of glycine (Gly) by glutathione synthetase (GS) (Figure 1). GSH is oxidized to GSSG by glutathione peroxidase (Gpx), and GSSG is reduced back to GSH by glutathione reductase (Gred). Ratios of GSH and GSSG mostly reflect the cellular redox conditions, which are variable in different cellular compartments.
Cellular GS and Gpx activities are critical in regulating GSH/GSSG ratios. Human GS is a homodimeric enzyme encoded by a single gene, whereas Gpx consists of a family of at least seven isozymes encoded by genes located on different chromosomes. Different isoforms have different tissue expression patterns [35].
4. Modulations of Copper Homeostasis and Pt Drug Cancer Chemotherapy
4.1. Roles of the Copper Transport System in Pt Drug Cancer Chemotherapy in Clinical Settings
4.1.1. hCtr1
Ishida et al. [36] analyzed an array-based hCtr1 expression dataset consisting of 91 patients with stage III or IV serous epithelial ovarian cancer who had been treated with a cytoreductive surgery followed by adjuvant chemotherapy of a Pt drug and a taxane deposited in TCGA. It was observed that elevated expression of hCtr1 was associated with increased disease-free survival. Similar results were obtained by analyzing an independent data set consisting of 285 patients [23]. Furthermore, in a meta-analysis of 12 studies with eight datasets in TCGA, it revealed that high hCtr1expression was significantly associated with a favorable overall survival (OS), progression-free survival (PFS), disease-free survival (DFS), and treatment response [37]. These results, collectively, strongly suggest that hCtr1 expression levels are correlated with cDDP efficacies in clinical settings.
cDDP has achieved a remarkable cure rate in advanced testicular cancer treatment [38]. The clinical mechanisms were investigated in animal models. Testis actively produces sperms in male fertilization. Spermatogenesis takes place in seminiferous epithelium, which consists of germ cells (GC) and Sertoli cells (SC). The somatic SC plays supportive roles in protection, nutrition, and proliferation of GC. Murine mCtr1 is highly expressed in SC [39], suggesting that mCtr1 in SC may contribute to the cDDP-induced testicular cell-killing. Indeed, SC-specific knock out of mCtr1 resulted in reduced sensitivity of testicular germ cells to cDDP treatment [40].
4.1.2. Atox1, ATP7A and ATP7B
It has been reported that deletion of Atox1 resulted in inability of cDDP delivery to ATP7A/ATP7B [41][42]. This may be associated with increased cDDP sensitivity. As for the roles of ATP7A and ATP7B, many studies have demonstrated that elevated expression of ATP7A and ATP7B is associated with poor outcomes of Pt-based cancer chemotherapy in cancers of ovary, breast, and lung (see review in [43] and references therein). In other studies, overexpression of ATP7A and ATP7B was associated with resistance to cDDP [44][45][46], carboplatin [47] or oxliplatin [48]. Likewise, elevated ATP7B levels were correlated with worse treatment outcomes of cDDP in various cancers [46][49].
4.1.3. COMMD1
COMMD1 is a multifunctional protein that interacts with many other proteins [50]. While the majority of COMMD1 is cytosolic, a small fraction is present in the nucleus. It was reported that elevated levels of nuclear COMMD1 confer sensitivity of ovarian cancer cells to Pt-based chemotherapy. Consistent with this in vitro finding, it was reported that increased expression of COMMD1 in the nucleus of ovarian tumors is associated with improved response to cDDP therapy [51].
4.2. Enhanced cDDP Cell-Killing Activity through Upregulation of hCtr1 Expression
The above observations indicate that almost all, if not the entire, processes of the copper transport system are involved in Pt-drug chemosensitivity regulation. These observations also suggest that targeting the Cu transport system may be an effective approach to improve Pt drug cancer therapy. In this review, we will focus on targeting hCtr1 expression.
It is important to note that the majority of cellular Cu is bound by cellular constituents and only a small fraction is free Cu for exchange. For example, about 90% of Cu in the circulation is bound by ceruloplasmin and the other 10% is bound by albumin, histidine, GSH, and transferrin [52][53]. At the intracellular level, in the entire process of Cu(I) transport system, from hCtr1 to the ATPases, Cu(I) movements involve directly coordinate transfers between donor and recipient molecules without metal dissociation. There is only a very small fraction that is loosely bound basal Cu pool or exchangeable Cu pool, which can be monitored using fluorescent sensor, Copper Fluor-3 imaging [54]. It is this labile Cu pool, but not the total cellular Cu content, that regulates hCtr1 expression. Regulation of Ctr1 expression by Cu bioavailability is an evolutionarily conserved mechanism from yeast [55] to humans [56].
Early work in the yeast S. cerevisiae demonstrated that Cu treatment induced yCtr1 internalization, which rapidly degraded [57]. Cu-induced hCtr1 internalization has also been reported in human cells [54][55].
5. Overcoming cDDP Resistance by Copper Chelators
Many human cancers have increased Cu contents that are implicated in tumor angiogenesis, proliferation, and migration. Likewise, Sp1 expression is also frequently elevated in many human cancers [58]. Many preclinical studies have demonstrated that Cu chelators suppress tumor growth in cultured cell models [59] and in animal tumor models (see review [60] and references therein). Cu chelators used in these studies include tetrathiomolybdate (TM) [60], D-penicillamine (D-Pen) [60], trientine triethylenetetraminee dihydrochloride [60], disulfram (DSF) [61][62], and elesclomol (STA-4783) [63] (see reference [60] for the chemical structures and their clinical trials for some of these Cu chelators). TM, D-pen, and trientine are traditional medicines used in treating Wilson’s disease, whereas DSF is a conventional anti-alcoholism drug, and STA-4783 is an anti-neurodegenerative agent recently found to have activity of escorting Cu to the brain in mouse Menke’s disease model [64].
The Cu-Sp1-hCtr1 cycle underscores that the cellular levels of Cu(I), Sp1, and hCtr1 are mutually regulated. The capacities of cellular hCtr1 levels that can be regulated by Cu(I) depletion are constrained and may vary among cell types. For example, cells with reduced basal hCtr1 levels have higher magnitudes of hCtr1 upregulation by Cu chelators than those with elevated basal hCtr1 levels. Because most cDDP-resistant variants are associated with reduced hCtr1 expression [65], these results suggest higher magnitude of hCtr1 induction by chelators in cDDP-resistant cells than in their drug-sensitive counterparts. This predication was confirmed in our study using three cultured cell models and three different Cu-lowering agents (trientine, D-pen, and TM); cDDP-resistant cancer cells exhibit a greater magnitude of hCtr1 upregulation by the Cu-lowering agents as compared with their drug-sensitive counterparts. These observations indicate that reversal of cDDP resistance is independent of Cu-lowering agents and of cell line-specificities [66]. Another study using high-throughput screening identified that disulfiram exhibits synergistic effects with cDDP in bladder cancer cells [67]. Moreover, other naturally occurring Cu-lowering products include carnosine dipeptide (alanyl-L-histidine), which has been reported to modulate the Sp1-hCtr1-Cu homeostasis system [68], and curcumin (a product of plant Curcuma longa used in food flavoring), which enhances the binding of Sp1 to Ctr1 and Sp1 promoters, and thus induces Ctr1 expression and chemosensitization to cDDP treatment [69]
Based on these preclinical observations, two clinical trials have been carried out in testing the efficacies of Cu-lowering agents as enhancers in Pt drug cancer chemotherapy: one (at MD Anderson Cancer Center) involved carboplatin plus trientine in 55 patients with advanced malignancies, 45 of which had prior failure in Pt drug treatment. The results showed that about 19% of patients (n = 9) who maintained low serum Cu levels after the treatments had significantly longer median PFS (p = 0.001) and OS (p = 0.03), as compared with those patients (n = 38) who did not [70][71]. The other study (at the National Cheng Kung University, Taiwan) involved carboplatin plus trientine and pegylated lyposomal doxorubicin in 18 Asian epithelial ovarian tubal and peritoneal cancers. The clinical benefit rate was 33.3 and 50.0% in the Pt-resistant and the partially Pt-sensitive group, respectively [72].
References
- Rottenberg, S.; Disler, C.; Perego, B. The rediscovery of platinum-based cancer therapy. Nat. Rev. Cancer 2021, 21, 37–50.
- Muggia, F.M.; Bonetti, A.; Hoeschele, J.D.; Rozencweig, M.; Howell, S.B. Platinum antitumor complexes: 50 years since Barnett Rosenberg′s discovery. J. Clin. Oncol. 2015, 33, 4219–4226.
- Chiruvella, V.; Annamaraju, P.; Guddati, P.A.K. Management of nephrotoxicity of chemotherapy and targeted agents: 2020. Am. J. Cancer. Res. 2020, 10, 4151–4164.
- Wertman, J.N.; Melong, N.; Stoyek, M.R.; Piccolo, O.; Langley, S.; Orr, B.; Steele, S.L.; Razaghi, B.; Berman, J.N. The identification of dual protective agents against cisplatin-induced oto- and nephrotoxicity using the zebrafish model. eLife 9 2020, 9.
- Rabik, C.A.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat. Rev. 2007, 33, 9–23.
- Wang, D.; Lippard, S.J. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov. 2005, 4, 307–320.
- Kim, E.S.; Lee, J.J.; He, G.; Chow, C.W.; Fujimoto, J.; Kalhor, N.; Swisher, S.G.; Wistuba, I.I.; Stewart, D.J.; Siddik, Z.H. Tissue platinum concentration and tumor response in non-small-cell lung cancer. J. Clin. Oncol. 2012, 30, 3345–3352.
- Gately, D.P.; Sharma, A.; Christen, R.D. Howell, S.B. Cisplatin and taxol activate different signal pathways regulating cellular injury-induced expression of GADD153. Br. J. Cancer 1996, 73, 18–23.
- Planells-Cases, R.; Lutter, D.; Guyader, C.; Gerhards, N.M.; Ullrich, F.; Elger, D.A.; Kucukosmanoglu, A.; Xu, G.; Voss, F.K.; Reincke, S.M.; et al. Subunit composition of VRAC channels determines substrate specificity and cellular resistance to Pt-based anti-cancer drugs. EMBO J. 2015, 34, 2993–3008.
- Konig, B.; Stauber, T. Biophysics and structure-function relationships of LRRC8-formed volume-regulated anion channels. Biophys. J. 2019, 116, 1185–1193.
- Sorensen, B.H.; Dam, C.S.; Sturup, S.; Lambert, I.H. Dual role of LRRC8A-containing transporters on cisplatin resistance in human ovarian cancer cells. J. Inorg. Biochem. 2016, 160, 287–295.
- Ruprecht, N.; Hofmann, L.; Hungerbuhler, M.N.; Kempf, C.; Heverhagen, J.T.; von Tengg-Kobligk, H. Generation of stable cisPt resistant lung adenocarcinoma cells. Pharmaceuticals 2020, 13, 109.
- Sprowl, J.A.; Ciarimboli, G.; Lancaster, C.S.; Giovinazzo, H.; Gibson, A.A.; Du, G.; Janke, L.J.; Cavaletti, G.; Shields, A.F.; Sparreboom, A. Oxaliplatin-induced neurotoxicity is dependent on the organic cation transporter OCT2. Proc. Natl. Acad. Sci. USA 2013, 110, 11199–11204.
- Gu, J.; Wang, L.; Li, T.; Tang, S.; Wang, Y.; Zhang, W.; Jiang, X. Role and mechanism of organic cation transporter 3 in oxaliplatin treatment of colon cancer in vitro and in vivo. Oncol. Rep. 2019, 42, 1355–1364.
- Jong, N.N.; Nakanishi, T.; Liu, J.J.; Tamai, I.; McKeage, M.J. Oxaliplatin transport mediated by organic cation/carnitine transporters OCTN1 and OCTN2 in overexpressing human embryonic kidney 293 cells and rat dorsal root ganglion neurons. J. Pharmacol. Exp. Ther. 2011, 338, 537–547.
- Kato, M.; Tsurudome, Y.; Kanemitsu, T.; Yasukochi, S.; Kanado, Y.; Ogino, T.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Diurnal expression of MRP4 in bone marrow cells underlies the dosing-time dependent changes in the oxaliplatin-induced myelotoxicity. Sci. Rep. 2020, 10, 13484.
- Samodelov, S.L.; Kullak-Ublick, G.A.; Gai, Z.; Visentin, M. Organic cation transporters in human physiology, pharmacology, and toxicology. Int. J. Mol. Sci. 2020, 21, 7890.
- Tatsumi, S.; Matsuoka, H.; Hashimoto, Y.; Hatta, K.; Maeda, K.; Kamoshida, S. Organic cation transporter 2 and tumor budding as independent prognostic factors in metastatic colorectal cancer patients treated with oxaliplatin-based chemotherapy. Int. J. Clin. Exp. Pathol. 2014, 7, 204–212.
- Ishida, S.; Lee, J.; Thiele, D.J.; Herskowitz, I. Uptake of the anticancer drug cisplatin mediated by the copper transporter Ctr1 in yeast and mammals. Proc. Natl. Acad. Sci. USA 2002, 99, 14298–14302.
- Georgatsou, E.; Mavrogiannis, L.A.; Fragiadakis, G.S.; Alexandraki, V. The yeast Fre1p/Fre2p cupric reductases facilitate copper uptake and are regulated by the copper-modulated Mac1p activator. J. Biol. Chem. 1997, 272, 13786–13792.
- Schwab, S.; Shearer, J.; Conklin, S.E.; Alies, B.; Haas, K.L. Sequence proximity between Cu(II) and Cu(I) binding sites of human copper transporter 1 model peptides defines reactivity with ascorbate and O2. J. Inorg. Biochem. 2016, 158, 70–76.
- Puig, S.; Lee, J.; Lau, M.; Thiele, D.J. Biochemical and genetic analyses of yeast and human high affinity copper transporters suggest a conserved mechanism for copper uptake. J. Biol. Chem. 2002, 227, 26021–26030.
- Liang, Z.D.; Stockton, D.; Savaraj, N.; Tien Kuo, M. Mechanistic comparison of human high-affinity copper transporter 1-mediated transport between copper ion and cisplatin. Mol. Pharmacol. 2009, 76, 843–853.
- Selvaraj, A.; Balamurugan, K.; Yepiskoposyan, H.; Zhou, H.; Egli, D.; Georgiev, O.; Thiele, D.J.; Schaffner, W. Metal-responsive transcription factor (MTF-1) handles both extremes, copper load and copper starvation, by activating different genes. Genes Dev. 2005, 19, 891–896.
- Ren, F.; Logeman, B.L.; Zhang, X.; Liu, Y.; Thiele, D.J.; Yuan, P. X-ray structures of the high-affinity copper transporter Ctr1. Nat. Commun. 2019, 10, 1386.
- Barresi, V.; Trovato-Salinaro, A.; Spampinato, G.; Musso, N.; Castorina, S.; Rizzarelli, E.; Condorelli, D.F. Transcriptome analysis of copper homeostasis genes reveals coordinated upregulation of SLC31A1, SCO1, and COX11 in colorectal cancer. FEBS Open Bio 2016, 6, 794–806.
- Perkal, O.; Qasem, Z.; Turgeman, M.; Schwartz, R.; Gevorkyan-Airapetov, L.; Pavlin, M.; Magistrato, A.; Major, D.T.; Ruthstein, S. Cu(I) Controls Conformational States in Human ATOX1 Metallochaperone: An EPR and Multiscale Simulation Study. J. Phys. Chem. B 2020, 124, 4399–4411.
- Wu, X.; Yuan, S.; Wang, E.; Tong, Y.; Ma, G.; Wei, K.; Liu, Y. Platinum transfer from hCTR1 to Atox1 is dependent on the type of platinum complex. Metallomics 2017, 9, 546–555.
- Kahra, D.; Kovermann, M.; Wittung-Stafshede, P. The C-terminus of human copper importer Ctr1 Acts as a binding site and transfers copper to Atox1. Biophys. J. 2016, 110, 95–102.
- Cox, D.W.; Moore, S.D. Copper transporting P-type ATPases and human disease. J. Bioenerg. Biomembr. 2002, 34, 333–338.
- Gudekar, N.; Shanbhag, V.; Wang, Y.; Ralle, M.; Weisman, G.A.; Petris, M.J. Metallothioneins regulate ATP7A trafficking and control cell viability during copper deficiency and excess. Sci. Rep. 2020, 10, 7856.
- Nardella, M.; Rosato, A.; Belviso, B.; Caliardro, R.; Natile, G.; Arnesano, F. Oxidation of human copper chaperone Atoxi and disusulfide bond cleavage by cisplatin and glutathione. Int. J. Mol. Sci. 2019, 20, 4390.
- Tapia, L.; Gonzalez-Aguero, M.; Cisternas, M.F.; Suazo, M.; Cambiazo, V.; Uauy, R.; Gonzalez, M. Metallothionein is crucial for safe intracellular copper storage and cell survival at normal and supra-physiological exposure levels. Biochem. J. 2004, 378, 617–624.
- Kozlowski, H.; Kolkowska, P.; Watly, J.; Krzywoszynska, K.; Potocki, S. General aspects of metal toxicity. Curr. Med. Chem. 2014, 21, 3721–3740.
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—an evolutionary overview. FEBS J. 2008, 275, 3959–3970.
- Ishida, S.; McCormick, F.; Smith-McCune, K.; Hanahan, D. Enhancing tumor-specific uptake of the anticancer drug cisplatin with a copper chelator. Cancer Cell 2010, 17, 574–583.
- Sun, S.; Cai, J.; Yang, Q.; Zhao, S.; Wang, Z. The association between copper transporters and the prognosis of cancer patients undergoing chemotherapy: A meta-analysis of literatures and datasets. Oncotarget 2017, 8, 16036–16051.
- Hanna, N.H.; Einhorn, L.H. Testicular cancer--discoveries and updates. N. Engl. J. Med. 2014, 371, 2005–2016.
- Ghaffari, R.; Di Bona, K.R.; Riley, C.L.; Richburg, J.H. Copper transporter 1 (CTR1) expression by mouse testicular germ cells, but not Sertoli cells, is essential for functional spermatogenesis. PLoS ONE 2019, 14, e0215522.
- Ghaffari, R.; Richburg, J.H. Mice with a Sertoli cell-specific knockout of the Ctr1 gene exhibit a reduced sensitivity to cisplatin-induced testicular germ cell apoptosis. Toxicol. Res. 2019, 8, 972–978.
- Hua, H.; Gunther, V.; Georgiev, O.; Schaffner, W. Distorted copper homeostasis with decreased sensitivity to cisplatin upon chaperone Atox1 deletion in Drosophila. Biometals 2011, 24, 445–453.
- Safaei, R.; Maktabi, M.H.; Blair, B.G.; Larson, C.A.; Howell, S.B. Effects of the loss of Atox1 on the cellular pharmacology of cisplatin. J. Inorg. Biochem. 2009, 103, 333–341.
- Petruzzelli, R.; Polishchuk, R.S. Activity and trafficking of copper-transporting ATPases in tumor development and defense against platinum-based drugs. Cells 2019, 8, 1080.
- Komatsu, M.; Sumizawa, T.; Mutoh, M.; Chen, Z.S.; Terada, K.; Furukawa, T.; Yang, X.L.; Gao, H.; Miura, N.; Sugiyama, T.; et al. Copper-transporting P-type adenosine triphosphatase (ATP7B) is associated with cisplatin resistance. Cancer Res. 2000, 60, 1312–1316.
- Leonhardt, K.; Gebhardt, R.; Mossner, J.; Lutsenko, S.; Huster, D. Functional interactions of Cu-ATPase ATP7B with cisplatin and the role of ATP7B in the resistance of cells to the drug. J. Biol. Chem. 2009, 284, 7793–7802.
- Nakagawa, T.; Abe, M.; Yamazaki, T.; Miyashita, H.; Niwa, H.; Kokubun, S.; Sato, Y. HEX acts as a negative regulator of angiogenesis by modulating the expression of angiogenesis-related gene in endothelial cells in vitro. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 231–237.
- Samimi, G.; Safaei, R.; Katano, K.; Holzer, A.K.; Rochdi, M.; Tomioka, M.; Goodman, M.; Howell, S.B. Increased expression of the copper efflux transporter ATP7A mediates resistance to cisplatin, carboplatin, and oxaliplatin in ovarian cancer cells. Clin. Cancer Res. 2004, 10, 4661–4669.
- Martinez-Balibrea, E.; Martinez-Cardus, A.; Musulen, E.; Gines, A.; Manzano, J.L.; Aranda, E.; Plasencia, C.; Neamati, N.; Abad, A. Increased levels of copper efflux transporter ATP7B are associated with poor outcome in colorectal cancer patients receiving oxaliplatin-based chemotherapy. Int. J. Cancer 2009, 124, 2905–2910.
- Nakayama, K.; Kanzaki, A.; Terada, K.; Mutoh, M.; Ogawa, K.; Sugiyama, T.; Takenoshita, S.; Itoh, K.; Yaegashi, N.; Miyazaki, K.; et al. Prognostic value of the Cu-transporting ATPase in ovarian carcinoma patients receiving cisplatin-based chemotherapy. Clin. Cancer Res. 2004, 10, 2804–2811.
- Weiskirchen, R.; Penning, L.C. COMMD1, a multi-potent intracellular protein involved in copper homeostasis, protein trafficking, inflammation, and cancer. J. Trace Elem. Med. Biol. 2021, 65, 126712.
- Fedoseienko, A.; Wieringa, H.W.; Wisman, G.B.; Duiker, E.; Reyners, A.K.; Hofker, M.H.; van der Zee, A.G.; van de Sluis, B.; van Vugt, M.A. Nuclear COMMD1 is associated with cisplatin sensitivity in ovarian cancer. PLoS ONE 2016, 11, e0165385.
- Eid, C.; Hemadi, M.; Ha-Duong, N.T.; El Hage Chahine, J.M. Iron uptake and transfer from ceruloplasmin to transferrin. Biochim. Biophys. Acta 2014, 1840, 1771–1781.
- Herman, S.; Lipinski, P.; Ogorek, M.; Starzynski, R.; Grzmil, P.; Bednarz, A.; Lenartowicz, M. Molecular regulation of copper homeostasis in the male gonad during the process of spermatogenesis. Int. J. Mol. Sci. 2020, 21, 9053.
- Dodani, S.C.; Firl, A.; Chan, J.; Nam, C.I.; Aron, A.T.; Onak, C.S.; Ramos-Torres, K.M.; Paek, J.; Webster, C.M.; Feller, M.B.; et al. Copper is an endogenous modulator of neural circuit spontaneous activity. Proc. Natl. Acad. Sci. USA 2014, 111, 16280–16285.
- Shi, H.; Jiang, Y.; Yang, Y.; Peng, Y.; Li, C. Copper metabolism in Saccharomyces cerevisiae: An update. Biometals 2020, 34, 3–14.
- Lai, Y.H.; Kuo, C.; Kuo, M.T.; Chen, H.H.W. Modulating chemosensitivity of tumors to platinum-based antitumor drugs by transcriptional regulation of copper homeostasis. Int. J. Mol. Sci. 2018, 19, 1486.
- Ooi, C.E.; Rabinovich, E.; Dancis, A.; Bonifacino, J.S.; Klausner, R.D. Copper-dependent degradation of the Saccharomyces cerevisiae plasma membrane copper transporter Ctr1p in the apparent absence of endocytosis. EMBO J. 1996, 15, 3515–3523.
- Safe, S.; Abbruzzese, J.; Abdelrahim, M.; Hedrick, E. Specificity protein transcription factors and cancer: Opportunities for drug development. Cancer Prev. Res. 2018, 11, 371–382.
- Helsel, M.E.; Franz, K.J. Pharmacological activity of metal binding agents that alter copper bioavailability. Dalton Trans. 2015, 44, 8760–8770.
- Baldari, S.; Di Rocco, G.; Toietta, G. Current biomedical use of copper chelation therapy. Int. J. Mol. Sci. 2020, 21, 1069.
- Cen, D.; Gonzalez, R.I.; Buckmeier, J.A.; Kahlon, R.S.; Tohidian, N.B.; Meyskens, F.L., Jr. Disulfiram induces apoptosis in human melanoma cells: A redox-related process. Mol. Cancer Ther. 2002, 1, 197–204.
- Chen, D.; Cui, Q.C.; Yang, H.; Dou, Q.P. Disulfiram, a clinically used anti-alcoholism drug and copper-binding agent, induces apoptotic cell death in breast cancer cultures and xenografts via inhibition of the proteasome activity. Cancer Res. 2006, 66, 10425–10433.
- Hasinoff, B.B.; Wu, X.; Yadav, A.A.; Patel, D.; Zhang, H.; Wang, D.S.; Chen, Z.S.; Yalowich, J.C. Cellular mechanisms of the cytotoxicity of the anticancer drug elesclomol and its complex with Cu(II). Biochem. Pharmacol. 2015, 93, 266–276.
- Guthrie, L.M.; Soma, S.; Yuan, S.; Silva, A.; Zulkifli, M.; Snavely, T.C.; Greene, H.F.; Nunez, E.; Lynch, B.; De Ville, C.; et al. Elesclomol alleviates Menkes pathology and mortality by escorting Cu to cuproenzymes in mice. Science 2020, 368, 620–625.
- Long, Y.; Tsai, W.B.; Chang, J.T.; Estecio, M.; Wangpaichitr, M.; Savaraj, N.; Feun, L.G.; Chen, H.H.; Kuo, M.T. Cisplatin-induced synthetic lethality to arginine-starvation therapy by transcriptional suppression of ASS1 is regulated by DEC1, HIF-1alpha, and c-Myc transcription network and is independent of ASS1 promoter DNA methylation. Oncotarget 2016, 7, 82658–82670.
- Liang, Z.D.; Long, Y.; Tsai, W.B.; Fu, S.; Kurzrock, R.; Gagea-Iurascu, M.; Zhang, F.; Chen, H.H.; Hennessy, B.T.; Mills, G.B.; et al. Mechanistic basis for overcoming platinum resistance using copper chelating agents. Mol. Cancer Ther. 2012, 11, 2483–2494.
- Kita, Y.; Hamada, A.; Saito, R.; Teramoto, Y.; Tanaka, R.; Takano, K.; Nakayama, K.; Murakami, K.; Matsumoto, K.; Akamatsu, S.; et al. Systematic chemical screening identifies disulfiram as a repurposed drug that enhances sensitivity to cisplatin in bladder cancer: A summary of preclinical studies. Br. J. Cancer 2019, 121, 1027–1038.
- Barca, A.; Ippati, S.; Urso, E.; Vetrugno, C.; Storelli, C.; Maffia, M.; Romano, A.; Verri, T. Carnosine modulates the Sp1-Slc31a1/Ctr1 copper-sensing system and influences copper homeostasis in murine CNS-derived cells. Am. J. Physiol. Cell Physiol. 2019, 316, C235–C245.
- Zhang, W.; Shi, H.; Chen, C.; Ren, K.; Xu, Y.; Liu, X.; He, L. Curcumin enhances cisplatin sensitivity of human NSCLC cell lines through influencing Cu-Sp1-CTR1 regulatory loop. Phytomedicine 2018, 48, 51–61.
- Fu, S.; Hou, M.M.; Wheler, J.; Hong, D.; Naing, A.; Tsimberidou, A.; Janku, F.; Zinner, R.; Piha-Paul, S.; Falchook, G.; et al. Exploratory study of carboplatin plus the copper-lowering agent trientine in patients with advanced malignancies. Investig. New Drugs 2014, 32, 465–472.
- Fu, S.; Naing, A.; Fu, C.; Kuo, M.T.; Kurzrock, R. Overcoming platinum resistance through the use of a copper-lowering agent. Mol. Cancer Ther. 2012, 11, 1221–1225.
- Huang, Y.F.; Kuo, M.T.; Liu, Y.S.; Cheng, Y.M.; Wu, P.Y.; Chou, C.Y. A dose escalation study of trientine plus carboplatin and pegylated liposomal doxorubicin in women with a first relapse of epithelial ovarian, tubal, and peritoneal cancer within 12 months after platinum-based chemotherapy. Front. Oncol. 2019, 9, 437.
- Leitao, M.M., Jr.; Hummer, A.; Dizon, D.S.; Aghajanian, C.; Hensley, M.; Sabbatini, P.; Venkatraman, E.; Spriggs, D.R. Platinum retreatment of platinum-resistant ovarian cancer after nonplatinum therapy. Gynecol. Oncol. 2003, 91, 123–129.
- Kavanagh, J.; Tresukosol, D.; Edwards, C.; Freedman, R.; Gonzalez de Leon, C.; Fishman, A.; Mante, R.; Hord, M.; Kudelka, A. Carboplatin reinduction after taxane in patients with platinum-refractory epithelial ovarian cancer. J. Clin. Oncol. 1995, 13, 1584–1588.
More
Information
Subjects:
Pharmacology & Pharmacy
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.2K
Revision:
1 time
(View History)
Update Date:
21 Jun 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No