 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Swarup Roy Choudhury | + 4785 word(s) | 4785 | 2020-06-08 08:59:05 | | | |
| 2 | Rita Xu | -1985 word(s) | 2800 | 2020-06-19 09:47:19 | | | | |
| 3 | Rita Xu | -13 word(s) | 2787 | 2020-10-27 04:49:59 | | |
Video Upload Options
Receptors form the crux for any biochemical signaling. Receptor-like kinases (RLKs) are conserved protein kinases in eukaryotes that establish signaling circuits to transduce information from outer plant cell membrane to the nucleus of plant cells, eventually activating processes directing growth, development, stress responses, and disease resistance. Plant RLKs share considerable homology with the receptor tyrosine kinases (RTKs) of the animal system, differing at the site of phosphorylation. Typically, RLKs have a membrane-localization signal in the amino-terminal, followed by an extracellular ligand-binding domain, a solitary membrane-spanning domain, and a cytoplasmic kinase domain. The functional characterization of ligand-binding domains of the various RLKs has demonstrated their essential role in the perception of extracellular stimuli, while its cytosolic kinase domain is usually confined to the phosphorylation of their substrates to control downstream regulatory machinery. Identification of the several ligands of RLKs, as well as a few of its immediate substrates have predominantly contributed to a better understanding of the fundamental signaling mechanisms. In the model plant Arabidopsis, several studies have indicated that multiple RLKs are involved in modulating various types of physiological roles via diverse signaling routes. Here, we summarize recent advances and provide an updated overview of transmembrane RLKs in Arabidopsis.
1. Introduction
Arabidopsis RLKs modulate growth and developmental responses by governing stem-cell maintenance, cell fate determination and patterning, male and female gametophyte development, pollen-pistil interactions, embryogenesis, hormone signaling, vascular patterning, organ development, and abscission. Some of these essential responses are discussed here.
2. Regulation in Anther and Ovule Development
The anther generally has four lobes and each lobe contains reproductive microsporocyte surrounded by various layers of somatic cells viz., tapetum, middle layer, endothecium, and epidermis. In Arabidopsis, multiple LRR-RLKs like excess microsporocytes1 (EMS1)/extra sporogenous cell (EXS), somatic embryogenesis receptor-like kinase 1/2 (SERK1/2), receptor-like protein kinase 2 (RPK2), barely any meristem 1/2 (BAM1/2), CLAVATA3 insensitive receptor kinase (CIK1/2/3/4), ERECTA (ER), and ERECTA-like 1/2 (ERL1/2) regulate anther development, especially, the differentiation and patterning of the somatic cell layers. EMS1/EXS was the first LRR-RLK to be identified that plays a crucial role in anther cell differentiation [1][2]. The anthers of ems1/exs mutants lack tapetum but produce large numbers of microsporocytes than the wild type. In addition, delayed expression of EMS1 in the ems1 mutant tapetal initials has been shown to aid in the generation of a functional tapetum and the diminution of microsporocyte numbers [3]. These results suggest that EMS1/EXS determines the fate of tapetal cells during early anther development. Tapetum determinant 1 (TPD1), a small secreted protein, is known to induce the phosphorylation of EMS1/EXS, thus, behaving as their ligand; and the signal is transduced downstream via phosphorylation of β-carbonic anhydrases (βCAs) [4][5]. Similarly, SERK1/2 has also been known to determine tapetal cell fate, as the anthers of serk1serk2 double mutants are phenotypically similar to that of ems1/exs mutant [6][7]. Moreover, SERK1 interacts with and transphosphorylates EMS1 to enhance its activity for guiding a co-regulatory network (Figure 1A) [8]. Corroborated by the phenotype of rpk2 mutants, it can be deduced that RPK2 is responsible for the differentiation of middle layers and tapetum during anther development. It essentially controls tapetal cell fate by triggering their degradation via modulation of the enzymes involved in cell wall metabolism and lignin biosynthesis [9] (Figure 1A). Both BAM1 and BAM2 are responsible for regulating early stages of anther differentiation, as confirmed by the lack of somatic cell layers, including endothecium, middle layer, and tapetum in bam1bam2 double mutants [10]. CLAVATA3 insensitive receptor kinases (CIK1/2/3/4) are co-receptors of BAM1/2 and RPK2, which regulate the determination of parietal cell fate and archesporial cell division [11] (Figure 1A). ERECTA (ER), ERECTA-Like 1 (ERL1), and ERL2 are also known to play essential roles in healthy anther lobe formation and anther cell differentiation via mitogen-activated protein kinases like MPK3/MPK6 (Figure 1A). The sterility of er-105 erl1-2 erl2-1 triple mutant and the phenotypic similarity of the anther lobes in single mutants of er-105 or erl1-2 or erl2-1 with that of mpk3 or mpk6 mutants suggests the correlation of these genes in the regulation of anther cell division and differentiation [12]. Further, a Lectin RLK, small, glued together, collapsed (SGC) has also been validated as a regulator of pollen development as its knockout had led to the development of small, glued-together and collapsed pollen and resulted in male sterility [13] (Figure 1A).
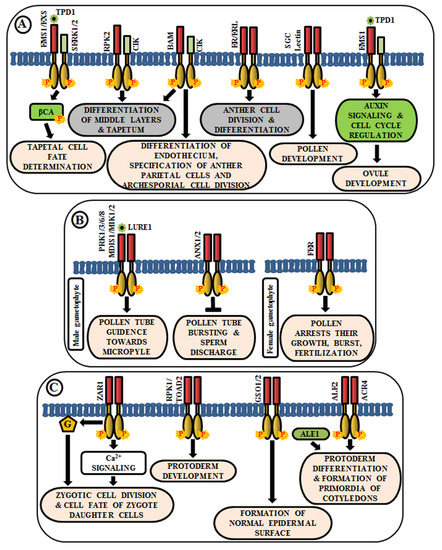
Figure 1. Arabidopsis RLKs in the regulation of growth and development. A few examples of RLKs that regulate (A) anther and ovule development, (B) pollen-pistil interaction, and (C) embryo development.
Knowledge about the role of RLKs in ovule development is very scarce. In Arabidopsis ovules, EMS1 is expressed in nucellar epidermis and chalaza, while TPD1 is weakly restricted to the distal end of integuments. Altered expression of cell-cycle genes and auxin signaling genes during ovule development, concomitant with the ectopic expression of TPD1, indicates the regulation of ovule development by TPD1-EMS1 [14] (Figure 1A).
3. Pollen-Pistil Interactions
Reproduction in angiosperms involves the release of an immobile male gamete from the pollen tube onto the compatible pistil. A fruitful pollen-pistil interaction is a pre-requisite for successful fertilization and this requires an accurate perception of ovule-emitted guidance cues by the receptors in pollen tubes. LURE1, an ovule-secreted peptide is perceived by RLKs like pollen receptor kinase 1 (PRK1), PRK3, PRK6, PRK8 in the pollen tube [15]. Recent studies ascertain the presence of other LURE receptors like Male Discoverer 1 (MDIS1), MDIS1-interacting receptor-like kinase1 (MIK1), and MIK2 [16][17]. Once the pollen tube reaches the micropyle, its growth is ceased and the sperm cells are released by its rupture. These processes are regulated by the RLK FERONIA (FER), which is expressed in the synergids of female gametophyte [18] (Figure 1B). The phenotypic study of fer mutants exhibited overgrowth of pollen tube and loss of its rupturing ability [19]. ANXUR1 and ANXUR2 (ANX1, ANX2) are homologs of FER-RLK, expressed at the tip of the pollen tube. The anx1anx2 double mutants have been found to arrest the growth of pollen tubes and promote bursting immediately after germination. These validate the clue that both FER-mediated and ANX-dependent signaling cascades act as a switch for accurate pollen tube growth and subsequent release of sperm cells for fertilization [20] (Figure 1B).
4. Role in Embryo Development
After successful fertilization, the zygote develops into embryo via repeated cell division and differentiation. Several genetic evidences suggest that multiple signaling cascades are essential for embryogenesis in Arabidopsis, and RLK is one amongst them. Predominantly, embryo development initiates from the asymmetric division of the zygote. Intriguingly, the transcript of ZYGOTIC ARREST 1 (ZAR1), a LRR-RLK, has been detected in the embryo sac before and after fertilization. It has been noticed in an eight-nucleate stage of embryo sac to different cells of mature embryo sac including the central cell, egg cell, and synergids. Even after fertilization, it was observed in the endosperm. Phenotypic analysis of zar1 mutants revealed the role of ZAR1 in the regulation of asymmetric division of zygote and determination of the cell fate of its daughter cells via the activation of calcium and G-protein signaling cascades [21] (Figure 1C). Besides ZAR1, receptor-like protein kinase 1 (RPK1) and Toadstool 2 (TOAD2) are considered indispensable for normal protoderm development, while GASSHO 1 (GSO1) and GSO2 are crucial for the formation of the proper epidermal surface during embryogenesis. The gso1gso2 double mutants have shown abnormal bending of embryos, highly permeable epidermal structure, and irregular stomatal patterning [22][23] (Figure 1C). Further, molecular analysis has detected the interaction of ALE2 (Abnormal Leaf Shape 2) and ACR4 (CRINKLY 4) with a subtilisin-like serine protease ALE1, which is essential for the formation of primordia of cotyledons during embryogenesis [24] (Figure 1C).
5. Organ Development
Coordinated cell growth, differentiationand morphogenesis are the three fundamental aspects of development that cause an organism to procure its shape and an intricate cascade of gene regulatory networks comprising RLKs are known to be implicated in this. In higher plants, all the aerial organs develop from shoot apical meristem (SAM). The maintenance of undifferentiated cells of SAM and organ formation through differentiation from the progeny cells are two processes maintained in a balance during the common developmental process. Interestingly, different RLKs are known to suffice this role. In Arabidopsis, CLAVATA1 or CLV1 (RLK), CLV2 (RLP) and CLV3 (secreted polypeptide) perform a pivotal role in meristem and organ development [25][26][27]. The CLV3 polypeptide acts as a ligand for CLV1 and CLV2 complex. This ligand-receptor binding promotes the activation of cytosolic kinase domain of CLV1 and subsequently, it initiates a signal transduction cascade to control gene expression and stem cell fate in the SAM by elevation of cytosolic calcium as secondary messengers [25][28][29] (Figure 2A). Meristematic receptor-like kinase (MRLK), a LRR-RLK expressed in shoot and root apical meristems, interacts with and phosphorylates a MADS-box transcription factor, AGL24, to regulate floral transition [30] (Figure 2A). Another LRR-RLK, ERECTA, which is expressed in the entire shoot apical meristem and developing organs, monitors organ shape and inflorescence architecture, upon the perception of epidermal patterning factors (EPFs)/EPF-like proteins (EPFLs) [31] (Figure 2A). Moreover, mutants of ERECTA-family LRR-RLKs conferred extreme dwarfism and abnormal flower development, suggesting that ERECTA-family RLKs control cell proliferation as well as organ growth and patterning like stomata formation, the shoot apical meristem (SAM) and flower development [32]. ERECTA can form complexes with a range of co-receptors like SERKs and transmembrane receptor-like proteins like Too Many Mouths (TMM) to activate the signaling pathway [33][34]. Botrytis-induced kinase 1 or BIK1, an RLCK, interacts and phosphorylates ER-family proteins to modulate leaf morphogenesis and inflorescence architecture [35] (Figure 2A).
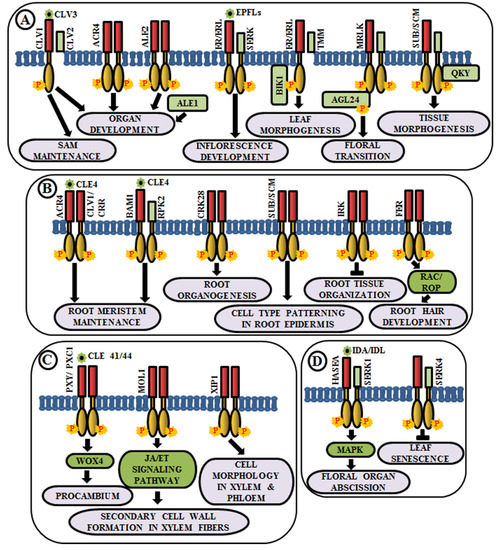
Figure 2. Arabidopsis RLKs in the regulation of growth and development. A few examples of RLKs that regulate (A) shoot development, (B) root development, (C) vascular tissue development, and (D) organ abscission.
Similar to aboveground organ development, several studies demonstrated the utmost importance of multiple RLKs in root development. Arabidopsis CRINKLY 4 (ACR4) is involved in the formation of proper lateral roots and columella stem cell differentiation in the root apical meristem [36][37]. ACR4 can regulate root meristem maintenance in response to the CLE4 peptide by forming heterodimers with CLV1 [38] (Figure 2B). Besides, ACR4, abnormal leaf shape 1 (ALE1) (a member of subtilisin-like serine protease family), and ALE2 (RLK) have been known to share partial overlapping roles in the formation of leafy organs [24] (Figure 2A). Similar to ACR4, cysteine-rich receptor-like kinases (CRKs), a member of one of the largest RLK families, is involved in root organogenesis. The crk28 mutants have displayed longer and branched roots, while CRK28 overexpression lines have shown the contrasting phenotype, i.e., delayed root growth and reduced lateral root formation [39] (Figure 2B).
Plasmodesmata are microchannels between two cells, through which trafficking of molecules occur. STRUBBELIG (SUB) is a RLK involved in inter-cell layer signaling which is required for tissue morphogenesis. The sub mutants have shown defects in floral organ shape, integument initiation, and outgrowth, asymmetry in leaf shape and stem morphology, as well as a reduction in plant height. This indicates the functional role of SUB across several cells in the floral meristem, ovule, and shoot apex [40][41]. Further genetic screening has led to the identification of a putative membrane-anchored C2-domain protein, encoded by QUIRKY (QKY), which is known to act as a downstream component of SUB signaling [42]. SUB and QKY interact in plasmodesmata to promote tissue morphogenesis (Figure 2A). Apart from aerial organs, SUB or SCRAMBLED (SCM) also regulates cell-type patterning in the root epidermis [43] (Figure 2B). The BAM1 (barely any meristem 1), a member of CLV1 class LRR-RLKs, is expressed preferentially in the quiescent center and its surrounding stem cells at the root tip and known to bind to the CLE peptide. BAM1 is capable of forming heteromeric complexes with RPK2 and inhibit cell proliferation in the root meristem [44] (Figure 2B). Inflorescence and root apices receptor kinase (IRK), a typical meristematic LRR-RLK, is known to be expressed in the outer plasma membrane of root endodermal cells and negatively regulates cell division to maintain tissue organization [45] (Figure 2B). Further, FERONIA (FER) receptor-like kinase functions upstream of Rho-like small G-protein or RAC/ROP during reactive oxygen species (ROS)-mediated root hair development. The FER activates RAC/ROP by GDP-GTP exchange to stimulate NADPH oxidase for ROS formation [46] (Figure 2B).
6. Vascular Tissue Development
The development of xylem and phloem from the vascular meristem is a multifaceted process. The RLK, phloem intercalated with xylem (PXY), maintains cell polarity during vascular development, which is ascertained by the presence of partially interspersed xylem and phloem, and irregular vascular development in pxy mutants [47]. The ligand for PXY receptor is tracheary element differentiation factor (TDIF), a peptide, which is encoded by CLAVATA3/ESR 41/44 (CLE41/44) genes [48]. The PXY-TDIF interaction activates the WUSCHEL-related homeobox 4 (WOX4) signaling pathway to regulate cell division in the procambium. Another LRR-RLK, PXY/TDR-CORRELATED (PXC1), acts as a positive regulator of secondary cell wall formation in xylem fibers [49] (Figure 2C). The CLE41/PXY/WOX4 cascade is antagonistically directed by the LRR-RLK more lateral growth 1 (MOL1), via regulating the stem cell homeostasis within the cambium. This MOL1 also attenuates ethylene and jasmonic acid hormone signaling pathways that positively influence cambium activity [50] (Figure 2C). The maintenance of the cell morphology organization during vascular development is accomplished by a RLK, xylem intermixed with phloem 1 (XIP1). Genetic evidences also unveil that XIP1 prevents ectopic lignification in phloem cells [51] (Figure 2C).
7. Regulation of Organ Abscission
Arabidopsis LRR–RLK HAESA (formerly named RLK5) exhibits developmentally regulated expression in the abscission layers of floral organs. The antisense suppression of the HAESA is known to delay the abscission of floral organs such as sepals, petals, and stamens [52]. Inflorescence deficient in abscission (IDA) and IDA-Like (IDL) proteins are considered as the ligands of HAESA (HAE) and HAESA-Like RLKs [53] (Figure 2D). The phenotypic analysis of ida mutant and overexpression of IDA gene validates the role of HAE in floral organ abscission via IDA/IDL perception. A phosphorylation-based activation mechanism of HAE leads to the stimulation of a MAP kinase-signaling cascade and initiates cell wall hydrolysis at the base of the abscising organs. SERK1 acts as a co-receptor of HAE and allows the binding of IDA, eventually leading to floral abscission pathway [54][55]. In contrast, an early leaf senescence phenotype observed in serk4-1 knockout mutant indicates that SERK4 acts as a co-receptor in negatively regulating leaf senescence, as well [56] (Figure 2D).
8. Modulation of Phytohormone Signaling
Brassinosteroids (BRs) are essential polyhydroxylated steroidal phytohormones crucial for plant development. The developmental defects of BR biosynthetic and signaling mutants are mostly similar, which include dwarfism, severely stunted and rounded leaf with a shorter petiole, delayed flowering, photomorphogenic malfunctions as well as senescence and reduced male fertility. The first BR signaling gene, whose mutation showed these phenotypes, has been named as brassinosteroid insensitive 1 (BRI1) [133]. BAK1 (BRI1-associated receptor kinase 1), a co-receptor of BRI1, is involved in BR perception and signaling via heterodimerization with BRI1 [57][58]. In addition, a close homologue of BRI1, BRI1-like receptor kinase (BRL1) is also responsible for BR perception [59] (Figure 3A). BAK1-associating receptor-like kinase 1 (BARK1), a LRR-RLK, specifically binds to BAK1 and its homologs. Overexpression of BARK1 enhances primary root growth and these roots are hypersensitive to BR-induced root growth inhibition, suggesting the role of BARK1 in BR-mediated lateral root development via auxin signaling [60] (Figure 3A). Apart from these, evidence achieved from bir1 mutants helps us to comprehend how it modulates immune response pathways and plant architecture as an interacting partner of BAK1 [61]. A member of somatic embryogenesis receptor, SERK3 acts as a co-receptor, which directly interacts with BRI1 [62] (Figure 3A).
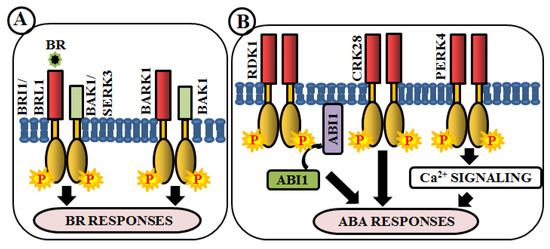
Figure 3. Arabidopsis RLKs in brassinosteroid (BR) and abscisic acid (ABA) signaling. RLK-mediated phosphorylation-based signaling circuits regulate BR (A) and ABA (B) responses.
Abscisic acid (ABA) is yet another vital phytohormone involved in the regulation of plant abiotic stress-related phenotype as well as developmental processes. Unlike BR, in Arabidopsis, RLKs are not accountable for direct ABA perception. A LRR-RLK, receptor dead kinase 1 (RDK1) is involved in ABA signal transduction via interaction with abscisic acid insensitive 1 (ABI1), a type 2C protein phosphatase, in the plasma membrane. Predominantly, this interaction is enhanced by exogenous application of ABA, underpinning the involvement of RDK1 to recruit ABI1 to the plasma membrane [63] (Figure 3B). Most recently, a cysteine-rich receptor-like kinase, CRK28, has shown an indirect relationship with ABA. The CRK28 overexpression lines have displayed slow root growth, reduced lateral root formation, and also ABA hypersensitivity; thereby being an important modulator of ABA signaling [39] (Figure 3B). PERK4 is also known to play an important role in ABA response. The perk4 mutants have shown reduced sensitivity to ABA concerning seed germination, seedling growth, and primary root tip growth. Moreover, perk4 mutant cells have retained lower cytosolic calcium concentration and Ca2+ channel currents. These results suggest that PERK4 contributes to the early stage of ABA signaling and inhibits root cell elongation via intracellular calcium signaling [64] (Figure 3B). Other RLKs like CRK5, CRK36, LRK10L1.2, and RPK1 are also known to be involved in ABA signaling during response to drought and oxidative stresses.
References
- Claudia Canales; Anuj M. Bhatt; Rod Scott; Hugh Dickinson; EXS, a Putative LRR Receptor Kinase, Regulates Male Germline Cell Number and Tapetal Identity and Promotes Seed Development in Arabidopsis. Current Biology 2002, 12, 1718-1727, 10.1016/s0960-9822(02)01151-x.
- Da-Zhong Zhao; Guan-Fang Wang; Brooke Speal; Hong Ma; The EXCESS MICROSPOROCYTES1 gene encodes a putative leucine-rich repeat receptor protein kinase that controls somatic and reproductive cell fates in the Arabidopsis anther. Genes & Development 2002, 16, 2021-2031, 10.1101/gad.997902.
- Xiaoqi Feng; Hugh G. Dickinson; Packaging the male germline in plants. Trends in Genetics 2007, 23, 503-510, 10.1016/j.tig.2007.08.005.
- Gengxiang Jia; Xiaodong Liu; Heather A. Owen; Dazhong Zhao; Signaling of cell fate determination by the TPD1 small protein and EMS1 receptor kinase. Proceedings of the National Academy of Sciences 2008, 105, 2220-2225, 10.1073/pnas.0708795105.
- Jian Huang; Zhiyong Li; Gabriel Biener; Erhui Xiong; Shikha Malik; Nathan Eaton; Catherine Z. Zhao; Valerică Raicu; Hongzhi Kong; Dazhong Zhao; et al. Carbonic Anhydrases Function in Anther Cell Differentiation Downstream of the Receptor-Like Kinase EMS1. The Plant Cell 2017, 29, 1335-1356, 10.1105/tpc.16.00484.
- Catherine Albrecht; Eugenia Russinova; Valérie Hecht; Erik Baaijens; Sacco C. De Vries; The Arabidopsis thaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES1 and 2 Control Male Sporogenesis. The Plant Cell 2005, 17, 3337-3349, 10.1105/tpc.105.036814.
- Jean Colcombet; Aurélien Boisson-Dernier; Roc Ros; Carlos E. Vera; Julian I Schroeder; Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASES1 and 2 Are Essential for Tapetum Development and Microspore Maturation. The Plant Cell 2005, 17, 3350-3361, 10.1105/tpc.105.036731.
- Zhiyong Li; Yao Wang; Jian Huang; Nagib Ahsan; Gabriel Biener; Joel Paprocki; Jay J. Thelen; Valerică Raicu; Dazhong Zhao; Two SERK Receptor-Like Kinases Interact with EMS1 to Control Anther Cell Fate Determination. Plant Physiology 2017, 173, 326-337, 10.1104/pp.16.01219.
- Shinji Mizuno; Yuriko Osakabe; Kyonoshin Maruyama; Takuya Ito; Keishi Osakabe; Takahide Sato; Kazuo Shinozaki; Kazuko Yamaguchi-Shinozaki; Receptor-like protein kinase 2 (RPK 2) is a novel factor controlling anther development in Arabidopsis thaliana. The Plant Journal 2007, 50, 751-766, 10.1111/j.1365-313x.2007.03083.x.
- Carey L.H. Hord; Changbin Chen; Brody J. Deyoung; Steven E. Clark; Hong Ma; The BAM1/BAM2 Receptor-Like Kinases Are Important Regulators of Arabidopsis Early Anther Development[W]. The Plant Cell 2006, 18, 1667-1680, 10.1105/tpc.105.036871.
- Yanwei Cui; Chong Hu; Yafen Zhu; Kaili Cheng; Xiaonan Li; Zhuoyun Wei; Li Xue; Fang Lin; Hongyong Shi; Jing Yi; et al.Suiwen HouKai HeJia LiXiaoping Gou CIK Receptor Kinases Determine Cell Fate Specification during Early Anther Development in Arabidopsis. The Plant Cell 2018, 30, 2383-2401, 10.1105/tpc.17.00586.
- Carey L.H. Hord; Yu-Jin Sun; Lynn J. Pillitteri; Keiko U. Torii; Huachun Wang; Shuqun Zhang; Hong Ma; Regulation of Arabidopsis Early Anther Development by the Mitogen-Activated Protein Kinases, MPK3 and MPK6, and the ERECTA and Related Receptor-Like Kinases. Molecular Plant 2008, 1, 645-658, 10.1093/mp/ssn029.
- Jinrong Wan; Ami Patel; Melanie Mathieu; Sung Yong Kim; Ng Xu; Gary Stacey; A lectin receptor-like kinase is required for pollen development in Arabidopsis. Plant Molecular Biology 2008, 67, 469-482, 10.1007/s11103-008-9332-6.
- Jian Huang; Asela J. Wijeratne; Chong Tang; Tianyu Zhang; Rebecca E. Fenelon; Heather A. Owen; Dazhong Zhao; Ectopic expression of TAPETUM DETERMINANT1 affects ovule development in Arabidopsis. Journal of Experimental Botany 2016, 67, 1311-1326, 10.1093/jxb/erv523.
- Hidenori Takeuchi; Tetsuya Higashiyama; A species-specific cluster of defensin-like genes encodes diffusible pollen tube attractants in Arabidopsis. PLOS Biology 2012, 10, e1001449, 10.1371/journal.pbio.1001449.
- Hidenori Takeuchi; Tetsuya Higashiyama; Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 2016, 531, 245-248, 10.1038/nature17413.
- Tong Wang; Liang Liang; Yong Xue; Peng-Fei Jia; Wei Chen; Meng-Xia Zhang; Ying-Chun Wang; Hong-Ju Li; Wei-Cai Yang; Corrigendum: A receptor heteromer mediates the male perception of female attractants in plants. Nature 2016, 536, 360, 10.1038/nature17985.
- Juan-Miguel Escobar-Restrepo; Norbert Huck; Sharon A. Kessler; Valeria Gagliardini; Jacqueline Gheyselinck; Wei-Cai Yang; Ueli Grossniklaus; The FERONIA Receptor-like Kinase Mediates Male-Female Interactions During Pollen Tube Reception. Science 2007, 317, 656-660, 10.1126/science.1143562.
- Norbert Huck; James M. Moore; Michael Federer; Ueli Grossniklaus; The Arabidopsis mutant feronia disrupts the female gametophytic control of pollen tube reception. Development 2003, 130, 2149-2159, 10.1242/dev.00458.
- Aurélien Boisson-Dernier; Sucharita Roy; Konstantinos Kritsas; Monica A. Grobei; Miloslawa Jaciubek; Julian I Schroeder; Ueli Grossniklaus; Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 2009, 136, 3279-3288, 10.1242/dev.040071.
- Tian-Ying Yu; Dong-Qiao Shi; Peng-Fei Jia; Jun Tang; Hong-Ju Li; Jie Liu; Wei-Cai Yang; The Arabidopsis Receptor Kinase ZAR1 Is Required for Zygote Asymmetric Division and Its Daughter Cell Fate. PLOS Genetics 2016, 12, e1005933, 10.1371/journal.pgen.1005933.
- Michael D. Nodine; Ramin Yadegari; Frans E. Tax; RPK1 and TOAD2 Are Two Receptor-like Kinases Redundantly Required for Arabidopsis Embryonic Pattern Formation. Developmental Cell 2007, 12, 943-956, 10.1016/j.devcel.2007.04.003.
- Ryo Tsuwamoto; Hiroyuki Fukuoka; Yoshihito Takahata; GASSHO1 and GASSHO2 encoding a putative leucine-rich repeat transmembrane-type receptor kinase are essential for the normal development of the epidermal surface in Arabidopsis embryos. The Plant Journal 2008, 54, 30-42, 10.1111/j.1365-313x.2007.03395.x.
- Hirokazu Tanaka; M. Watanabè; M. Sasabe; T. Hiroe; T. Tanaka; Hirokazu Tsukaya; M. Ikezaki; C. Machida; Y. Machida; Novel receptor-like kinase ALE2 controls shoot development by specifying epidermis in Arabidopsis. Development 2007, 134, 1643-1652, 10.1242/dev.003533.
- Sangho Jeong; Amy E. Trotochaud; Steven E. Clark; The Arabidopsis CLAVATA2 Gene Encodes a Receptor-like Protein Required for the Stability of the CLAVATA1 Receptor-like Kinase. The Plant Cell 1999, 11, 1925-1934, 10.1105/tpc.11.10.1925.
- S. E. Clark; Organ Formation at the Vegetative Shoot Meristem. The Plant Cell 1997, 9, 1067-1076, 10.1105/tpc.9.7.1067.
- J. C. Fletcher; Signaling of Cell Fate Decisions by CLAVATA3 in Arabidopsis Shoot Meristems. Science 1999, 283, 1911-1914, 10.1126/science.283.5409.1911.
- Michael Lenhard; Thomas Laux; Stem cell homeostasis in the Arabidopsis shoot meristem is regulated by intercellular movement of CLAVATA3 and its sequestration by CLAVATA1. Development 2003, 130, 3163-3173, 10.1242/dev.00525.
- Hsuan Chou; Yingfang Zhu; Yi Ma; Gerald A. Berkowitz; The CLAVATA signaling pathway mediating stem cell fate in shoot meristems requires Ca2+as a secondary cytosolic messenger. The Plant Journal 2016, 85, 494-506, 10.1111/tpj.13123.
- Hidetomo Fujita; Miho Takemura; Emi Tani; Kyoko Nemoto; Akiho Yokota; Takayuki Kohchi; An Arabidopsis MADS-Box Protein, AGL24, is Specifically Bound to and Phosphorylated by Meristematic Receptor-Like Kinase (MRLK). Plant and Cell Physiology 2003, 44, 735-742, 10.1093/pcp/pcg092.
- Ryusuke Yokoyama; Taku Takahashi; Atsushi Kato; Keiko U. Torii; Y Komeda; The Arabidopsis ERECTA gene is expressed in the shoot apical meristem and organ primordia. The Plant Journal 1998, 15, 301-310, 10.1046/j.1365-313x.1998.00203.x.
- Elena D. Shpak; Michael B. Lakeman; Keiko U. Torii; Dominant-Negative Receptor Uncovers Redundancy in the Arabidopsis ERECTA Leucine-Rich Repeat Receptor–Like Kinase Signaling Pathway That Regulates Organ Shape. The Plant Cell 2003, 15, 1095-1110, 10.1105/tpc.010413.
- Jin Suk Lee; Takeshi Kuroha; Marketa Hnilova; Dmitriy Khatayevich; Masahiro Kanaoka; Jessica M. McAbee; Mehmet Sarikaya; Candan Tamerler; Keiko U. Torii; Direct interaction of ligand-receptor pairs specifying stomatal patterning. Genes & Development 2012, 26, 126-136, 10.1101/gad.179895.111.
- Xiangzong Meng; Xin Chen; Hyunggon Mang; Chenglong Liu; Xiao Yu; Xiquan Gao; Keiko U. Torii; Ping He; Libo Shan; Differential Function of Arabidopsis SERK Family Receptor-like Kinases in Stomatal Patterning.. Current Biology 2015, 25, 2361-2372, 10.1016/j.cub.2015.07.068.
- Sufen Chen; Jun Liu; Yu Liu; Lijuan Chen; Ting Sun; Nan Yao; Hong-Bin Wang; Bing Liu; BIK1 and ERECTA Play Opposing Roles in Both Leaf and Inflorescence Development in Arabidopsis. Frontiers in Plant Science 2019, 10, 1480, 10.3389/fpls.2019.01480.
- Ive De Smet; Valya Vassileva; Bert De Rybel; Mitchell P. Levesque; Wim Grunewald; Daniel Van Damme; Giel Van Noorden; Mirande Naudts; Gert Van Isterdael; Rebecca De Clercq; et al.Jean Y. WangNicholas MeuliSteffen VannesteJirí FrimlPierre HilsonGerd JürgensGwyneth IngramDirk InzéPhilip N BenfeyTom Beeckman Receptor-Like Kinase ACR4 Restricts Formative Cell Divisions in the Arabidopsis Root. Science 2008, 322, 594-597, 10.1126/science.1160158.
- Yvonne Stahl; René H Wink; Gwyneth Ingram; Rüdiger Simon; A Signaling Module Controlling the Stem Cell Niche in Arabidopsis Root Meristems. Current Biology 2009, 19, 909-914, 10.1016/j.cub.2009.03.060.
- Yvonne Stahl; Stéphanie Grabowski; Andrea Bleckmann; Ralf Kühnemuth; Stefanie Weidtkamp-Peters; Karine Gustavo Pinto; Gwendolyn K. Kirschner; Julia B. Schmid; René H. Wink; Adrian Hülsewede; et al.Suren FelekyanClaus A. M. SeidelRüdiger Simon Moderation of Arabidopsis Root Stemness by CLAVATA1 and ARABIDOPSIS CRINKLY4 Receptor Kinase Complexes. Current Biology 2013, 23, 362-371, 10.1016/j.cub.2013.01.045.
- Ramón Pelagio-Flores; Edith Muñoz-Parra; Salvador Barrera-Ortiz; Randy Ortiz-Castro; Jorge Saenz-Mata; María Azucena Ortega-Amaro; Juan Francisco Jiménez-Bremont; José López‐Bucio; The cysteine-rich receptor-like protein kinase CRK28 modulates Arabidopsis growth and development and influences abscisic acid responses. Planta 2019, 251, 2, 10.1007/s00425-019-03296-y.
- Su-Hwan Kwak; John Schiefelbein; A Feedback Mechanism Controlling SCRAMBLED Receptor Accumulation and Cell-Type Pattern in Arabidopsis. Current Biology 2008, 18, 1949-1954, 10.1016/j.cub.2008.10.064.
- Ram Kishor Yadav; Lynette Fulton; Martine Batoux; Kay Schneitz; The Arabidopsis receptor-like kinase STRUBBELIG mediates inter-cell-layer signaling during floral development. Developmental Biology 2008, 323, 261-270, 10.1016/j.ydbio.2008.08.010.
- Prasad Vaddepalli; A. Herrmann; Lynette Fulton; M. Oelschner; Stefan Hillmer; T. F. Stratil; A. Fastner; Ulrich Hammes; Thomas Ott; David G. Robinson; et al.Kay Schneitz The C2-domain protein QUIRKY and the receptor-like kinase STRUBBELIG localize to plasmodesmata and mediate tissue morphogenesis in Arabidopsis thaliana. Development 2014, 141, 4139-4148, 10.1242/dev.113878.
- Su-Hwan Kwak; John Schiefelbein; The role of the SCRAMBLED receptor-like kinase in patterning the Arabidopsis root epidermis. Developmental Biology 2007, 302, 118-131, 10.1016/j.ydbio.2006.09.009.
- Noriko Shimizu; Takashi Ishida; Masashi Yamada; Shuji Shigenobu; Ryo Tabata; Atsuko Kinoshita; Katsushi Yamaguchi; Mitsuyasu Hasebe; Kanako Mitsumasu; Shinichiro Sawa; et al. BAM 1 and RECEPTOR-LIKE PROTEIN KINASE 2 constitute a signaling pathway and modulate CLE peptide-triggered growth inhibition in Arabidopsis root. New Phytologist 2015, 208, 1104-1113, 10.1111/nph.13520.
- Roya Campos; Jason Goff; Cecilia Rodriguez-Furlan; Jaimie M. Van Norman; The Arabidopsis Receptor Kinase IRK Is Polarized and Represses Specific Cell Divisions in Roots. Developmental Cell 2020, 52, 183-195, 10.1016/j.devcel.2019.12.001.
- Qiaohong Duan; Daniel Kita; Chao Li; Alice Y. Cheung; Hen-Ming Wu; FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proceedings of the National Academy of Sciences 2010, 107, 17821-17826, 10.1073/pnas.1005366107.
- Kate Fisher; Simon R Turner; PXY, a Receptor-like Kinase Essential for Maintaining Polarity during Plant Vascular-Tissue Development. Current Biology 2007, 17, 1061-1066, 10.1016/j.cub.2007.05.049.
- Yuki Hirakawa; Hidefumi Shinohara; Yuki Kondo; Asuka Inoue; Ikuko Nakanomyo; Mari Ogawa; Shinichiro Sawa; Kyoko Ohashi-Ito; Yoshikatsu Matsubayashi; Hiroo Fukuda; et al. Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proceedings of the National Academy of Sciences 2008, 105, 15208-15213, 10.1073/pnas.0808444105.
- Jiehua Wang; Melis Kucukoglu; Linbin Zhang; Peng Chen; Daniel Decker; Ove Nilsson; Brian Jones; Göran Sandberg; Bo Zheng; The Arabidopsis LRR-RLK, PXC1, is a regulator of secondary wall formation correlated with the TDIF-PXY/TDR-WOX4 signaling pathway. BMC Plant Biology 2013, 13, 94, 10.1186/1471-2229-13-94.
- Nial Gursanscky; Virginie Jouannet; Karin Grünwald; Pablo Sánchez; Martina Laaber‐Schwarz; Thomas Greb; MOL1 is required for cambium homeostasis in Arabidopsis. The Plant Journal 2016, 86, 210-220, 10.1111/tpj.13169.
- Anthony C. Bryan; Adam Obaidi; Michael Wierzba; Frans E. Tax; XYLEM INTERMIXED WITH PHLOEM1, a leucine-rich repeat receptor-like kinase required for stem growth and vascular development in Arabidopsis thaliana. Planta 2012, 235, 111-122, 10.1007/s00425-011-1489-6.
- Tsung-Luo Jinn; Julie M. Stone; John C. Walker; HAESA, an Arabidopsis leucine-rich repeat receptor kinase, controls floral organ abscission. Genes & Development 2000, 14, 108-117.
- Grethe-Elisabeth Stenvik; Melinka A. Butenko; Reidunn B. Aalen; Identification of a putative receptor-ligand pair controlling cell separation in plants. Plant Signaling & Behavior 2008, 3, 1109-1110, 10.4161/psb.3.12.7009.
- Isaiah Taylor; Ying Wang; Kati Seitz; John Baer; Stefan Bennewitz; Brian P. Mooney; John C. Walker; Analysis of Phosphorylation of the Receptor-Like Protein Kinase HAESA during Arabidopsis Floral Abscission. PLOS ONE 2016, 11, e0147203, 10.1371/journal.pone.0147203.
- Julia Santiago; Benjamin Brandt; Mari Wildhagen; Ulrich Hohmann; Ludwig A Hothorn; Melinka A Butenko; Michael Hothorn; Mechanistic insight into a peptide hormone signaling complex mediating floral organ abscission. eLife 2016, 5, e15075, 10.7554/eLife.15075.
- Xiaoxu Li; Salman Ahmad; Akhtar Ali; Cun Guo; Hong Li; Jing Yu; Yan Zhang; XiaoMing Gao; Yongfeng Guo; Characterization of Somatic Embryogenesis Receptor-Like Kinase 4 as a Negative Regulator of Leaf Senescence in Arabidopsis. Cells 2019, 8, 50, 10.3390/cells8010050.
- Kyoung Hee Nam; Jianming Li; BRI1/BAK1, a Receptor Kinase Pair Mediating Brassinosteroid Signaling. Cell 2002, 110, 203-212, 10.1016/s0092-8674(02)00814-0.
- Jia Li; Jiangqi Wen; Kevin A Lease; Jason T Doke; Frans E. Tax; John C. Walker; BAK1, an Arabidopsis LRR Receptor-like Protein Kinase, Interacts with BRI1 and Modulates Brassinosteroid Signaling. Cell 2002, 110, 213-222, 10.1016/s0092-8674(02)00812-7.
- Aifen Zhou; Huachun Wang; John C. Walker; Jia Li; BRL1, a leucine‐rich repeat receptor‐like protein kinase, is functionally redundant with BRI1 in regulating Arabidopsis brassinosteroid signaling. The Plant Journal 2004, 40, 399-409, 10.1111/j.1365-313x.2004.02214.x.
- Min Hee Kim; Yoon Kim; Ju Won Kim; Hyun-Suk Lee; Woo Sung Lee; Seong-Ki Kim; Zhi-Yong Wang; Soo-Hwan Kim; Identification of Arabidopsis BAK1-Associating Receptor-Like Kinase 1 (BARK1) and Characterization of its Gene Expression and Brassinosteroid-Regulated Root Phenotypes. Plant and Cell Physiology 2013, 54, 1620-1634, 10.1093/pcp/pct106.
- Michael P. Wierzba; Frans E. Tax; An Allelic Series of bak1 Mutations Differentially Alter bir1 Cell Death, Immune Response, Growth, and Root Development Phenotypes in Arabidopsis thaliana. Genetics 2015, 202, 689-702, 10.1534/genetics.115.180380.
- Xiaoping Gou; Hongju Yin; Kai He; Junbo Du; Jing Yi; Shengbao Xu; Honghui Lin; Steven D. Clouse; Jia Li; Genetic Evidence for an Indispensable Role of Somatic Embryogenesis Receptor Kinases in Brassinosteroid Signaling. PLOS Genetics 2012, 8, e1002452, 10.1371/journal.pgen.1002452.
- Dhinesh Kumar; Ritesh Kumar; Dongwon Baek; Tae-Kyung Hyun; Woo Sik Chung; Dae-Jin Yun; Jae-Yean Kim; Arabidopsis thaliana RECEPTOR DEAD KINASE1 Functions as a Positive Regulator in Plant Responses to ABA. Molecular Plant 2017, 10, 223-243, 10.1016/j.molp.2016.11.011.
- Ling Bai; Guozeng Zhang; Yun Zhou; Zhaopei Zhang; Wei Wang; Yanyan Du; Zhongyi Wu; Chun-Peng Song; Plasma membrane-associated proline-rich extensin-like receptor kinase 4, a novel regulator of Ca2+signalling, is required for abscisic acid responses inArabidopsis thaliana. The Plant Journal 2009, 60, 314-327, 10.1111/j.1365-313x.2009.03956.x.


