 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | You-Lin Tain | + 1882 word(s) | 1882 | 2021-06-09 11:52:46 | | | |
| 2 | Camila Xu | Meta information modification | 1882 | 2021-06-11 02:59:55 | | | | |
| 3 | Camila Xu | + 657 word(s) | 2539 | 2021-06-11 03:01:43 | | | | |
| 4 | Camila Xu | Meta information modification | 2539 | 2021-06-11 03:02:33 | | |
Video Upload Options
Diverse animal models resembling human illnesses and pregnancy complications have been evaluated, such as hypertensive disorders of pregnancy , preeclampsia , chronic kidney disease , diabetes , polycystic ovary syndrome , maternal inflammation , maternal hypoxia , and sleep disorder .
1. Introduction
The association between fetal development and the increased risk of adult disease has attracted a great deal of attention to the concept of developmental programming or developmental origins of health and disease (DOHaD) These findings, combined with numerous subsequent epidemiologic investigations, indicate that the perinatal period, a critical window of organogenesis, is a vulnerable time in terms of the impact of adverse environmental insults [1]. Several hypotheses, such as thrifty phenotype [2], maternal capital [3], and predictive adaptive responses [4], have been developed to explain the epidemiological observations of an association between early life insults and diseases in adulthood. However, these hypotheses do not propose mechanistic pathways by which disease proceeds or suggest potential interventions for the prevention of adult diseases.
Hypertension and related cardiovascular diseases are leading causes of mortality worldwide [5]. The WHO reported that 1 in 4 men and 1 in 5 women have hypertension [6]. Due to the multifactorial nature of hypertension, the use of various animal models, which induce hypertension by various mechanisms and produce the same end result, is advantageous [7][8]. In the past decades, novel drug classes and interventional strategies for the treatment of hypertension have been developed using hypertensive animal models [9].
These include maternal undernutrition or overnutrition, maternal disease states, lifestyle changes, substance abuse, environmental exposure to toxins/chemicals, and medication use during pregnancy [10][11][12][13][14]. Hypertension, diabetes, kidney disease, and inflammation are common maternal diseases that complicate pregnancy. On the other hand, programming processes geared toward disease could be reversed by shifting therapy from adulthood to the perinatal period, that is to say, by reprogramming [15]. Although the pathogenesis behind hypertension of developmental origins is poorly understood at present, our understanding of animal models used to study common mechanistic pathways has advanced greatly in recent years, which helps in developing efficient strategies to reprogram hypertension and prevent it from happening.
This review summarizes the contributions of animal models to DOHaD research with a focus on hypertension. It is proposed that integrating evidence from diverse animal models is essential in order to advance our understanding of hypertension of developmental origins and develop novel reprogramming strategies to alleviate the global burden of hypertension.
We retrieved related literature from all articles indexed in PubMed/MEDLINE. “blood pressure”, “developmental programming”, “DOHaD”, “animal model”, “mother”, “maternal”, “pregnancy”, “gestation”, “offspring”, “progeny”, “prenatal”, “perinatal”, “reprogramming”, and “hypertension”. Additional studies were then selected and assessed based on appropriate references in eligible papers.
2. Choice of Animal Models
A broad range of animal models have been established to validate that the associations found in human observational studies can be replicated under experimental conditions. Second, animal models can be classified according to molecular mechanisms. To date, hypertension of developmental origins has been attributed to mechanisms [10][11][12][13][14][15] including reduced nephron number, oxidative stress, an aberrant renin–angiotensin system (RAS), gut microbiota dysbiosis, and sex differences, among others. A summary of the selection of animal models for the study of hypertension of developmental origins is depicted in Figure 1.
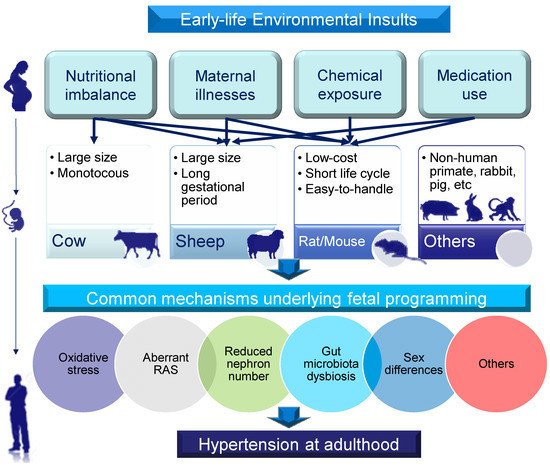
Figure 1. Schematic illustration of the selection of animal models for studying hypertension of developmental origins in adulthood according to early-life environmental insults, animal species, and common mechanisms. Lines with arrows (top section) indicate types of early-life insults produced in particular species of animals to induce hypertension in adult offspring. The study of other animals in DOHaD research (non-human primates, rabbits, pigs, etc.) is limited.
3. Selection of Appropriate Animal Models to Study Hypertension of Developmental Origins
Even though significant advances have been made in developing diverse animal models to study hypertension of developmental origins, the need for meaningful clinical translation remains a research priority. The following conditions should be taken into consideration when we select animal models. Second, the gestation period and litter size should be comparable to those of humans. Third, it is crucial that animal models share similar features of adverse outcomes to those seen in human studies, which can be measured.
Across different species, critical development periods for major organ systems are not uniform. Blood pressure is tightly controlled by coordination among the kidney, heart, brain, and other organ systems. As such, the translatability of studies performed in animals should be approached with caution, as many key stages of BP-controlled organ development that occur before birth in humans occur after birth in some species [16][17][18][19].
Many animal studies on hypertension of developmental origins focus on renal programming [20][18]. Unlike in humans, rat kidneys continue to develop after birth and complete at 1 to 2 weeks postnatally [21]. Accordingly, adverse environmental conditions during pregnancy as well as lactation can impair kidney development, consequently resulting in hypertension in rodents [20]. For example, repeated dexamethasone administration on embryonic days 15 and 16 [22], from gestational days 16 to 22 [23], or from postnatal days 1 to 3 [24] was associated with developmental programming of hypertension in adult rat offspring.
Another unsolved problem is that almost no studies have taken a comprehensive approach to simultaneously evaluating every BP-controlled organ system in response to in utero exposure at specific developmental stages to assess their relative vulnerability in an experiment. Due to the complex nature of the interplay between organogenesis and environmental insults, the programming effect on various organs might be dissimilar in different animal species. Hence, it is apparent that the selected animal paradigm should mirror the timing of human organ development as closely as possible so that the effects of early-life insults can be fully assessed.
The advantages of a shorter gestation period and higher offspring yield compared to large animal models make rodent models the most commonly used in DOHaD research. There is a large set of studies on hypertension of developmental origins that were carried out in rats (Table 1 and Table 2). If an early-life insult is induced by surgical manipulation or if delivery requires repeated procedures, short gestation in rodents could become disadvantageous. In addition, the short gestation time may not allow for the permanent resolution of developmental plasticity and the identification of critical time periods that are vulnerable to insults.
Table 1. Summary of animal models of the developmental programming of hypertension, categorized according to maternal illness and pregnancy complications.
| Maternal Illnesses and Conditions | Animal Models | Species/Gender | Age at Hypertension Development | Ref. |
|---|---|---|---|---|
| Hypertensive disorders of pregnancy | Genetic hypertension model | SHR/M | 12 weeks | [25] |
| 2-kidney, 1-clip renovascular hypertension model | SD rat/M,F | 16 weeks | [26] | |
| Preeclampsia | Intraperitoneal administration of 60 mg/kg suramin on gestational days 10 and 11 | SD rat/M | 12 weeks | [27] |
| Subcutaneous administration of 60 mg/kg L-NAME during pregnancy | SD rat/M | 12 weeks | [28] | |
| Reduced uterine perfusion | SD rat/M | 16 weeks | [29] | |
| Chronic kidney disease | 0.5% adenine supplementation from 3 weeks before pregnancy until 3 weeks after delivery | SD rat/M | 12 weeks | [30] |
| Type 1 diabetes | Single intraperitoneal injection of 45 mg/kg STZ on gestational day 0 | SD rat/M | 12 weeks | [31] |
| Single intraperitoneal injection of 35 mg/kg STZ on gestational day 0 | SD rat/M | 6 months | [32] | |
| Type 2 diabetes | Mother rat received single intraperitoneal injection of 50 mg/kg STZ at newborn stage | SD rat/M | 12 weeks | [31] |
| Anemia | Iron-deficiency diet from 4 weeks before pregnancy until delivery | Rowett hooded Lister rat/M & F | 16 weeks | [33] |
| Polycystic ovary syndrome | Subcutaneous injection of 5 mg/kg testosterone cypionate on gestational day 20 | Wistar rat/F | 120 days | [34] |
| Maternal inflammation | Intraperitoneal administration of 0.79 mg/kg LPS on gestational days 8, 10, and 12 | SD rat/M & F | 12 weeks | [35] |
| Intraperitoneal injection of 2.37 mg/kg zymosan on gestation days 8, 10, and 12 | SD rat/M | 66 weeks | [36] | |
| Maternal hypoxia | Hypoxia maintained at constant inspired fraction of oxygen of 13% from gestational day 6 to 20 | Wistar rat/M | 4 months | [37] |
| Hypoxia maintained at 10% oxygen from gestational day 105 to 145 | Sheep/F | 9 months | [38] | |
| Sleep disorder | Sleep restriction | Wistar rat/M | 3 months | [39] |
| 24 h constant light exposure during pregnancy | SD rat/M | 12 weeks | [40] |
Studies tabulated according to types of maternal illnesses and conditions, animal model, and age at evaluation. L−NAME = NG-nitro-L-arginine-methyl ester; STZ = streptozotocin; LPS = lipopolysaccharide; SHR = spontaneously hypertensive rat; SD = Sprague-Dawley.
Table 2. Summary of animal models of the developmental programming of hypertension, categorized according to chemical and medication exposure.
| Chemical or Medication | Animal Models | Species/Gender | Age at Hypertension Development | Ref. |
|---|---|---|---|---|
| TCDD | Oral administration of 200 ng/kg TCDD on gestational days 14 and 21 and postnatal days 7 and 14 | SD rat/M | 12 weeks | [41] |
| Oral administration of 200 ng/kg TCDD on gestational days 14 and 21 and postnatal days 7 and 14 | SD rat/M | 16 weeks | [42] | |
| Bisphenol A | Oral administration of 50 μg/kg/day bisphenol A during pregnancy and lactation | SD rat/M | 16 weeks | [43] |
| Nicotine | Nicotine administration via osmotic mini-pump at 4 μg/kg/min from gestational day 4 to postnatal day 10 | SD rat/M | 8 months | [44] |
| Alcohol | Ethanol 1 g/kg by oral gavage on gestational days 13.5 and 14.5 | SD rat/M,F | 6 months | [45] |
| Caffeine | Subcutaneous injection of 20 mg/kg caffeine daily during pregnancy | C57BL/6 mouse/M | 3 months | [46] |
| Cyclosporine | Cyclosporine 3.3 mg/kg from gestational day 10 to postnatal day 7 | SD rat/M | 11 weeks | [47] |
| Gentamicin | Subcutaneous injection of 110 mg/kg gentamicin from gestational day 10 to 15 or 15 to 20 | SD rat/F | 1 year | [48] |
| Minocycline | Minocycline 50 mg/kg via oral gavage during pregnancy and lactation | SD rat/M | 12 weeks | [49] |
| Tenofovir | Tenofovir 100 mg/kg diet from 1 week before mating and during pregnancy | Wistar rat/M | 6 months | [50] |
| Glucocorticoid | Intraperitoneal injection of 0.2 mg/kg dexamethasone on gestational days 15 and 16 | SD rat/M | 12 weeks | [22] |
| Intraperitoneal injection of 0.1 mg/kg dexamethasone from gestational day 16 to 22 | SD rat/M | 12 weeks | [23] | |
| Intraperitoneal injection of 0.5 mg/kg dexamethasone on postnatal day 1, 0.3 mg/kg on day 2, and 0.1 mg/kg on day 3. | SD rat/M | 12 weeks | [24] | |
| Intramuscular injection of 0.17 mg/kg betamethasone on gestational days 80 and 81 | Sheep/M,F | 18 months | [51] | |
| Intravenous treatment with 0.48 mg/h dexamethasone for 48 h on gestational day 27 | Sheep/M,F | 16 months | [52] |
Studies tabulated according to type of chemical or medication, animal model, and age at evaluation. TCDD = 2,3,7,8-tetrachlorodibenzo-p-dioxin; SD = Sprague-Dawley.
Unlike humans, rodents generally have more than one offspring, and litter sizes of 8–12 pups are usually seen. Such a large litter size is also a disadvantage when compared to singleton births common in humans and large animal models. Accordingly, normalizing the size of each litter after birth should be considered to control for differences in offspring food intake, maternal care, and pup growth [53]. Since these limitations exist, the complete translation of findings in rodents to human medicine is seriously compromised.
On the other hand, gestational length in sheep is around 150 days, during which the fetal size and development rate are similar to those of humans [54]. With the use of ewe models, maternal caloric restriction [55], maternal hypoxia [38], and prenatal glucocorticoid exposure [51][52] have been shown to cause hypertension in adult progeny. Although these early insults have shown the same adverse effects on offspring BP in sheep and rats, whether different gestation periods and litter sizes differentially impact hypertension of developmental origins in rats and large animals awaits further evaluation.
As we mentioned earlier, rats are the most commonly used species for the developmental programming of hypertension. However, a critical assessment of the data show that this phenomenon is mostly observed when BP is typically measured by the tail cuff method; in contrast, hypertension is not detected in telemetrically instrumented animals [56]. Although BP data obtained from the tail cuff method are reported to correlate well with findings of direct arterial catheter and telemetry methods [57], part of the increased BP in offspring found after early-life insults may be due to an increased stress response related to sympathetic nerve activity.
In adulthood, one rat month is roughly equivalent to three human years [58]. Accordingly, Table 1 lists the timing of hypertension development measured in rats from 12 weeks to 8 months of age, which is equivalent to humans of a specific age group ranging from childhood to early adulthood. Thus, there remain gaps in our knowledge regarding the long-term adverse effects of maternal insults on BP in older adult offspring.
Several species have been studied for cardiovascular outcomes programmed by maternal adverse exposure, including guinea pigs [59], swine [60], and non-human primates [61]. However, none of them have been used to study hypertension of developmental origins. It is important to remember that large animals should not be neglected, as they are generally more physiologically suitable models with regard to human conditions.
In the current review, the wide range of early-life insults certainly influenced the outcomes, resulting in the reported heterogeneity. The results depended strongly on the applied measurement technique and animal model. A huge percentage of studies employed male-only small animal models with small sample sizes. Future animal studies should improve the methodological quality by applying randomization, blinding, and sample size calculation techniques in order to avoid bias and collect data of better quality.
Currently, reprogramming strategies could be categorized as nutritional intervention, lifestyle modification, or pharmacological therapy. It stands to reason that avoiding in utero exposure to adverse conditions is the most effective strategy for preventing hypertension of developmental origins. Another approach is the use of nutritional intervention during pregnancy and lactation [62]. Although the targeting of specific nutrients as a reprogramming strategy opens a new avenue for prevention [63], there remains a lack of accurate dietary recommendations for specific nutritional requirements for pregnant women in case of deficiencies [64][65].
Research on short-lived rodent models has provided significant results, revealing potential pharmacological therapies for preventing hypertension of developmental origins. The standardization of animal experiments will improve the comparability of such studies. During the preparation of the current review, we found that almost no studies tested different doses or the use of different species. Additionally, the follow-up period after the cessation of interventions in most cited studies was rather short.
The efficacy of the intervention can be influenced by its duration with respect to organ development in a dose- and species-specific manner. Thus, further translational research into the pharmacokinetics and metabolism of pharmacological intervention is required to validate and compare its safety and therapeutic potential between humans and other species.
4. Conclusions and Future Perspectives
Various small (e.g., rat and mouse) and large (e.g., cow and sheep) animal models have made important contributions to the DOHaD research field, giving rise to convincing evidence of a causal relationship between various early-life insults and the risk of developing hypertension in later life. Our review highlights that animal models are not only used to investigate the mechanisms behind hypertension of developmental origins, but also have an impact on the development of early-life interventions as a reprogramming strategy to prevent the development of hypertension in adulthood.
There are still several questions that need to be answered. Additionally, this review did not consider the potential for the programming of hypertension by paternal factors that clearly exist in the DOHaD field [66]. Moreover, little reliable information currently exists with regard to optimal doses and durations of pharmacological interventions for pregnant women and the long-term effects on their offspring. Currently, preventive strategies should focus on avoiding exposure to theoretically harmful agents perinatally and promoting a healthy lifestyle.
Each of the abovementioned animal models was used to study a specific hypothesis and neither can be considered superior with regard to all aspects of research on hypertension of developmental origins. Therefore, further research is needed to gain a better understanding of the types of early-life insults, other mechanisms behind hypertension of developmental origins, the ideal therapeutic dose and duration of early intervention, and the appropriate animal species. It is proposed that taking a DOHaD approach with maximum use of the animal evidence should be of benefit in reducing the global burden of hypertension.
References
- Hanson, M.; Gluckman, P. Developmental origins of noncommunicable disease: Population and public health implications. Am. J. Clin. Nutr. 2011, 94, 1754S–1758S.
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20.
- Wells, J.C. Maternal capital and the metabolic ghetto: An evolutionary perspective on the transgenerational basis of health inequalities. Am. J. Hum. Biol. 2010, 22, 1–17.
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736.
- Bromfield, S.; Muntner, P. High blood pressure: The leading global burden of disease risk factor and the need for worldwide prevention programs. Curr. Hypertens. Rep. 2013, 15, 134–136.
- World Health Organization. Hypertension. 2019. Available online: (accessed on 13 April 2021).
- Lerman, L.O.; Kurtz, T.W.; Touyz, R.M.; Ellison, D.H.; Chade, A.R.; Crowley, S.D.; Mattson, D.L.; Mullins, J.J.; Osborn, J.; Eirin, A.; et al. Animal Models of Hypertension: A Scientific Statement From the American Heart Association. Hypertension 2019, 73, e87–e120.
- Pinto, Y.M.; Paul, M.; Ganten, D. Lessons from rat models of hypertension: From Goldblatt to genetic engineering. Cardiovasc. Res. 1998, 39, 77–88.
- Oparil, S.; Schmieder, R.E. New approaches in the treatment of hypertension. Circ. Res. 2015, 116, 1074–1095.
- Ojeda, N.B.; Grigore, D.; Alexander, B.T. Developmental programming of hypertension: Insight from animal models of nutritional manipulation. Hypertension 2008, 52, 44–50.
- Bagby, S.P. Maternal nutrition, low nephron number, and hypertension in later life: Pathways of nutritional programming. J. Nutr. 2007, 137, 1066–1072.
- Chong, E.; Yosypiv, I.V. Developmental programming of hypertension and kidney disease. Int. J. Nephrol. 2012, 2012, 760580.
- Paixão, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 144.
- Hsu, C.N.; Tain, Y.L. Early Origins of Hypertension: Should Prevention Start Before Birth Using Natural Antioxidants? Antioxidants 2020, 9, 1034.
- Tain, Y.L.; Joles, J.A. Reprogramming: A Preventive Strategy in Hypertension Focusing on the Kidney. Int. J. Mol. Sci. 2015, 17, 23.
- Hsu, C.N.; Tain, Y.L. Developmental Origins of Kidney Disease: Why Oxidative Stress Matters? Antioxidants 2021, 10, 33.
- Tain, Y.L.; Hsu, C.N. Interplay between oxidative stress and nutrient sensing signaling in the developmental origins of cardiovascular disease. Int. J. Mol. Sci. 2017, 18, 841.
- Kett, M.M.; Denton, K.M. Renal programming: Cause for concern? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R791–R803.
- Hunter, D.S.; Hazel, S.J.; Kind, K.L.; Owens, J.A.; Pitcher, J.B.; Gatford, K.L. Programming the brain: Common outcomes and gaps in knowledge from animal studies of IUGR. Physiol. Behav. 2016, 164, 233–248.
- Tain, Y.L.; Chan, S.H.H.; Chan, J.Y.H. Biochemical basis for pharmacological intervention as a reprogramming strategy against hypertension and kidney disease of developmental origin. Biochem. Pharmacol. 2018, 153, 82–90.
- Hartman, H.A.; Lai, H.L.; Patterson, L.T. Cessation of renal morphogenesis in mice. Dev. Biol. 2007, 310, 379–387.
- Tain, Y.L.; Sheen, J.M.; Chen, C.C.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free Radic. Res. 2014, 48, 580–586.
- Tai, I.H.; Sheen, J.M.; Lin, Y.J.; Yu, H.R.; Tiao, M.M.; Chen, C.C.; Huang, L.T.; Tain, Y.L. Maternal N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and prevents programmed hypertension in male offspring exposed to prenatal dexamethasone and postnatal high-fat diet. Nitric Oxide 2016, 53, 6–12.
- Chang, H.Y.; Tain, Y.L. Postnatal dexamethasone-induced programmed hypertension is related to the regulation of melatonin and its receptors. Steroids 2016, 108, 1–6.
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal N-Acetylcysteine Therapy Prevents Hypertension in Spontaneously Hypertensive Rat Offspring: Implications of Hydrogen Sulfide-Generating Pathway and Gut Microbiota. Antioxidants 2020, 9, 856.
- Guo, Q.; Feng, X.; Xue, H.; Jin, S.; Teng, X.; Duan, X.; Xiao, L.; Wu, Y. Parental Renovascular Hypertension-Induced Autonomic Dysfunction in Male Offspring Is Improved by Prenatal or Postnatal Treatment With Hydrogen Sulfide. Front. Physiol. 2019, 10, 1184.
- Tain, Y.L.; Hsu, C.N.; Lee, C.T.; Lin, Y.J.; Tsai, C.C. N-Acetylcysteine Prevents Programmed Hypertension in Male Rat Offspring Born to Suramin-Treated Mothers. Biol. Reprod. 2016, 95, 8.
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal N(G)-Nitro-L-arginine methyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636.
- Ojeda, N.B.; Hennington, B.S.; Williamson, D.T.; Hill, M.L.; Betson, N.E.; Sartori-Valinotti, J.C.; Reckelhoff, J.F.; Royals, T.P.; Alexander, B.T. Oxidative stress contributes to sex differences in blood pressure in adult growth-restricted offspring. Hypertension 2012, 60, 114–122.
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Adenine-Induced Chronic Kidney Disease Programs Hypertension in Adult Male Rat Offspring: Implications of Nitric Oxide and Gut Microbiome Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 7237.
- Tain, Y.L.; Lee, W.C.; Hsu, C.N.; Lee, W.C.; Huang, L.T.; Lee, C.T.; Lin, C.Y. Asymmetric dimethylarginine is associated with developmental programming of adult kidney disease and hypertension in offspring of streptozotocin-treated mothers. PLoS ONE 2013, 8, e55420.
- Dib, A.; Payen, C.; Bourreau, J.; Munier, M.; Grimaud, L.; Fajloun, Z.; Loufrani, L.; Henrion, D.; Fassot, C. In Utero Exposure to Maternal Diabetes Is Associated With Early Abnormal Vascular Structure in Offspring. Front. Physiol. 2018, 9, 350.
- Gambling, L.; Dunford, S.; Wallace, D.I.; Zuur, G.; Solanky, N.; Srai, K.S.; McArdle, H.J. Iron deficiency during pregnancy affects post-natal blood pressure in the rat. J. Physiol. 2003, 552, 603–610.
- Sherman, S.B.; Sarsour, N.; Salehi, M.; Schroering, A.; Mell, B.; Joe, B.; Hill, J.W. Prenatal androgen exposure causes hypertension and gut microbiota dysbiosis. Gut Microbes 2018, 9, 400–421.
- Wang, J.; Yin, N.; Deng, Y.; Wei, Y.; Huang, Y.; Pu, X.; Li, L.; Zheng, Y.; Guo, J.; Yu, J.; et al. Ascorbic Acid Protects against Hypertension through Downregulation of ACE1 Gene Expression Mediated by Histone Deacetylation in Prenatal Inflammation-Induced Offspring. Sci. Rep. 2016, 6, 39469.
- Liao, W.; Wei, Y.; Yu, C.; Zhou, J.; Li, S.; Pang, Y.; Li, G.; Li, X. Prenatal exposure to zymosan results in hypertension in adult offspring rats. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1413–1418.
- Giussani, D.A.; Camm, E.J.; Niu, Y.; Richter, H.G.; Blanco, C.E.; Gottschalk, R.; Blake, E.Z.; Horder, K.A.; Thakor, A.S.; Hansell, J.A.; et al. Developmental programming of cardiovascular dysfunction by prenatal hypoxia and oxidative stress. PLoS ONE 2012, 7, e31017.
- Brain, K.L.; Allison, B.J.; Niu, Y.; Cross, C.M.; Itani, N.; Kane, A.D.; Herrera, E.A.; Skeffington, K.L.; Botting, K.J.; Giussani, D.A. Intervention against hypertension in the next generation programmed by developmental hypoxia. PLoS Biol. 2019, 17, e2006552.
- Thomal, J.T.; Palma, B.D.; Ponzio, B.F.; Franco Mdo, C.; Zaladek-Gil, F.; Fortes, Z.B.; Tufik, S.; Gomes, G.N. Sleep restriction during pregnancy: Hypertension and renal abnormalities in young offspring rats. Sleep 2010, 33, 1357–1362.
- Tain, Y.L.; Lin, Y.J.; Chan, J.Y.H.; Lee, C.T.; Hsu, C.N. Maternal melatonin or agomelatine therapy prevents programmed hypertension in male offspring of mother exposed to continuous light. Biol. Reprod. 2017, 97, 636–643.
- Hsu, C.N.; Chan, J.Y.H.; Yu, H.R.; Lee, W.C.; Wu, K.L.H.; Chang-Chien, G.P.; Lin, S.; Hou, C.Y.; Tain, Y.L. Targeting on Gut Microbiota-Derived Metabolite Trimethylamine to Protect Adult Male Rat Offspring against Hypertension Programmed by Combined Maternal High-Fructose Intake and Dioxin Exposure. Int. J. Mol. Sci. 2020, 21, 5488.
- Hsu, C.N.; Lin, Y.J.; Lu, P.C.; Tain, Y.L. Maternal resveratrol therapy protects male rat offspring against programmed hypertension induced by TCDD and dexamethasone exposures: Is it relevant to aryl hydrocarbon receptor? Int. J. Mol. Sci. 2018, 19, 2459.
- Hsu, C.N.; Lin, Y.J.; Tain, Y.L. Maternal exposure to bisphenol A combined with high-fat diet-induced programmed hypertension in adult male rat offspring: Effects of resveratrol. Int. J. Mol. Sci. 2019, 20, 4382.
- Xiao, D.; Huang, X.; Li, Y.; Dasgupta, C.; Wang, L.; Zhang, L. Antenatal Antioxidant Prevents Nicotine-Mediated Hypertensive Response in Rat Adult Offspring. Biol. Reprod. 2015, 93, 66.
- Gray, S.P.; Denton, K.M.; Cullen-McEwen, L.; Bertram, J.F.; Moritz, K.M. Prenatal exposure to alcohol reduces nephron number and raises blood pressure in progeny. J. Am. Soc. Nephrol. 2010, 21, 1891–1902.
- Serapiao-Moraes, D.F.; Souza-Mello, V.; Aguila, M.B.; Mandarim-de-Lacerda, C.A.; Faria, T.S. Maternal caffeine administration leads to adverse effects on adult mice offspring. Eur. J. Nutr. 2013, 52, 1891–1900.
- Slabiak-Blaz, N.; Adamczak, M.; Gut, N.; Grajoszek, A.; Nyengaard, J.R.; Ritz, E.; Wiecek, A. Administration of cyclosporine a in pregnant rats—The effect on blood pressure and on the glomerular number in their offspring. Kidney Blood Press. Res. 2015, 40, 413–423.
- Chahoud, I.; Stahlmann, R.; Merker, H.J.; Neubert, D. Hypertension and nephrotoxic lesions in rats 1 year after prenatal expo-sure to gentamicin. Arch. Toxicol. 1988, 62, 274–284.
- Hsu, C.N.; Chan, J.Y.H.; Wu, K.L.H.; Yu, H.R.; Lee, W.C.; Hou, C.Y.; Tain, Y.L. Altered Gut Microbiota and Its Metabolites in Hypertension of Developmental Origins: Exploring Differences between Fructose and Antibiotics Exposure. Int. J. Mol. Sci. 2021, 22, 2674.
- Gois, P.H.; Canale, D.; Luchi, W.M.; Volpini, R.A.; Veras, M.M.; Costa Nde, S.; Shimizu, M.H.; Seguro, A.C. Tenofovir during pregnancy in rats: A novel pathway for programmed hypertension in the offspring. J. Antimicrob. Chemother. 2015, 70, 1094–1105.
- Gwathmey, T.M.; Shaltout, H.A.; Rose, J.C.; Diz, D.I.; Chappell, M.C. Glucocorticoid-induced fetal programming alters the functional complement of angiotensin receptor subtypes within the kidney. Hypertension 2011, 57, 620–626.
- Dodic, M.; Abouantoun, T.; O’Connor, A.; Wintour, E.M.; Moritz, K.M. Programming effects of short prenatal exposure to dexamethasone in sheep. Hypertension 2002, 40, 729–734.
- Chahoud, I.; Paumgartten, F.J.R. Influence of litter size on the postnatal growth of rat pups: Is there a rationale for litter-size standardization in toxicity studies? Environ. Res. 2009, 109, 1021–1027.
- Barry, J.S.; Anthony, R.V. The pregnant sheep as a model for human pregnancy. Theriogenology 2008, 69, 55–67.
- Gopalakrishnan, G.S.; Gardner, D.S.; Rhind, S.M.; Rae, M.T.; Kyle, C.E.; Brooks, A.N.; Walker, R.M.; Ramsay, M.M.; Keisler, D.H.; Stephenson, T.; et al. Programming of adult cardiovascular function after early maternal undernutrition in sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R12–R20.
- Van Abeelen, A.F.; Veenendaal, M.V.; Painter, R.C.; de Rooij, S.R.; Thangaratinam, S.; van der Post, J.A.; Bossuyt, P.M.; Elias, S.G.; Uiterwaal, C.S.; Grobbee, D.E.; et al. The fetal origins of hypertension: A systematic review and meta-analysis of the evidence from animal experiments of maternal undernutrition. J. Hypertens. 2012, 30, 2255–2267.
- Krege, J.H.; Hodgin, J.B.; Hagaman, J.R.; Smithies, O. A noninvasive computerized tail-cuff system for measuring blood pressure in mice. Hypertension 1995, 25, 1111–1115.
- Sengupta, P. The Laboratory Rat: Relating Its Age with Human’s. Int. J. Prev. Med. 2013, 4, 624–630.
- Morrison, J.L.; Botting, K.J.; Darby, J.R.T.; David, A.L.; Dyson, R.M.; Gatford, K.L.; Gray, C.; Herrera, E.A.; Hirst, J.J.; Kim, B.; et al. Guinea pig models for translation of the developmental origins of health and disease hypothesis into the clinic. J. Physiol. 2018, 596, 5535–5569.
- Gonzalez-Bulnes, A.; Astiz, S.; Ovilo, C.; Lopez-Bote, C.J.; Torres-Rovira, L.; Barbero, A.; Ayuso, M.; Garcia-Contreras, C.; Vazquez-Gomez, M. Developmental Origins of Health and Disease in swine: Implications for animal production and biomedical research. Theriogenology 2016, 86, 110–119.
- Kuo, A.H.; Li, C.; Li, J.; Huber, H.F.; Nathanielsz, P.W.; Clarke, G.D. Cardiac remodeling in a baboon model of intrauterine growth restriction mimics accelerated ageing. J. Physiol. 2017, 595, 1093–1110.
- Hsu, C.N.; Tain, Y.L. The Good, the Bad, and the Ugly of Pregnancy Nutrients and Developmental Programming of Adult Disease. Nutrients 2019, 11, 894.
- Hsu, C.N.; Tain, Y.L. The Double-Edged Sword Effects of Maternal Nutrition in the Developmental Programming of Hypertension. Nutrients 2018, 10, 1917.
- Haider, B.A.; Bhutta, Z.A. Multiple-micronutrient supplementation for women during pregnancy. Cochrane Database Syst. Rev. 2017, 4, CD004905.
- Schwarzenberg, S.J.; Georgieff, M.K.; Committee on Nutrition. Advocacy for Improving Nutrition in the First 1000 Days to Support Childhood Development and Adult Health. Pediatrics 2018, 141, e20173716.
- Safi-Stibler, S.; Gabory, A. Epigenetics and the Developmental Origins of Health and Disease: Parental environment signalling to the epigenome, critical time windows and sculpting the adult phenotype. Semin. Cell Dev. Biol. 2020, 97, 172–180.



