 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Gyu-Jin Rho | + 3360 word(s) | 3360 | 2021-04-14 08:39:13 | | | |
| 2 | Conner Chen | Meta information modification | 3360 | 2021-06-10 12:26:37 | | |
Video Upload Options
In the present era, infertility is one of the major issues which restricts many couples to have their own children. Infertility is the inability to achieve a clinical pregnancy after regular unprotected sexual intercourse for the period of one year or more. Various factors including defective male or female germ cell development, unhealthy and improper lifestyles, diseases like cancer and associated chemo-or-radiation therapies, congenital disorders, etc., may be responsible for infertility. Therefore, it is highly important to understand the basic concepts of germ cell development including primordial germ cell (PGC) formation, specification, migration, entry to genital ridges and their molecular mechanisms, activated pathways, paracrine and autocrine signaling, along with possible alteration which can hamper germ cell development and can cause adversities like cancer progression and infertility. Knowing all these aspects in a proper way can be very much helpful in improving our understanding about gametogenesis and finding possible ways to cure related disorders.
1. Female Germ Cell: In Vivo Developmental Stages (Primordial Germ Cells to Oocyte)
Before attaining full maturity, female germ cells go through various developmental stages. Brief description about these stages is as follows:
1.1. Primordial Germ Cell Development
Mammalian germ cell development begins in the form of PGCs which are the embryonic precursors of gametes [1][2] which on later stages becomes male or female gametes often termed as sperm and oocyte, respectively. Here in this review we will mainly discuss both the human and mouse female germ cell development in vivo and in vitro. A number of developmental changes initiated from proximal epiblast cells, later accompanied by attainment of various markers expression under different pathway activations are held responsible for developing a mature sperm or oocyte with functional ability. The developmental route mainly comprised of following steps: gastrulation → epiblast cells formation → primordial germ cell → gonocyte → oogonium → primary oocyte → secondary oocyte → antral follicle → mature oocyte. Earlier developmental processes are under the control of both of the WNT3 and bone morphogenic protein (BMP) family transcription factors and activation of pathways associated with them. In the mouse at around E5.75, PGCs formation is governed by WNT3 secretion by posterior visceral endoderm (VE) and posterior epiblast cells which is further followed by secretion of BMP family member proteins such as BMP4 and BMP8b by extraembryonic ectoderm (ExE) at around E6.0 [1]. WNT3 signaling regulates PGC induction by its downstream target Brachyury, a mesodermal marker which promotes the expression of both somatic as well as germ cell markers, whereas BMP4 governs the suppression of somatic genes while up regulating germline determinant genes Blimp1 and Prdm14 [3]. BMP8b also plays important role in mouse PGC generation. In a mouse study, null mutations in BMP8b were shown to cause complete lack of PGCs whereas reduced number of PGCs were observed in BMP8b heterozygotes [4]. Many other reports have also shown severe reduction in the number of PGCs from mice embryos in which targeted deletion of either BMP2, BMP4, and BMP8b or their downstream signaling components including Smad1, 4, 5 was performed [5][6][7][8][9]. Similarly, no PRDM1 positive PGCs were observed in mutant mouse embryos which undergo targeted deletion of Wnt3 [3]. The Wnt signaling pathway involving Wnt5a and its receptor Ror2 has also been associated with successful migration and functionality of PGCs [10]. These findings strongly advocate the utility and role of BMP and Wnt signaling pathways in development, migration and proper functioning of the developing PGCs which further determines the specification and development of gametes. It is worthy to notice the indispensable role of Fragilis, Smad1, and Stella which helps in the specification of PGCs and also provides distinction from somatic cells [11][12][13]. Any deformity or suppression of these valuable factors results into impaired PGC/gamete production and hampers their functional ability. Beginning with a small number of ~30–40 cells, PGCs proliferate and finally migrates towards genital ridges where they undergo genome wide DNA demethylation, histone modification, imprinting erasure and reactivation of X chromosome [14][15]. Until this stage, sex specification is not achieved as PGCs are bipotent in nature and express common germ cell markers shared by both male and female gametes. In humans, PGC specification begins at around the third gestational week during which erasure of DNA methylation takes place [16][17][18][19]. Differences have been observed among transcription factors requirement for PGCs specification and maintenance among humans and mice [20][21][22]. During progressive stages, cells undergo a transitional shift from pluripotency associated marker expression to germ cell specificity in a gradual manner and attain expression of early PGC makers, late PGC markers, meiosis markers, undergo epigenetic reprograming and finally attains mature germ cell markers [17]. While analyzing the transcriptome of human PGCs, Guo and colleagues [17] found that both pluripotency and germ cell specific genes are simultaneously expressed from their migrating to the gonadal stage. Diagrammatical outflow of the PGC development and specification has been given in Figure 1.
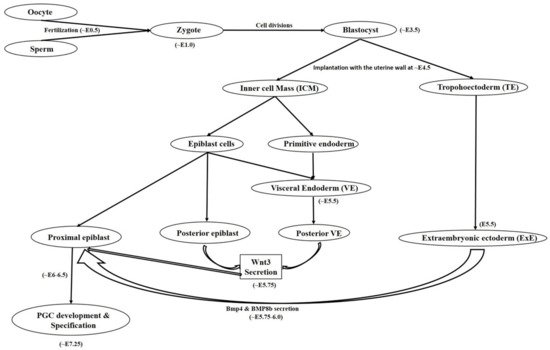
Figure 1. Diagrammatical outflow of the PGC development and specification in mice. Oocyte and sperm after getting fertilized produces zygote. While undergoing further cell divisions, zygote attain blastocyst stage at around embryonic development day 3.5 (~E3.5) having an outer trophoblast cell monolayer and pluripotent inner cell mass (ICM) which is further separated into epiblast and primitive endoderm. Implantation of the blastocyst with the uterine wall takes place at ~E4.5. Further developmental stages are comprised of extraembryonic ectoderm (ExE) development from trophoectoderm and visceral endoderm (VE) from primitive endoderm. Furthermore, Wnt3 is secreted by both of the posterior epiblast and VE and BMP4 & BMP8b secreted by ExE. Wnt3 and BMPs secretions finally assists in production of a few primordial germ cells (PGCs) from proximal epiblast and finally PGC specification takes place.
1.2. Primordial Germ Cell Migration
Once the PGCs are formed, their migration to gonadal ridges constitutes an important developmental step towards gamete specification and their proper functioning. It is worth noticing that the concept of signaling guidance for PGCs towards the gonads was first of all described in zebrafish [23]. Both intrinsic motility factors and external guidance cues are required by the PGCs to successfully migrate and maintain their survivability [24]. In the beginning, BMP signaling from ExE governs PGC formation from epiblast cells, which are further mediated by BLIMP1 or PRDM1 transcriptional regulators which not only stimulate PGC specific gene such as Stella but also suppresses the expression of somatic cell genes cascade [25][26]. Like BLIMP1, PRDM14 also regulates mouse PGC specification, lacking or knockdown of both the factors either resulted into improper PGC differentiation and defected migration or induced sterility in mice models [27][28]. Along with BLIMP1 and PRDM14, AP2γ has also been found essential in regulating mammalian PGC specification [29]. AP2γ is expressed by mouse PGCs from E7.25 to E12.5 and its targeted disruption has resulted into male and female sterility. It was also believed that PGC migration in mouse is also coordinated by interferon induced transmembrane protein 1 (IFITM1), as RNA interference induced IFITM1 knockdown in the primitive streak resulted into PGC migration failure towards endoderm, indicating possible role of IFITM1 in PGC migration from mesoderm to endoderm [30]. However, contradicting results were shown by Lange UC et al. (2008), where embryonal deletion of the IFITM1 gene family could not cause impaired PGC migration [31]. During the period of migration and arrival of PGCs towards hindgut, Nanos3 plays multiple vital roles by maintaining germ cell’s undifferentiated state, protecting PGCs against apoptosis by suppressing Bax-dependent and independent pathways and thereby helps in germ cell development [32]. Nanos3 is expressed by migrating PGCs at various germ cell specific developmental stages in both testis and ovaries. Moreover, without its expression, germ cell detection in the ovary and testis is not possible [13][32]. Failure in migration can cause developmental issues and infertility, and also may cause teratoma formation in extra gonadal regions [10]. At around E11.5, most PGCs are migrated towards genital ridges and become populated there at around E12.5. During later stages, establishment of sexually dimorphic sex cords takes place between E13.5–17.5 and birth and most of the ectopic PGCs die due to apoptotic pathways and rest of them transforms into male and female gametes in their respective sex gonads [13]. Researchers believed that PGCs lose their migration properties once they associate with somatic cells and reach gonads [24]. One possible reason for their ceasing behavior may be due to the cell-to-cell interactions between PGCs and somatic cells. Proper mechanism behind this stoppage is not known and need in depth elucidation. Diagrammatical outflow of the PGC migration has been given in Figure 2.
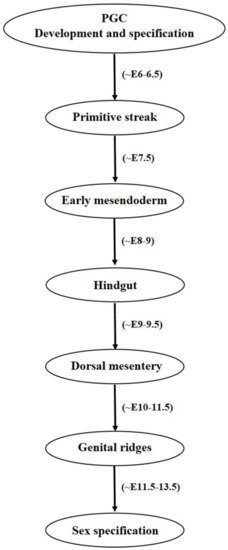
Figure 2. Diagrammatical outflow of the PGC migration. PGCs undergo a series of events before reaching gonadal ridges and finally sex specification. They are identified as a few precursor cells at the base of allantois and specified under the effect of molecular cues associated with Wnt3 and BMP pathways. After passing through a number of stages, PGCs finally enter genital ridges and specified as male and female gametes.
2. Functional Efficacy of the Female Germ Cells
Ability to generate offspring by either getting fertilized with healthy spermatocyte or parthenogenetically has been the prime function of the oocytes. Unlike male gametes, female germ cells (oocytes) have been thought to have limited number which decline with the age and there has been contradictory views regarding regenerative activity in juvenile or adult ovaries. Pathological conditions such as cancer or ovarian disorders including premature ovarian failure (POF) and premature ovarian insufficiency (POI) put additional threats to female reproductive life and ultimately results into infertility. Therefore, researchers throughout the globe have consistently made extensive efforts to develop methodologies so that functional efficacy of the in vitro derived oocytes can be enhanced. Although, due to complexities in deriving oocyte-like cells (OLCs) in vitro (ranging from ethical issues concerned with use of pluripotent ESCs or iPSCs, low response by less potent MSCs, or xenogeneic concerns regarding use of follicular fluid or specialized cells from different organisms), not much success has been achieved. Only a few researchers have reported derivations of healthy offspring by using ovary-derived cells, i.e., premeiotic fetal germ cells, germline stem cells, primordial germ cells, granulosa cells, and ovarian stem cells especially in nonhuman species (murine) [33][34][35][36][37]. However, getting the in vitro derived fertilizable oocytes and finally healthy progeny is still an unachieved distant milestone. Similarly, in vitro differentiation into fertilizable OLCs using adult MSC sources other than ovary derived cells is still far away. Therefore, many efforts will be needed to achieve this goal.
3. Developmental Impairments and Associated Abnormalities
A number of factors are responsible for gametes specific developmental impairments and associated in utero or postnatal abnormalities. Proper development and functionality of a gene and also efficiency of the traits under control are dependent on the governing (regulating) signaling pathways, autocrine and paracrine factors, surrounding environmental cues and nourishing elements (in the form of cytokines and growth factors). Shortcomings in any of the above said necessities may cause partial or permanent defects. Diagrammatical representation of factors affecting germ cell development and their functionality has been depicted in Figure 3. These can be categorized as follows:
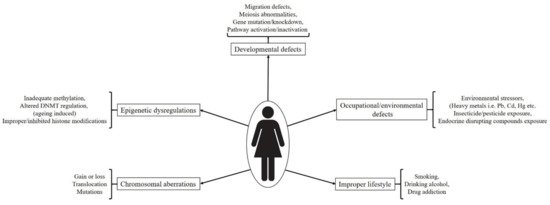
Figure 3. Diagrammatical representation of factors affecting germ cell development and their functionality. All germ cell developmental stages are affected by a number of factors including mutations, chromosomal abnormalities, improper lifestyles, epigenetic dysregulations, environmental toxicants etc. Which ultimately leads to functional impairment followed by disease development. Abbreviation- Pb: Lead; Cd: Cadmium; Hg: Mercury; DNMT: DNA methyltransferase).
3.1. Chromosomal Aberrations
Chromosomal aberrations correspond to insertion, deletion or translocation of some or the whole part (arms) of a chromosome. These abnormalities have been associated with the occurrence of impaired functioning of the various body parts and hence are the causatives of many complications including ovarian germ cell tumors (OGSTs). According to Matyakhina et al., all malignant human tumors occur due to chromosomal aberrations induced chromosomal instability [38]. These disturbances happen due to abnormal segregation of chromosomes at the time of cell division. While observing abnormal karyotypes from the patients with OGSTs, comparative genomic hybridization (CGH) and fluorescence in situ hybridization (FISH) techniques revealed that gains or loss from the chromosomal arms or some of the whole chromosomes resulted into dysgerminomas (DGs), endodermal sinus tumors (ESTs) and immature teratomas [39][40][41]. The gain and loss of different chromosome numbers such as loss of 1p, gain of 1q or whole chromosomal gains chromosome numbers including 3, 8, 14 and 21 have been observed in female patients with malignant OGCTs. Among OGCTs, dysgerminomas were associated with abnormalities in 12p chromosome [39]. While performing dual color fluorescence in situ hybridization (FISH), Paolo Cossu-Rocca and colleagues observed that aberrations in the 12p chromosome were associated with 81% of total dysgerminomas [41] and seminomas (testicular germ cell tumor). Chromosomal abnormalities have also been correlated with ovarian granulosa cell tumors [42][43]. Comparative genomic hybridization and FISH techniques revealed that gains of chromosomes 12 and 14 and losses of chromosomes 22 and X were causatives of ovarian granulosa cell tumors [42]. In a similar study, patients with stage 1 granulosa cell tumors were shown to have chromosomal abnormalities in the form of losses from 1p33 to p36, 16p13.1, 16q and 22q chromosomes whereas gains were observed in chromosomes including 7p15 to p21, 12 and 14 [43]. Among these chromosomes, losses from chromosome 22 and gains in chromosome 14 were more prevalent. However, none of the conditions exhibited alarming degree of metastasis. In general, due to low prevalence rate (comparatively less than testicular germ cell tumors) in comparison to other extra-gonadal tumors, not many reports are available. However, it is equally important to draw attention on the role of chromosomal aberrations in the occurrence of diseases especially OGCTs as they can be diagnosed and can be proved helpful in avoiding future complications. Therefore, techniques like karyotyping, CGH and FISH should also be given preferences so that anomalies against normal behavior can be diagnosed and eradicated in time.
3.2. Spontaneous Genetic and Epigenetic Regulations
Epigenetics plays an important role in regulating overall transformation and functional activity of germ cells which upon fertilization can transmit proper information to the next generation [44][45]. A series of gradual events including PGC specification, meiotic entry, epigenetic imprinting of PGCs, histone modification and X-chromosome inactivation, DNA methylation erasure and its resumption during gamete formation, are highly important during gametogenesis [46]. The actual reason behind epigenetic information erasure in germ cells is not known but it may be attributed to the restricted transmission of unwanted acquired epimutations during life [18]. Although, except ESCs and iPSCs (only few reports), other stem cell sources derived gamete-like cells could either failed to initiate meiosis, or failed to prove their functionality by producing healthy viable offsprings upon fertilization. This may be perhaps due to developmental issues related to improper epigenetic modifications [46]. During migration period, PGCs go through genome wide DNA demethylation, histone modification by means of downregulation of H3 lysine 9 dimethylation (H3K9me2), upregulation of H3 lysine 27 trimethylation (H3K27me3), imprinting erasure and reactivation of X chromosome [14][15]. By the end of E12.5, H3K9me3 and H3K64me3 are gradually lost from PGCs and all these events are vital for efficient differentiation into gametes [47][48].
3.3. External/Environmental Factors and Incorrect Lifestyle Associated Defects/Exposure to Toxicants In Utero
A healthy life with proper functionality of all the organs depends a lot on many factors including surrounding environment, lifestyle, routine activities (exercising, food and drinking habits), and opted occupation. All these factors have also been associated with the reproductive life of an individual especially during female pregnancies. Here below is the detailed description about these factors in relation to female reproductive health.
3.3.1. Chemical Intoxicants
Chemical intoxication especially during cancer treatments have been associated with reproductive disorders in both males and females. Fetal exposure to cyclophosphamide was shown to induce testicular germ cell tumor (TGCT), gonadal toxicity and reduced spermatogenesis [49]. Offspring produced from cyclophosphamide exposed mice showed high incidence of TGCT and reduced testicular and epididymal sperm count whereas primordial follicle loss and increased follicular growth activation was observed in female offspring. Endocrine disrupting compounds (EDCs) represents another category of reproductive toxicants which produces deleterious effects on male and female gametes by interfering natural hormone activities and other associated developmental processes [50]. Women (at different reproductive life stages) exposed to environmental di(2-ethylhexyl) phthalate (DEHP) and its metabolites were shown to have impaired reproduction and reduced fertility. Moreover, presence of DEHP and its major metabolite mono(2-ethyhexyl) phthalate (MEHP) in the urine samples of females have been associated with reduced number of high-quality fertilizable oocytes, and even early pregnancy loss [51][52][53]. In a recent study, Jing-Cai Liu et al. demonstrated that increased PDE3A expression in oocytes as a result of DEHP exposure results into impaired follicle assembly [54]. It was further evidenced that decreased levels of cAMP and PKA in oocytes resulted in an increase in PDE3A and ultimately caused impaired follicle assembly in DEHP-exposed ovaries. These compounds have also been demonstrated to cause deleterious reproductive effects, impaired folliculogenesis and ovulation, and decreased pregnancy rates in non-human species including rats, mice and cows [55][56][57]. Therefore, exposure to such deleterious and toxic chemicals must be avoided and in case of cancer treatment it is better to preserve the functional gametes so that fertility can be restored later on by transplanting back the autologous gametes into the donor body. Use of stem cells based therapeutic agents (stem cell therapy) or very small embryonic cell-like cells (which can survive cancer therapies) can be another efficient and safer way to restore fertility [58].
3.3.2. Smoking, Alcoholism and Drug Abuse
Incorrect lifestyle, i.e., smoking, alcohol intake, and illicit drug consumption, also causes serious detrimental effects on reproductive lives of both males and females. The harmful effects of smoking on female reproductive system specifically impaired ovarian function were evidenced by decreased estradiol and progesterone production by in vitro cultured human luteinized granulosa cells under direct environmental tobacco smoke exposure [59]. A concentration (environmental tobacco smoke exposure) dependent increase in the expression of CYP1B1 both at gene and protein levels was seen in the cultured granulosa cells. High activity polymorphism of CYP1B1 in cigarette smokers was also correlated with high risk of ovarian cancer development with disturbed hormonal regulation [60]. While analyzing retrieved ovarian cells from women undergoing assisted reproductive technologies (ART), major ovotoxic action of smoking was found to be oxidative stress and DNA damage in granulosa and oocyte cumulus cells, causing impaired functionality including improper oocyte maturation, gonadotropins binding to their receptors, increased zona pellucida thickness and reduced fertilization capacity [61][62][63][64]. In a cigarette smoke induced chronic obstructive pulmonary disease mouse model study, direct nasal exposure resulted into ovotoxicity by means of higher levels of primordial follicle depletion, apoptosis in antral follicle oocytes, induced oxidative stress and reduced availability of follicles for ovulation [65]. Similar ill effects were observed in other murine model studies targeting cigarette smoke induced ovarian impairment [66][67][68][69]. The effect of alcohol consumption on the female reproductive system and fertility has shown many contradictory results. In a study involving 124 patients undergoing routine urine inspection regarding steroidal hormone measurement, women with alcoholic habits during menstruation were shown to have >50% reduced conception probability during menstrual cycles [70]. Caffeine consumption further enhanced alcohol induced negative effects whereas no adverse effect was seen in participants consuming caffeine only. It has been reported that high consumption rate (rather than moderate alcohol consumption) results into decreased fertility [71]. However, a number of cohort case studies have exhibited no correlation between alcohol consumption and female infertility [72][73][74]. Illicit drug use has also been associated with the female reproductive system related issues. Previously, it was demonstrated that women with regular marijuana intake have increased risk for ovulatory dysfunction [75]. Another study concerning women with occasional or frequent marijuana intake habits, showed longer follicular phase, delayed ovulation, and slightly increased rate of ovulatory inhibition in comparison to non-marijuana users [76]. Contrastingly, in a recent report it was found that women with habits of co-use of marijuana with tobacco were shown to have shorter luteal phase than only tobacco users [77] however no differences between follicular phase and menstrual cycle lengths were observed among both types of participants. Missing correlating link between these studies corresponds to a number of factors including heterogeneity in the study design, volume or amount of cigarette/alcohol/illicit drug consumption on occasional or regular basis, exact time or duration of consumption (such as early age, during pregnancy or later stages), past intake history with no current consumption etc. Therefore, it’s hard to conclude whether such incorrect habits or life styles plays a direct or definite role in deteriorating reproductive life to a greater extent or not. However, as all of these lifestyles poses negative effects on the health therefore must be avoided much longer before conceiving or attaining pregnancy. Factors causing defects in germ cell development and fertility impairment has been depicted in Figure 3.
References
- Hayashi, M.; Kawaguchi, T.; Durcova-Hills, G.; Imai, H. Generation of germ cells from pluripotent stem cells in mammals. Reprod. Med. Biol. 2018, 17, 107–114.
- Felici, M.D. Germ stem cells in the mammalian adult ovary: Considerations by a fan of the primordial germ cells. Mol. Hum. Reprod. 2010, 16, 632–636.
- Aramaki, S.; Hayashi, K.; Kurimoto, K.; Ohta, H.; Yabuta, Y.; Iwanari, H.; Mochizuki, Y.; Hamakubo, T.; Kato, Y.; Shirahige, K.; et al. A mesodermal factor, T, specifies mouse germ cell fate by directly activating germline determinants. Dev. Cell 2013, 27, 516–529.
- Ying, Y.; Qi, X.; Zhao, G.Q. Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 7858–7862.
- Ying, Y.; Zhao, G.Q. Cooperation of Endoderm-Derived BMP2 and Extraembryonic Ectoderm-Derived BMP4 in Primordial Germ Cell Generation in the Mouse. Dev. Biol. 2001, 232, 484–492.
- Lawson, K.A.; Dunn, N.R.; Roelen, B.A.; Zeinstra, L.M.; Davis, A.M.; Wright, C.V.; Korving, J.P.; Hogan, B.L.M. Bmp4 is required for the generation of primordial germ cells in the mouse embryos. Genes. Dev. 1999, 13, 424–436.
- Tremblay, K.D.; Dunn, N.R.; Robertson, E.J. Mouse embryos lacking Smad1 signals display defects in extra-embryonic tissues and germ cell formation. Development 2001, 128, 3609–3621.
- Chu, G.C.; Dunn, N.R.; Anderson, D.C.; Oxburgh, L.; Robertson, E.J. Differential requirements for Smad4 in TGFβ-dependent patterning of the early mouse embryo. Development 2004, 131, 3501–3512.
- Chang, H.; Matzuk, M.M. Smad5 is required for mouse primordial germ cell development. Mech. Dev. 2001, 104, 61–67.
- Cantú, A.V.; Laird, D.J. Primordial germ cell migration and the Wnt signaling pathway. Anim. Reprod. 2017, 14, 89–101.
- Saitou, M.; Payer, B.; Lange, U.C.; Erhardt, S.; Barton, S.C.; Surani, M.A. Specification of germ cell fate in mice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 1363–1370.
- Lange, U.C.; Saitou, M.; Western, P.S.; Barton, S.C.; Surani, M.A. The Fragilis interferon-inducible gene family of transmembrane proteins is associated with germ cell specification in mice. BMC. Dev. Biol. 2003, 3, 1–11.
- Tres, L.L.; Rosselot, C.; Kierszenbaum, A.L. Primordial Germ Cells What Does It Take to be alive? Mol. Reprod. Dev. 2004, 68, 1–4.
- Saitou, M.; Kagiwada, S.; Kurimoto, K. Epigenetic reprogramming in mouse pre-implantation development and primordial germ cells. Development 2012, 139, 15–31.
- Lee, H.J.; Hore, T.A.; Reik, W. Reprogramming the Methylome: Erasing Memory and Creating Diversity. Cell Stem Cell 2014, 14, 710–719.
- Gkountela, S.; Zhang, K.X.; Shafiq, T.A.; Liao, W.W.; Calvopiña, J.H.; Chen, P.Y.; Clark, A.T. DNA Demethylation Dynamics in the Human Prenatal Germline. Cell 2015, 161, 1425–1436.
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452.
- Canovas, S.; Campos, R.; Aguilar, E.; Cibelli, J.B. Progress towards human primordial germ cell specification in vitro. Mol. Hum. Reprod. 2017, 23, 4–15.
- Chen, D.; Sun, N.; Hou, L.; Kim, R.; Faith, J.; Aslanyan, M.; Tao, Y.; Zheng, Y.; Fu, J.; Liu, W.; et al. Human Primordial Germ Cells Are Specified from Lineage-Primed Progenitors. Cell Rep. 2019, 29, 4568–4582.e5.
- Chen, D.; Liu, W.; Lukianchikov, A.; Hancock, G.V.; Zimmerman, J.; Lowe, M.G.; Kim, R.; Galic, Z.; Irie, N.; Surani, M.A.; et al. Germline competency of human embryonic stem cells depends on eomesodermin. Biol. Reprod. 2017, 97, 850–861.
- Irie, N.; Weinberger, L.; Tang, W.W.C.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268.
- Sasaki, K.; Yokobayashi, S.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Kurimoto, K.; Ohta, H.; Moritoki, Y.; Iwatani, C.; Tsuchiya, H.; et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell 2015, 17, 178–194.
- Raz, E. Primordial germ-cell development: The zebrafish perspective. Nat. Rev. Genet. 2003, 4, 690–700.
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49.
- Saitou, M.; Payer, B.; O’Carroll, D.; Ohinata, Y.; Surani, M.A. Blimp1 and the emergence of the germ line during development in the mouse. Cell Cycle 2005, 4, 1736–1740.
- Vincent, S.D.; Dunn, N.R.; Sciammas, R.; Sharpiro-Shalef, M.; Davis, M.M.; Calame, K.; Bikoff, E.K.; Robertson, E.J. The zinc finger transcriptional repressor Blimp1/Prdm1 is dispensable for early axis formation but is required for specification of primordial germ cells in the mouse. Development 2005, 132, 1315–1325.
- Kurimoto, K.; Yamaji, M.; Seki, Y.; Saitou, M. Specification of the germ cell lineage in mice: A process orchestrated by the PR-domain proteins, Blimp1 and Prdm14. Cell Cycle 2008, 7, 3514–3518.
- Yamaji, M.; Seki, Y.; Kurimoto, K.; Yabuta, Y.; Yuasa, M.; Shigeta, M.; Yamanaka, K.; Ohinata, Y.; Saitou, M. Critical function of Prdm14 for the establishment of the germ cell lineage in mice. Nat. Genet. 2008, 40, 1016–1022.
- Tanaka, S.S.; Yamaguchi, Y.L.; Tsoi, B.; Lickert, H.; Tam, P.P. IFITM/Mil/fragilis family proteins IFITM1 and IFITM3 play distinct roles in mouse primordial germ cell homing and repulsion. Dev. Cell 2005, 9, 745–756.
- Lange, U.C.; Adams, D.J.; Lee, C.; Barton, S.; Schneider, R.; Bradley, A.; Surani, M.A. Normal germ line establishment in mice carrying a deletion of the Ifitm/Fragilis gene family cluster. Mol. Cell Biol. 2008, 28, 4688–4696.
- Weber, S.; Eckert, D.; Nettersheim, D.; Gillis, A.J.; Schafer, S.; Kuckenberg, P.; Ehlermann, J.; Werling, U.; Biermann, K.; Looijenga, L.H.; et al. Critical function of AP-2g/TCFAP2C in mouse embryonic germ cell maintenance. Biol. Reprod. 2010, 82, 214–223.
- Julaton, V.T.A.; Pera, R.A.R. NANOS3 function in human germ cell development. Hum. Mol. Genet. 2011, 20, 2238–2250.
- Shen, W.; Zhang, D.; Qing, T.; Cheng, J.; Bai, Z.; Shi, Y.; Ding, M.; Deng, H. Live Offspring Produced by Mouse Oocytes Derived from Premeiotic Fetal Germ Cells. Biol. Reprod. 2006, 75, 615–623.
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636.
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from Oocytes Derived from in Vitro Primordial Germ Cell–like Cells in Mice. Science 2012, 338, 971–975.
- Morohaku, K.; Tanimoto, R.; Sasaki, K.; Kawahara-Miki, R.; Kono, T.; Hayashi, K.; Hiraoe, Y.; Obata, Y. Complete in vitro generation of fertile oocytes from mouse primordial germ cells. Proc. Natl. Acad. Sci. USA 2016, 113, 9021–9026.
- Tian, C.; Liu, L.; Ye, X.; Fu, H.; Sheng, X.; Wang, L.; Wang, H.; Heng, D.; Liu, L. Functional Oocytes Derived from Granulosa Cells. Cell Rep. 2019, 29, 4256–4267.
- Matyakhina, L.; Lenherr, S.M.; Stratakis, C.A. Protein Kinase A and Chromosomal Stability. Ann. N. Y. Acad. Sci. 2002, 968, 148–157.
- Kraggerud, S.M.; Szymanska, J.; Abeler, V.M.; Kaern, J.; Eknaes, M.; Heim, S.; Teixeira, M.R.; Trope, C.G.; Peltomäki, P.; Lothe, R.A. DNA Copy Number Changes in Malignant Ovarian Germ Cell Tumors. Cancer Res. 2000, 60, 3025–3030.
- Bussey, K.J.; Lawce, H.J.; Himoe, E.; Shu, X.O.; Suijkerbuijk, R.F.; Olson, S.B.; Magenis, R.E. Chromosomes 1 and 12 abnormalities in pediatric germ cell tumors by interphase fluorescence in situ hybridization. Cancer Genet. Cytogenet. 2001, 125, 112–118.
- Cossu-Rocca, P.; Zhang, S.; Roth, L.M.; Eble, J.N.; Zheng, W.; Abdul Karim, F.W.; Michael, H.; Emerson, R.E.; Jones, T.D.; Hattab, E.M.; et al. Chromosome 12p abnormalities in dysgerminoma of the ovary: A FISH analysis. Mod. Pathol. 2006, 19, 611–615.
- Mayr, D.; Kaltz-Wittmer, C.; Arbogast, S.; Amann, G.; Aust, D.E.; Diebold, J. Characteristic Pattern of Genetic Aberrations in Ovarian Granulosa Cell Tumors. Mod. Pathol. 2002, 15, 951–957.
- Lin, Y.S.; Eng, H.L.; Jan, Y.J.; Lee, H.S.; Ho, W.L.; Liou, C.P.; Lee, W.Y.; Tzeng, C.C. Molecular cytogenetics of ovarian granulosa cell tumors by comparative genomic hybridization. Gynecol. Oncol. 2005, 97, 68–73.
- Feng, L.; Chen, X. Epigenetic Regulation of Germ Cells— Remember or Forget? Curr. Opin. Genet. Dev. 2015, 31, 20–27.
- Cinalli, R.M.; Rangan, P.; Lehmann, R. Germ cells are forever. Cell 2008, 132, 559–562.
- Sun, Y.C.; Wang, Y.Y.; Ge, W.; Cheng, S.F.; Dyce, P.W.; Shen, W. Epigenetic regulation during the differentiation of stem cells to germ cells. Oncotarget 2017, 8, 57836–57844.
- Hajkova, P.; Ancelin, K.; Waldmann, T.; Lacoste, N.; Lange, U.C.; Cesari, F.; Lee, C.; Almouzni, G.; Schneider, R.; Surani, M.A. Chromatin dynamics during epigenetic reprogramming in the mouse germ line. Nature 2008, 452, 877–881.
- Daujat, S.; Weiss, T.; Mohn, F.; Lange, U.C.; Ziegler-Birling, C.; Zeissler, U.; Lappe, M.; Schübeler, D.; Torres-Padilla, M.E.; Schneider, R. H3K64 trimethylation marks heterochromatin and is dynamically remodeled during developmental reprogramming. Nat. Struct. Mol. Biol. 2009, 16, 777–781.
- Paul, B.C.; Ana, L.D.; Hazel, L.K.; Richard, A.A.; Angabin, M.; Marvin, L.M.; Gunapala, S. Fetal Cyclophosphamide Exposure Induces Testicular Cancer and Reduced Spermatogenesis and Ovarian Follicle Numbers in Mice. PLoS ONE 2014, 9, e93311.
- Zvi, R.; Alisa, K.E.; Dorit, K. Effect of environmental contamination on female and male gametes—A lesson from bovines. Anim. Reprod. 2020, 17, e20200041.
- Toft, G.; Jönsson, B.A.G.; Lindh, C.H.; Jensen, T.K.; Hjollund, N.H.; Vested, A.; Bonde, J.P. Association between Pregnancy Loss and Urinary Phthalate Levels around the Time of Conception. Environ. Health Perspect. 2012, 120, 458–463.
- Tranfo, G.; Caporossi, L.; Paci, E.; Aragona, C.; Romanzi, D.; De Carolis, C.; De Rosa, M.; Capanna, S.; Papaleo, B.; Pera, A. Urinary phthalate monoesters concentration in couples with infertility problems. Toxicol. Lett. 2012, 213, 15–20.
- Machtinger, R.; Gaskins, A.J.; Racowsky, C.; Mansur, A.; Adir, M.; Baccarelli, A.A.; Calafat, A.M.; Hauser, R. Urinary concentrations of biomarkers of phthalates and phthalate alternatives and IVF outcomes. Environ. Int. 2018, 111, 23–31.
- Liu, J.C.; Yan, Z.H.; Li, B.; Yan, H.C.; Felici, M.D.; Shen, D. Di (2-ethylhexyl) phthalate impairs primordial follicle assembly by increasing PDE3A expression in oocytes. Environ. Pollut. 2020, 116088, online ahead of print.
- Gupta, R.K.; Singh, J.M.; Leslie, T.C.; Meachum, S.; Flaws, J.A.; Yao, H.H. C-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol. Appl. Pharmacol. 2010, 242, 224–230.
- Hannon, P.R.; Niermann, S.; Flaws, J.A. Acute exposure to Di(2-Ethylhexyl) phthalate in adulthood causes adverse reproductive outcomes later in life and accelerates reproductive aging in female mice. Toxicol. Sci. 2016, 150, 97–108.
- Kalo, D.; Roth, Z. Low level of mono(2-ethylhexyl) phthalate reduces oocyte developmental competence in association with impaired gene expression. Toxicology 2017, 377, 38–48.
- Bharti, D.; Jang, S.J.; Lee, S.Y.; Lee, S.L.; Rho, G.J. In Vitro Generation of Oocyte Like Cells and Their In Vivo Efficacy: How Far We have been Succeeded. Cells 2020, 9, 557.
- Justin, D.V.; Catherine, A.V.; Craig, B.M.; Nicolas, R.L.; Alan, J.C. In vitro exposure to environmental tobacco smoke induces CYP1B1 expression in human luteinized granulosa cells. Reprod. Toxicol. 2006, 22, 731–737.
- Goodman, M.T.; McDuffie, K.; Kolonel, L.N.; Terada, K.; Donlon, T.A.; Wilkens, L.R.; Guo, C.; Marchand, L.L. Case-control study of ovarian cancer and polymorphisms in genes involved in catecholestrogen formation and metabolism. Cancer Epidemiol. Biomark. Prev. 2001, 10, 209–216.
- Alviggi, C.; Cariati, F.; Conforti, A.; De Rosa, P.; Vallone, R.S.I.; Pivonello, R.; De Placido, G. The effect of FT500 Plus® on ovarian stimulation in PCOS women. Reprod. Toxicol. 2016, 59, 40–44.
- Budani, M.C.; Carletti, E.; Tiboni, G.M. Cigarette smoke is associated with altered expression of antioxidant enzymes in granulosa cells from women undergoing in vitro fertilization. Zygote 2017, 25, 296–303.
- Sinko, I.; Morocz, M.; Zadori, J.; Kokavszky, K.; Rasko, I. Effect of cigarette smoking on DNA damage of human cumulus cells analyzed by comet assay. Reprod. Toxicol. 2005, 20, 65–71.
- Shiloh, H.; Lahav-Baratz, S.; Koifman, M.; Ishai, D.; Bidder, D.; Weiner-Meganzi, Z.; Dirnfeld, M. The impact of cigarette smoking on zona pellucida thickness of oocytes and embryos prior to transfer into the uterine cavity. Hum. Reprod. 2004, 19, 157–159.
- Sobinoff, A.P.; Beckett, E.L.; Jarnicki, A.G.; Sutherland, J.M.; McCluskey, A.; Hansbro, P.M.; McLaughlin, E.A. Scrambled and fried: Cigarette smoke exposure causes antral follicle destruction and oocyte dysfunction through oxidative stress. Toxicol. Appl. Pharmacol. 2013, 271, 156–167.
- Jennings, P.C.; Merriman, J.A.; Beckett, E.L.; Hansbro, P.M.; Jones, K.T. Increased zona pellucida thickness and meiotic spindle disruption in oocytes from cigarette smoking mice. Hum. Reprod. 2011, 26, 878–884.
- Paixao, L.L.; Gaspar-Reis, R.P.; Gonzalez, G.P.; Santos, A.S.; Santana, A.C.; Santos, R.M.; Spritzer, P.M.; Nascimento-Saba, C.C. Cigarette smoke impairs granulosa cell proliferation and oocyte growth after exposure cessation in young Swiss mice: An experimental study. J. Ovarian Res. 2012, 5, 25.
- Mai, Z.; Lei, M.; Yu, B.; Du, H.; Liu, J. The effects of cigarette smoke extract on ovulation, oocyte morphology and ovarian gene expression in mice. PLoS ONE 2014, 9, e95945.
- Lee, H.M.; Kim, C.W.; Hwang, K.A.; Sung, J.H.; Lee, J.K.; Choi, K.C. Cigarette smoke impaired maturation of ovarian follicles and normal growth of uterus inner wall of female wild-type and hypertensive rats. Reprod. Toxicol. 2017, 73, 232–240.
- Hakim, R.B.; Gray, R.H.; Zakur, H. Alcohol and caffeine consumption and decreased fertility. Fertil. Steril. 1998, 70, 632–637.
- Eggert, J.; Theobald, H.; Engfeldt, H. Effects of alcohol consumption on female fertility during an 18-year period. Fertil. Steril. 2004, 81, 379–383.
- Juhl, M.; Andersen, A.M.N.; Gronbaek, M.; Olsen, J. Moderate alcohol consumption and waiting time to pregnancy. Hum. Reprod. 2001, 16, 2705–2709.
- Tolstrup, J.S.; Kjaer, S.K.; Holst, C.; Sharif, H.; Munk, C.; Osler, M.; Schmidt, L.; Andersen, A.M.N.; Gronbaek, M. Alcohol use as predictor for infertility in representative population of Danish women. Acta Obstet. Gynecol. Scand. 2003, 82, 744–749.
- Chavarro, J.E.; Rich-Edwards, J.W.; Rosner, B.A.; Willett, W.C. Caffeinated and Alcoholic Beverage Intake in Relation to Ovulatory Disorder Infertility. Epidemiology 2009, 20, 374–381.
- Mueller, B.A.; Daling, J.R.; Weiss, N.S.; Moore, D.E. Recreational drug use and the risk of primary infertility. Epidemiology 1990, 1, 195–200.
- Jukic, A.M.Z.; Weinberg, C.R.; Baird, D.D.; Wilcox, A.J. Lifestyle and reproductive factors associated with follicular phase length. J. Womens Health (Larchmt) 2007, 16, 1340–1347.
- Lammert, S.; Harrison, K.; Tosun, N.; Allen, S. Menstrual Cycle in Women Who Co-use Marijuana and Tobacco. J. Addict. Med. 2018, 12, 207–211.

