 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hans-Peter Fiedler | + 1422 word(s) | 1422 | 2021-05-31 12:20:41 | | | |
| 2 | Karina Chen | Meta information modification | 1422 | 2021-06-04 10:37:26 | | |
Video Upload Options
Abyssomicins represent a new family of polycyclic macrolactones. The first described compounds of the abyssomicin family were abyssomicin B, C, atrop-C, and D, produced by the marine actinomycete strain Verrucosispora maris AB-18-032, which was isolated from a sediment collected in the Sea of Japan. Among the described abyssomicins, only abyssomicin C and atrop-abyssomicin C show a high antibiotic activity against Gram-positive bacteria, including multi-resistant and vancomycin-resistant strains. The inhibitory activity is caused by a selective inhibition of the enzyme 4-amino-4-deoxychorismate synthase, which catalyzes the transformation of chorismate to para-aminobenzoic acid, an intermediate in the folic acid pathway.
1. Introduction
An efficient search for new bioactive compounds, such as antibiotics, demands specific requirements:
First, there is a need for a massive set of taxonomically characterized and dereplicated microorganisms isolated from terrestrial and/or marine habitats, with a high potency of producing unique secondary metabolites. A representative example of such a source are members of the order Actinomycetales, which are known as potent producers of unique secondary metabolites. Second, there needs to be a specific target that is essential for a pathogenic organism but is not present in humans in order to provide selectivity in toxic effects. Third, highly efficient analytic equipment and techniques are required to identify novel secondary metabolites in the culture broth or extracts from mycelium and culture filtrate, such as the combination of HPLC with diode array detection and/or mass spectrometry and characterization employing a home-made or commercial database. One of our screening programs was focused on the inhibition of the chorismate pathway, taking over an idea of Emeritus Professor Hans Zähner, one of the pioneers in antibiotic research and chair of the Institute of Microbiology for over 30 years at the University of Tübingen, who retired in 1994. The chorismate pathway leads to the biosynthesis of the aromatic amino acids, and to the biosynthesis of para-aminobenzoic acid (pAba), an intermediate in the biosynthesis of folic acid. This pathway is present in plants, fungi, prokaryotes, and some parasites, such as Plasmodium and Toxoplasma, but it is not present in humans (Figure 1). The biosynthesis of pAba is catalyzed by two enzymes, 4-amino-4-deoxychorismic acid (ADC) synthase and ADC lyase, which were the target of our screening concept.
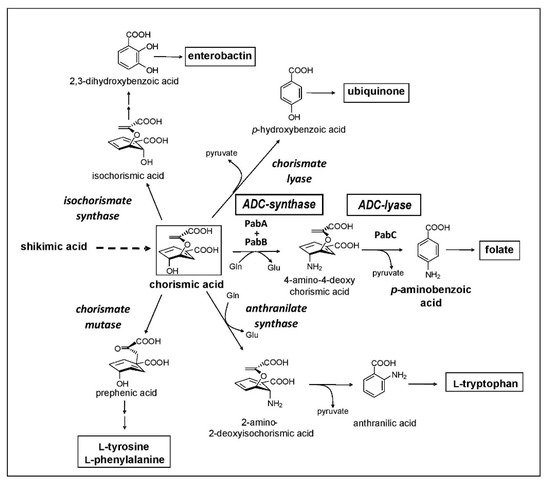
Figure 1. The bacterial chorismate biosynthetic pathway.
As promising candidates for the production of new secondary metabolites, we selected 201 characterized strains from our own collection and strains from my collaborators Alan T. Bull (University of Kent) and Michael Goodfellow (Newcastle University) belonging to the families Streptomycetaceae and Micromonosporaceae, and rare actinomycete genera, isolated both from terrestrial and marine habitats. Extracts were subjected to the screening assay based on a simple but highly selective agar plate diffusion assay. Only one extract obtained from strain AB-18-032, a marine isolate from a sediment collected from the Sea of Japan belonging to the family Micromonosporaceae, inhibited pAba biosynthesis selectively. The secondary metabolite pattern of the active extract was analyzed by HPLC-diode array, whereby three metabolites were characterized by our HPLC-UV-Vis Database as presumably new compounds, which we named abyssomicins.
The first announcement of abyssomicins was done two years later in 2003 in a patent application [1] because of their pharmacologically relevant properties which favored their application as promising therapeutics against infection diseases, especially of pathogenic multi-resistant Gram-positive bacteria, including vancomycin-resistant Staphylococcus aureus. Subsequently, we published the screening method, fermentation, isolation, and biological activities of abyssomicins and the taxonomy of the producing strain [2]. Structure elucidation of abyssomicins and their mode of action was done by the group of Roderich Süssmuth (Technical University Berlin) [3][4][5]. The complete characterization of strain AB-18-032 applying polyphasic taxonomy was done by the group of Michael Goodfellow (Newcastle University). Strain AB-18-032 was classified in the genus Verrucosispora as a new species and was named Verrucosispora maris sp. nov. as a type strain [6]. Investigations in the genetic analysis and description of the whole genome sequence of Verrucosispora maris AB-18-023 were done by the groups of Roderich Süssmuth and James E.M Stach (Newcastle University) [7][8]. The structures of the main compounds produced by Verrucosispora maris AB-18-023, abyssomicin C and atrop-abyssomicin C, are shown in Figure 2.
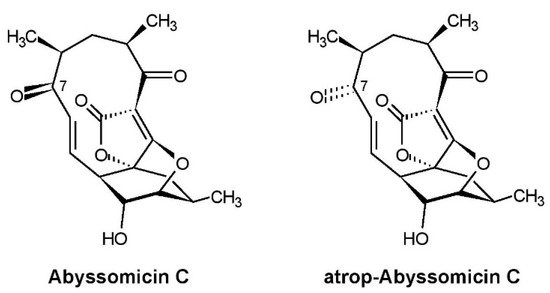
Figure 2. Structures of abyssomicin C and atrop-abyssomicin C produced by Verrucosispora maris AB-18-032.
2. Structure Elucidation and Synthesis Strategies
From abyssomicin B, C, and D, single crystals were obtained, and their relative configuration was determined by X-ray structure. NMR analyses confirmed the X-ray data [3]. The absolute stereochemistry was determined by both the Mosher method [9] and the Helmchen method [10].
A detailed analysis of the culture filtrate from fermentations of strain AB-18-032 revealed two additional signals in the chromatogram of HPLC-DAD-ESI-MS runs that were related to abyssomicins according to their UV-visible properties, and besides abyssomicin C, the presence of atrop-abyssomicin C as the main product of strain AB-18-032 could be confirmed [4]. The compounds, abyssomicin G and H, were isolated from the culture filtrate of the fermentation of strain AB-18-032 in analogy to abyssomicins B–D, and the purified compounds were analyzed by mass spectrometry and 1D and 2D NMR spectroscopy. The structures of all abyssomicins isolated from strain AB-18-032 are shown in Figure 3.
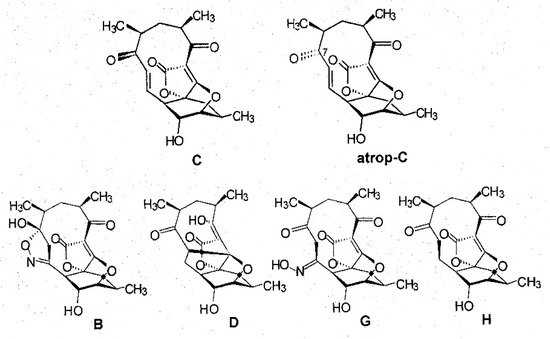
Figure 3. Structures of abyssomicins produced by Verrucosispora maris AB-18-032.
Abyssomicin C was the topic of several total synthetic strategies, based on its unique and complex spirotetronate structure, and on the need for structure optimization with regard to clinical applications. The group of Erik J. Sorensen published a Diels–Alder macrocyclization that enabled an efficient asymmetric synthesis of abyssomicin C [11]. The group of Martin E. Maier prepared the synthesis of the core structure of abyssomicin C containing an oxybicyclooctane ring and a tetronate by a Diels–Alder strategy [12]. A further strategy for the synthesis of abyssomicins C and D was reported by the group of Barry B. Snider [13]. While working on another route for the total synthesis of abyssomicin C, the group of Kyriacos C. Nicolaou discovered atrop-abyssomicin C, a novel isomer of abyssomicin C [14]. Atrop-abyssomicin C was found simultaneously as a natural product of Verrucosispora maris AB-18-032 [4]. The importance of spirotetronate antibiotics and their improved synthetic routes are highlighted in two review articles by the group of E.A. Theodorakis [15][16].
Continuous interest in abyssomicins has been shown by Vidali et al., applying biomimetic approaches toward the synthesis of abyssomicin C and atrop-abyssomicin C based on an intramolecular Diels–Alder reaction of a butenolide derivative attached to a keto-triene side chain [17].
3. Biological Activity and Mode of Action
Among all abyssomicins, only abyssomicin C and atrop-abyssomicin C showed a strong antimicrobial activity, which is restricted to Gram-positive bacteria. The MIC value of atrop-abyssomicin C against multi-resistant Staphylococcus aureus N313 was 3.5 µg/mL. MIC values in the range of 13–16 µg/mL were determined against vancomycin-resistant S. aureus Mu50, Enterococcus faecalis VanA, VanB, and E. faecium VanA, and against multi-resistant S. epidermidis CNS 184. Abyssomicin C showed 1.5-fold lowered MIC values compared to atrop-abyssomicin C. Gram-negative bacteria, filamentous fungi, and yeasts were not sensitive against abyssomicin C and atrop-abyssomicin C [2][4].
The antitumor activity of atrop-abyssomicin C was tested against various human tumor cell lines, such as HM02 (gastric adenocarcinoma), Hep G2 (hepatocellular carcinoma), and MCF7 (breast carcinoma), exhibiting a moderate inhibition with GI50 values of 7.9, 1.5, and 6.1 µg/mL, respectively. Atrop-abyssomicin C showed a slight activity against Trypanosoma brucei rhodesiense, with an IC50 value of 0.68 µg/mL.
Abyssomicin C and atrop-abyssomicin C were the first known natural inhibitors of pAba biosynthesis. Two enzymes catalyze pAba biosynthesis from chorismate, 4-amino-4-deoxychorismate (ADC) synthase, which converts chorismate and glutamine into ADC and glutamate, and ADC lyase, which catalyzes an elimination reaction of ADC to produce pAba. ADC synthase is a heterodimer composed of two nonidentical subunits, PabA and PabB. PabA functions as a glutamine amidotransferase, while PabB catalyzes the substitution of the chorismate 4-hydroxy group by an amino group while retaining the original configuration. The Süssmuth group has shown that the PabB subunit of ADC synthase is the molecular target of abyssomicin C and atrop-abyssomicin C. The antibiotics act as covalent binders in a Michael addition to the side chain of Cys 263, located in the proximity of the active site of PabB, and inhibit pAba formation and, consequently, folate biosynthesis [5]. The increased antimicrobial activity of atrop-abyssomicin C can be explained by its more powerful Michael acceptor properties [14].
The oxabicyclooctane ring system of abyssomicin C, and atrop-abyssomicin B shows a striking similarity to one solution conformation of chorismate, which suggests that both antibiotics act as substrate mimetics (Figure 4).
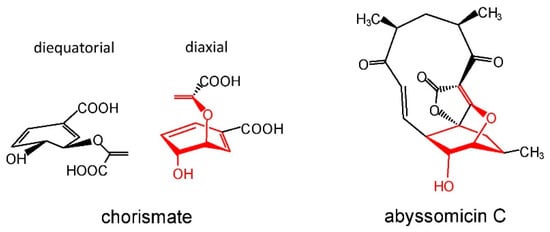
References
- Fiedler, H.-P.; Süssmuth, R.D.; Zähner, H.; Bull, A.T. Polyzyklische Makrolactone. 103 47 472.2 DE, 1 October 2003.
- Riedlinger, J.; Reicke, A.; Zähner, H.; Krismer, B.; Bull, A.T.; Maldonado, L.A.; Ward, A.C.; Goodfellow, M.; Bister, B.; Bischoff, D.; et al. Abyssomicins, inhibitors of the para-aminobenzoic acid pathway produced by the marine Verrucosispora strain AB-18-032. J. Antibiot. 2004, 57, 271–279.
- Bister, B.; Bischoff, D.; Ströbele, M.; Riedlinger, J.; Reicke, A.; Wolter, F.; Bull, A.T.; Zähner, H.; Fiedler, H.-P.; Süssmuth, R.D. Abyssomicin C—A polycyclic antibiotic from a marine Verrucosispora strain as an inhibitor of the p-aminobenzoic acid/tetrahydrofolate biosynthetic pathway. Angew. Chem. Int. Ed. 2004, 43, 2574–2576.
- Keller, S.; Nicholson, G.; Drahl, C.; Sorensen, E.; Fiedler, H.-P.; Süssmuth, R. Abyssomicins G and H and atrop-abyssomicin from the marine Verrucosispora strain AB-18-032. J. Antibiot. 2007, 60, 391–394.
- Keller, S.; Schadt, H.S.; Ortel, I.; Süssmuth, R.D. Action of atrop-abyssomicin C as an inhibitor of 4-amino-4-deoxychorismate synthase PabB. Angew. Chem. Int. Ed. 2007, 46, 8284–8286.
- Goodfellow, M.; Stach, J.E.M.; Brown, R.; Bonda, A.N.V.; Jones, A.L.; Mexson, J.; Fiedler, H.-P.; Zucci, T.D.; Bull, A.T. Verrucosispora maris sp. nov., a novel deep-sea actinomycete isolated from a marine sediment which produces abyssomicins. Antonie Leeuwenhoek 2012, 101, 185–193.
- Gottardi, E.M.; Krawczyk, J.M.; von Suchodoletz, H.; Schadt, S.; Mühlenweg, A.; Uguru, G.C.; Pelzer, S.; Fiedler, H.-P.; Bibb, M.J.; Stach, J.E.M.; et al. Abyssomicin biosynthesis: Formation of an unusual polyketide, antibiotic-feeding studies and genetic analysis. ChemBioChem 2011, 12, 1401–1410.
- Roh, H.; Uguru, G.C.; Ko, H.-J.; Kim, S.; Kim, B.-Y.; Goodfellow, M.; Bull, A.T.; Kim, K.H.; Bibb, M.J.; Choi, I.-G.; et al. Genome sequence of the abyssomicin and proximicin-producing marine actinomycete Verrucosispora maris AB-18-032. J. Bacteriol. 2011, 193, 3391–3392.
- Braun, S.; Kalinowski, H.-O.; Berger, S. 150 and More Basic NMR Experiments; Wiley-VCH: Weinheim, Germany, 1998; pp. 266–268.
- Helmchen, G. Über eine neue Methode zur Bestimmung der absoluten Konfiguration von chiralen sekundären Alkoholen und Aminen: NMR-Spektroskopie von diastereomeren Estern und Amiden der α-Phenylbutter- und Hydratropasäure. Tetrahedron Lett. 1974, 15, 1527–1530.
- Zapf, C.W.; Harrrison, B.A.; Drahl, C.; Sorensen, E.J. A Diels-Alder macrocyclization enables an efficient asymmetric synthesis of the antibacterial natural product abyssomicin C. Angew. Chem. Int. Ed. 2005, 44, 6533–6537.
- Rath, J.-P.; Kinast, S.; Maier, M.E. Synthesis of the fully functionalized core structure of the antibiotic abyssomicin C. Org. Lett. 2005, 7, 3089–3092.
- Snider, B.B.; Zou, Y. Synthesis of the carbocyclic skeleton of abyssomicins C and D. Org. Lett. 2005, 7, 4939–4941.
- Nicolaou, K.C.; Harrison, S.T. Total synthesis of abyssomicin C and atrop-abyssomicin C. J. Am. Chem. Soc. 2007, 129, 429–440.
- Lacoske, M.H.; Theodorakis, E.A. Spirotetronate polyketides as leads in drug discovery. J. Nat. Prod. 2015, 78, 562–575.
- Braddock, A.A.; Theodorakis, E.A. Marine spirotetronates: Biosynthetic edifices that inspire drug discovery. Mar. Drugs 2019, 17, 232.
- Vidali, V.P.; Canko, A.; Peroulias, A.D.; Georgas, E.T.; Bouzas, E.; Herniman, J.M.; Couladouros, E.A. An improved biomimetic formal synthesis of abyssomicin C and atrop-abyssomicin C. Eur. J. Org. Chem. 2020, 29, 4547–4557.

