 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | ALESSIA DI GIANCAMILLO | + 2147 word(s) | 2147 | 2021-05-25 11:42:50 | | | |
| 2 | Lily Guo | -8 word(s) | 2139 | 2021-06-04 03:03:50 | | |
Video Upload Options
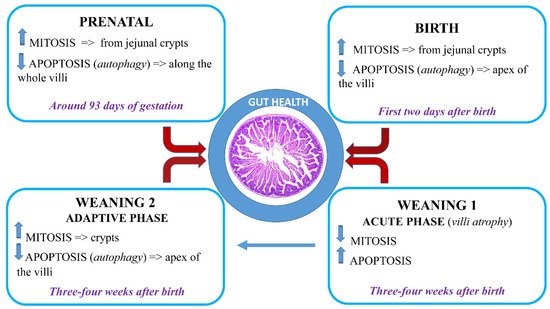
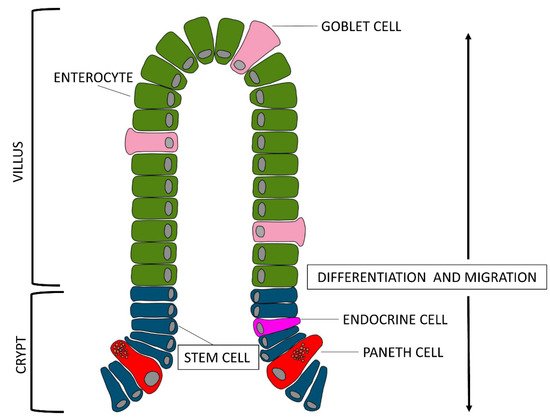
During the prenatal, neonatal, and weaning periods, the porcine gastrointestinal tract undergoes several morpho-functional, changes together with substantial modification of the microbial ecosystem. Modifications of the overall structure of the small intestine also occur, as well as a rapid increase of the volume, mainly in the last period of gestation: intestinal villi, starting from jejunum, appears shortly before the sixth week of gestation, and towards the end of the third month, epithelial cells diversify into enterocytes, goblet cells, endocrine, and Paneth cells. Moreover, in the neonatal period, colostrum induces an increase in intestinal weight, absorptive area, and brush border enzyme activities: intestine doubles its weight and increases the length by 30% within three days of birth.
1. Introduction
2. Gut Anatomy in Brief
Focus on the Small Intestine
3. The Development of the Small Intestine during the Preterm Period
References
- Domeneghini, C.; di Giancamillo, A.; Arrighi, S.; Bosi, G. Gut-trophic feed additives and their effects upon the gut structure and intestinal metabolism: State of the art in the pig, and perspectives towards humans. Histol. Histopathol. Cell. Mol. Biol. 2006, 21, 273–283.
- Pluske, J.R.; Turpin, D.L.; Kim, J.-C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4, 187–196.
- Lærke, H.N.; Hedemann, M.S. The Digestive System of The Pig (Chapter 5). In Nutritional Physiology of Pigs—Online Publication; Videncenter for Svineproduktion: Foulum, Denmark, 2012.
- Moore, B.A.; Morris, G.P.; Vanner, S. A novel in vitro model of Brunner’s gland secretion in the guinea pig duodenum. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G477–G485.
- Sumigray, K.D.; Terwilliger, M.; Lechler, T. Morphogenesis and Compartmentalization of the Intestinal Crypt. Dev. Cell 2018, 45, 183–197.e5.
- Modina, S.C.; Polito, U.; Rossi, R.; Corino, C.; di Giancamillo, A. Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets. Anim. Open Access J. MDPI 2019, 9, 1045.
- Zhu, G.; Hu, J.; Xi, R. The cellular niche for intestinal stem cells: A team effort. Cell Regen. 2021, 10, 1.
- Gribble, F.M.; Reimann, F. Function and mechanisms of enteroendocrine cells and gut hormones in metabolism. Nat. Rev. Endocrinol. 2019, 15, 226–237.
- Ouellette, A.J. Paneth Cells. In Physiology of the Gastrointestinal Tract; Elsevier: Amsterdam, The Netherlands, 2012; Volume 2, pp. 1211–1228. ISBN 978-0-12-382026-6.
- van der Hee, B.; Loonen, L.M.P.; Taverne, N.; Taverne-Thiele, J.J.; Smidt, H.; Wells, J.M. Optimized procedures for generating an enhanced, near physiological 2D culture system from porcine intestinal organoids. Stem Cell Res. 2018, 28, 165–171.
- Verdile, N.; Mirmahmoudi, R.; Brevini, T.A.L.; Gandolfi, F. Evolution of pig intestinal stem cells from birth to weaning. Animal 2019, 13, 2830–2839.
- van der Flier, L.G.; Clevers, H. Stem Cells, Self-Renewal, and Differentiation in the Intestinal Epithelium. Annu. Rev. Physiol. 2009, 71, 241–260.
- Sangild, P.T.; Fowden, A.L.; Trahair, J.F. How does the foetal gastrointestinal tract develop in preparation for enteral nutrition after birth? Livest. Prod. Sci. 2000, 66, 141–150.
- Bjornvad, C.R.; Schmidt, M.; Petersen, Y.M.; Jensen, S.K.; Offenberg, H.; Elnif, J.; Sangild, P.T. Preterm birth makes the immature intestine sensitive to feeding-induced intestinal atrophy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1212–R1222.
- Trahair, J.F.; Sangild, P.T. Fetal organ growth in response to oesophageal infusion of amniotic fluid, colostrum, milk or gastrin-releasing peptide: A study in fetal sheep. Reprod. Fertil. Dev. 2000, 12, 87–95.
- Zabielski, R.; Godlewski, M.M.; Guilloteau, P. Control of development of gastrointestinal system in neonates. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59 (Suppl. 1), 35–54.
- Pácha, J. Development of Intestinal Transport Function in Mammals. Physiol. Rev. 2000, 80, 1633–1667.
- Kulkarni, D.H.; Newberry, R.D. Intestinal Macromolecular Transport Supporting Adaptive Immunity. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 729–737.
- van Ginneken, C.; Weyns, A.; van Meir, E. Stereologic evaluation of the pig gastric wall and of somatostatinergic and serotoninergic immunoreactive mucosal cells during perinatal development. Eur. J. Morphol. 2001, 39, 113–120.
- Willemen, S.A.; De Vos, M.; Huygelen, V.; Fransen, E.; Tambuyzer, B.R.; Casteleyn, C.; Van Cruchten, S.; Van Ginneken, C. Ghrelin in the gastrointestinal tract and blood circulation of perinatal low and normal weight piglets. Animal 2013, 7, 1978–1984.
- McPherson, R.L.; Ji, F.; Wu, G.; Blanton, J.R.; Kim, S.W. Growth and compositional changes of fetal tissues in pigs. J. Anim. Sci. 2004, 82, 2534–2540.
- Darwich, A.S.; Aslam, U.; Ashcroft, D.M.; Rostami-Hodjegan, A. Meta-Analysis of the Turnover of Intestinal Epithelia in Preclinical Animal Species and Humans. Drug Metab. Dispos. 2014, 42, 2016–2022.
- Godlewski, M.M.; Słupecka, M.; Woliński, J.; Skrzypek, T.; Skrzypek, H.; Motyl, T.; Zabielski, R. Into the unknown--the death pathways in the neonatal gut epithelium. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2005, 56 (Suppl. 3), 7–24.
- Yen, T.-H.; Wright, N.A. The gastrointestinal tract stem cell niche. Stem Cell Rev. 2006, 2, 203–212.
- Sangild, P.T.; Thymann, T.; Schmidt, M.; Stoll, B.; Burrin, D.G.; Buddington, R.K. Invited review: The preterm pig as a model in pediatric gastroenterology. J. Anim. Sci. 2013, 91, 4713–4729.
- Buddington, R.K.; Sangild, P.T.; Hance, B.; Huang, E.Y.; Black, D.D. Prenatal gastrointestinal development in the pig and responses after preterm birth. J. Anim. Sci. 2012, 90 (Suppl. 4), 290–298.
- Biernat, M.; Woliński, J.; Godlewski, M.M.; Motyl, T.; Morisset, J.; Zabielski, R. Apoptosis in the Gut of Neonatal Piglets; University of Alberta: Edmonton, AB, Canada, 2003; pp. 46–48.
- Levine, B.; Klionsky, D.J. Development by Self-Digestion: Molecular Mechanisms and Biological Functions of Autophagy. Dev. Cell 2004, 6, 463–477.
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075.
- Haq, S.; Grondin, J.; Banskota, S.; Khan, W.I. Autophagy: Roles in intestinal mucosal homeostasis and inflammation. J. Biomed. Sci. 2019, 26, 19.
- Lamparska-Przybysz, M.; Gajkowska, B.; Motyl, T. Cathepsins and BID are involved in the molecular switch between apoptosis and autophagy in breast cancer MCF-7 cells exposed to camptothecin. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2005, 56 (Suppl. 3), 159–179.
- Motyl, T.; Gajkowska, B.; Zarzyńska, J.; Gajewska, M.; Lamparska-Przybysz, M. Apoptosis and autophagy in mammary gland remodeling and breast cancer chemotherapy. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2006, 57 (Suppl. 7), 17–32.


