 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tariq Khan | + 3244 word(s) | 3244 | 2021-05-26 04:02:21 | | | |
| 2 | Peter Tang | Meta information modification | 3244 | 2021-05-31 11:30:53 | | |
Video Upload Options
Antimicrobial resistance is mushrooming as a silent pandemic. It is considered among the most common priority areas identified by both national and international agencies. The global development of multidrug-resistant strains now threatens public health care improvement by introducing antibiotics against infectious agents.
1. Introduction
2. Drivers of Antimicrobial Resistance
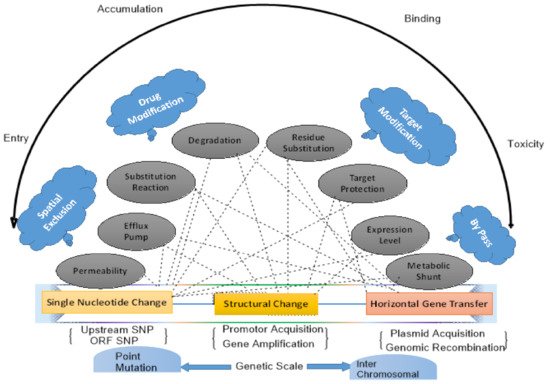
3. Global Dissemination of Antibiotic Resistance
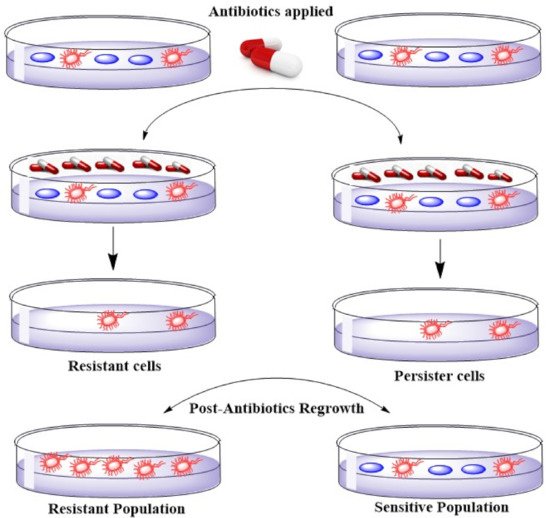
4. Emerging Resistance–Development of Resistant Strains
5. ESKAPE, Healthcare Concomitant Bugs–Bad Bugs with No Drugs
|
Bacterial Strain |
Gram Staining Type |
Resistance Type |
Antibiotics |
Treatment Option |
Resistance Level |
|---|---|---|---|---|---|
|
Acinetobacter |
Negative |
Multidrug |
Ceftazidime, aminoglycoside, fluoroquinolones, carbapenems |
Carbapenems, b-Lactamase inhibitors, Tigecycline, Aminoglycosides, Polymyxin therapy, Synergy, and combination therapy |
High level |
|
E. coli |
Negative |
Multidrug |
Cephalosporins (ESBL-producers), fluoroquinolones, aminoglycosides |
GyrB/ParE programme, EV-035 |
High level |
|
K. pneumoniae |
Negative |
Multidrug |
Cephalosporins (ESBL-producers), fluoroquinolones, aminoglycosides, carbapenems |
POL7080 and ACHN-975 compounds |
High level |
|
P. aeruginosa |
Negative |
Multidrug |
Piperacillin/tazobactam, ceftazidime, ciprofloxacin, aminoglycosides, carbapenems |
POL7080 and ACHN-975 compounds |
High level |
|
Enterococcus spp. |
Positive |
Multidrug |
Ampicillin, aminoglycosides, glycopeptides |
RX-04 lead series, 50S ribosomal subunit; inhibit translation by stabilizing a distorted mode of P-tRNA binding |
High level |
|
S. aureus |
Positive, |
Multidrug |
β-lactam antibiotics (except new anti- methicillin-resistant S. aureus cephalosporins), macrolides, fluoroquinolones, aminoglycosides |
RX-04 lead series, 50S ribosomal subunit; inhibit translation by stabilizing a distorted mode of P-tRNA binding |
High level |
6. General Mechanism of Antimicrobial Resistance
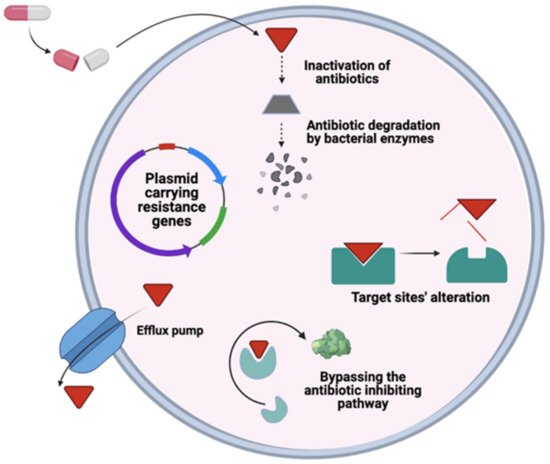
|
Resistance |
Proposed Mechanism |
Examples |
Ref. |
|---|---|---|---|
|
Inactivation of Drug |
Use of hydrolysis or modification |
b-lactamase for b-lactam resistance, acetyltransferases for aminoglycoside resistance |
|
|
Alteration of Target |
Reduction of binding affinity to the drug by bypassing the drug target |
DNA gyrase mutation for fluoroquinolone resistance |
[55] |
|
Drug influx Reduction |
By decreasing permeability |
Gram-negative outer membrane |
[56] |
|
Extrusion of Drug |
Efflux pumps |
Accessory membrane fusion proteins |
[57] |
|
Horizontal gene transfer |
By resistance determinants from other microorganisms |
[58] |
7. Alternative Mechanisms for Combating Multidrug Resistance in ESKAPE Pathogens
7.1. CRISPR-Cas9
7.2. Nanotechnology and Nanoparticles to Combat Multidrug Resistance
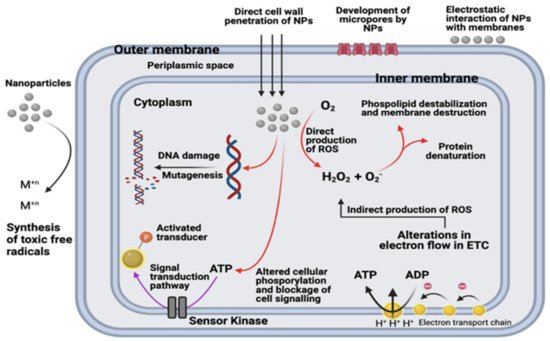
|
Nanoparticles (NP) |
Mode of Action/Mechanism of Nanoparticles Against ESKAPE Pathogens |
Antibiotic Used |
Microorganism |
Synergic Effects (Antibiotics-Nanoparticles) |
Ref. |
|---|---|---|---|---|---|
|
AgNPs |
Damage the bacterial cell membrane and disrupt the activity of membranous enzymes. Cell wall distraction by cell DNA was condensed to a tension state and could have lost its replicating abilities |
Doxycycline |
K. pneumoniae |
Observed |
[68] |
|
Gentamicin and Neomycin |
S. aureus |
AgNPs + Gentamicin showed resistance in 50% strains while AgNPs + Neomycin showed synergy 45% of the strains. |
[69] |
||
|
E. coli, S. aureus |
Observed increase in activity was such that Erythromycin showed 18.9.6%, Kanamycin = 27.9.3%, Chloramphenicol = 18.1.3%, and Ampicillin = 74.8.9% |
[69] |
|||
|
β-Lactam, cefotaxime |
E. coli, S. aureus |
Synergistic increase in activity was such that 17.2%, 13.5% for E. coli and S. aureus, respectively |
[70] |
||
|
Ampicillin, chloramphenicol, and kanamycin |
S. aureus, E. coli, and P. aeruginosa |
Synergistic effects observed |
[71] |
||
|
Beta-lactam: cephem |
S. aureus |
Cephalothin and cefazolin showed a 30% increase in activity when used in combination with 20 μg/ mL AgNPs against Micrococcus luteus, and Bacillus subtilis |
[72] |
||
|
AuNPs |
Disturb membrane potential by inhibiting ATPase activities; inhibit the subunit of the ribosome from binding tRNA. Cellular death induced by gold nanoparticles do not include reactive oxygen species-based mechanisms |
Ampicillin, streptomycin, and kanamycin |
E. coli and S. aureus |
15%, 12%, and 34% increase in inhibition zone for E. coli with A/S/K+Au, respectively; 20%, 109%, and 18% increase in inhibition zone for M. luteus A/S/K+AuNPs, respectively; 12% and 34% increase in inhibition zone for S. aureus with A/ K+AuNPs, respectively |
[73] |
|
Beta lactams: cefaclor |
S. aureus and E. coli |
MICs of cefaclor reduced gold nanoparticles were 10 mg/mL and 100 mg/mL for S. aureus and E. coli, respectively |
[74] |
||
|
ZnONPs |
Interactions between reactive oxygen species and membrane proteins result in cell damage. ZnO-NPs disrupt bacterial cell membrane integrity, reduce cell surface hydrophobicity, and down-regulate the transcription of oxidative stress-resistance genes in bacteria |
Ceftriaxone |
E. coli |
Synergistic antibacterial effects against E. coli have been observed by ZnO nanorods with ceftriaxone |
[75] |
|
Ciprofloxacin |
S. aureus and E. coli |
Increase in inhibition zones in S. aureus = 27% and 22% in E. coli when ciprofloxacin and ZnONPs were applied in synergism |
[76] |
||
|
Beta lactams, aminoglycosides, and azolides |
S. aureus |
The highest increase was observed for penicillin G and amikacin, i.e., 10 mm increase in the zone of inhibition, whereas for clarithromycin, a 2 mm increase had been observed |
[77] |
||
|
TiO2NPs |
Electrostatic interaction between TiO2 NPs and the bacterial cell surface results in suppression of cell division, degradation of the cell wall and cytoplasmic membrane due to the production of reactive oxygen species such as hydroxyl radicals and hydrogen peroxide |
Penicillin G, amikacin, cephalexin, cefotaxime |
MRSA |
10 mm increase in zone size. TiO2 nanoparticles significantly improved antibiotic efficacy against S. aureus when combined with beta-lactams, cephalosporins, and aminoglycosides |
[78] |
|
Fe3O4NPs |
Generation of reactive oxygen species from the disruption of the electronic transport chains owing to the resilient affinity of the iron-based nanoparticles for the cell membrane. Reactive oxygen species generated by Fe3O4 nanoparticles kill bacteria without harming non-bacterial cells |
Streptomycin |
S. aureus, E. coli, and P. aeruginosa |
Zones of inhibition at concentrations (10, 20, 40, and 80): S. aureus (15 mm, 14 mm, 17 mm, 20 mm), E. coli (12 mm, 14 mm, 15 mm, 17 mm), P. aeruginosa (13 mm, 14 mm, 15 mm, 18 mm) |
|
|
Kanamycin and rifampicin |
E. coli and S. aureus |
Kanamycin formed an inhibition zone against both, whereas rifampicin formed an inhibitory zone against S. aureus only |
[81] |
||
|
Amoxicillin |
E. coli and S. aureus |
A total of 9.9% and 8.9% increase in inhibitory effect observed in the presence of Cu NPs for E. coli and S. aureus, respectively |
[80] |
||
|
CuNPs |
Generation of reactive oxygen species, lipid peroxidation, protein oxidation, and DNA degradation. Cu2+ ions released from nanoparticles penetrate bacterial cells and are subsequently oxidized intracellularly |
Amikacin, ciprofloxacin, gentamicin, norfloxacin |
E. coli, P. aeruginosa, Klebsiella spp. S. aureus |
At 60 mg/mL, 18 mm for E. coli, 16 mm for Klebsiella |
[82] |
|
BiNPs |
Production of reactive oxygen species |
Ciprofloxacin, norfloxacin, tetracycline, and metronidazole |
K. pneumoniae |
A synergistic effect was observed between all antibiotics and BiNPs. |
[83] |
|
Cefotaxime, ampicillin, ceftriaxone, cefepime |
E. coli, K. pneumoniae, and P.aeruginosa |
Significant decrease in MIC decrease with cefotaxime and ZnO NPs against K. pneumoniae (85.7%), P. aeruginosa (70%), and E. coli (50%) has been observed. Meanwhile, a decrease in MIC due to ZnO NP with other antibiotics has been observed. |
[84] |
||
|
Norfloxacin, Ofloxacin, and Cephalexin |
P. aeruginosa, E. coli |
Significant increase in inhibition zone of antibiotics with ZnONPshave been observed against all isolates. |
[67] |
References
- Lekshmi, M.; Ammini, P.; Kumar, S.; Varela, M.F. The food production environment and the development of antimicrobial resistance in human pathogens of animal origin. Microorganisms 2017, 5, 11.
- Price, N.; Klein, J.L. Infectious Diseases and Emergencies; Oxford University Press (OUP): Oxford, UK, 2016.
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129.
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19.
- Kümmerer, K.; Henninger, A. Promoting resistance by the emission of antibiotics from hospitals and households into effluent. Clin. Microbiol. Infect. 2003, 9, 1203–1214.
- Franco, B.E.; Martínez, M.A.; Rodríguez, M.A.S.; I Wertheimer, A. The determinants of the antibiotic resistance process. Infect. Drug Resist. 2009, 2, 1–11.
- Rice, L.B. Progress and challenges in implementing the research on ESKAPE pathogens. Infect. Control. Hosp. Epidemiol. 2010, 31, S7–S10.
- Li, X.-Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria. Drugs 2004, 64, 159–204.
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470.
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Genet. 2014, 12, 35–48.
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J. Multistate point-prevalence survey of health care–associated infections. N. Eng. J. Med. 2014, 370, 1198–1208.
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433.
- Levy, S.B. Balancing the drug-resistance equation. Trends Microbiol. 1994, 2, 341–342.
- Levy, S. From Tragedy the Antibiotic Era is Born. The Antibiotic Paradox: How the Misuse of Antibiotics Destroys Their Curative Povers, 2nd ed.; Perseus Publishing: Cambridge, MA, USA, 2002; pp. 1–14.
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281.
- Voorhees, E.M.; Hersh, W.R. Overview of the TREC 2012 Medical Records Track; NIST Publications: Gaithersburg, MD, USA, 2012.
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755.
- Wang, H.; Dzink-Fox, J.L.; Chen, M.; Levy, S.B. Genetic characterization of highly fluoroquinolone-resistant clinical Escherichia coli strains from China: Role ofacrR mutations. Antimicrob. Agents Chemother. 2001, 45, 1515–1521.
- Levy, S.B. Factors impacting on the problem of antibiotic resistance. J. Antimicrob. Chemother. 2002, 49, 25–30.
- Schneiders, T.; Amyes, S.; Levy, S. Role of AcrR and RamA in fluoroquinolone resistance in clinical Klebsiella pneumoniae isolates from Singapore. Antimicrob. Agents Chemother. 2003, 47, 2831–2837.
- Piddock, L.J. Mechanisms of fluoroquinolone resistance: An update 1994–1998. Drugs 1999, 58, 11–18.
- Hiramatsu, K. Vancomycin resistance in staphylococci. Drug Resist. Updat. 1998, 1, 135–150.
- Weigel, L.M.; Clewell, D.B.; Gill, S.R.; Clark, N.C.; McDougal, L.K.; Flannagan, S.E.; Kolonay, J.F.; Shetty, J.; Killgore, G.E.; Tenover, F.C. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571.
- Tenover, F.C.; Weigel, L.M.; Appelbaum, P.C.; McDougal, L.K.; Chaitram, J.; McAllister, S.; Clark, N.; Killgore, G.; O’Hara, C.M.; Jevitt, L.; et al. Vancomycin-resistant Staphylococcus aureus isolate from a patient in Pennsylvania. Antimicrob. Agents Chemother. 2004, 48, 275–280.
- Lipsitch, M.; Samore, M.H. Antimicrobial use and antimicrobial resistance: A population perspective. Emerg. Infect. Dis. 2002, 8, 347–354.
- Schmieder, R.; Edwards, R. Insights into antibiotic resistance through metagenomic approaches. Futur. Microbiol. 2012, 7, 73–89.
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P. Antibiotic-resistance genes in waste water. Trends Microbiol. 2018, 26, 220–228.
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Micobiol. 2010, 8, 251–259.
- Wichmann, F.; Udikovic-Kolic, N.; Andrew, S.; Handelsman, J. Diverse antibiotic resistance genes in dairy cow manure. mBio 2014, 5, e01017-13.
- Clemente, J.C.; Pehrsson, E.C.; Blaser, M.J.; Sandhu, K.; Gao, Z.; Wang, B.; Magris, M.; Hidalgo, G.; Contreras, M.; Noya-Alarcón, Ó.; et al. The microbiome of uncontacted Amerindians. Sci. Adv. 2015, 1, e1500183.
- Moore, A.M.; Ahmadi, S.; Patel, S.; Gibson, M.K.; Wang, B.; Ndao, I.M.; Deych, E.; Shannon, W.D.; Tarr, P.I.; Warner, B.B.; et al. Gut resistome development in healthy twin pairs in the first year of life. Microbiome 2015, 3, 1–10.
- Donato, J.J.; Moe, L.A.; Converse, B.J.; Smart, K.D.; Berklein, F.C.; McManus, P.S.; Handelsman, J. Metagenomic analysis of apple orchard soil reveals antibiotic resistance genes encoding predicted bifunctional proteins. Appl. Environ. Microbiol. 2010, 76, 4396–4401.
- Perron, G.G.; Whyte, L.; Turnbaugh, P.J.; Goordial, J.; Hanage, W.P.; Dantas, G.; Desai, M.M. Functional characterization of bacteria isolated from ancient arctic soil exposes diverse resistance mechanisms to modern antibiotics. PLoS ONE 2015, 10, e0069533.
- Parsley, L.C.; Consuegra, E.J.; Kakirde, K.S.; Land, A.M.; Harper, W.F.; Liles, M.R. Identification of diverse antimicrobial resistance determinants carried on bacterial, plasmid, or viral metagenomes from an activated sludge microbial assemblage. Appl. Environ. Microbiol. 2010, 76, 3753–3757.
- Hawkey, P.M.; Jones, A.M. The changing epidemiology of resistance. J. Antimicrob. Chemother. 2009, 64 (Suppl. S1), i3–i10.
- Cantón, R.; Coque, T.M. The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475.
- Poirel, L.; Liard, A.; Rodriguez-Martinez, J.-M.; Nordmann, P. Vibrionaceae as a possible source of Qnr-like quinolone resistance determinants. J. Antimicrob. Chemother. 2005, 56, 1118–1121.
- Minh, N.N.Q.; Thuong, T.C.; Khuong, H.D.; Nga, T.V.T.; Thompson, C.; Campbell, J.I.; De Jong, M.; Farrar, J.J.; Schultsz, C.; Van Doorn, H.R.; et al. The co-selection of fluoroquinolone resistance genes in the gut flora of vietnamese children. PLoS ONE 2012, 7, e42919.
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606.
- Summers, A.O. Generally overlooked fundamentals of bacterial genetics and ecology. Clin. Infect. Dis. 2002, 34 (Suppl. S3), S85–S92.
- Sidhu, M.S.; Heir, E.; Leegaard, T.; Wiger, K.; Holck, A. Frequency of disinfectant resistance genes and genetic linkage with β-lactamase transposon Tn552 among clinical staphylococci. Antimicrob. Agents Chemother. 2002, 46, 2797–2803.
- Barbosa, T.M.; Levy, S.B. The impact of antibiotic use on resistance development and persistence. Drug Resist. Updat. 2000, 3, 303–311.
- Weinstein, R.A. Controlling antimicrobial resistance in hospitals: Infection control and use of antibiotics. Emerg. Infect. Dis. 2001, 7, 188–192.
- Gagliotti, C.; Balode, A.; Baquero, F.; Degener, J.; Grundmann, H.; Gür, D.; Jarlier, V.; Kahlmeter, G.; Monen, J.; Monnet, D.; et al. Escherichia coli and Staphylococcus aureus: Bad news and good news from the European Antimicrobial Resistance Surveillance Network (EARS-Net, formerly EARSS), 2002 to 2009. Eurosurveillance 2011, 16, 19819.
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Micrbiol. 2007, 5, 48–56.
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect Dis. 2008, 197, 1079–1081.
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976.
- Klevens, R.M.; Edwards, J.R.; Richards, C.L., Jr.; Horan, T.C.; Gaynes, R.P.; Pollock, D.A.; Cardo, D.M. Estimating health care-associated infections and deaths in US hospitals, 2002. Pub. Health Rep. 2007, 122, 160–166.
- Theuretzbacher, U. Global antibacterial resistance: The never-ending story. J. Glob. Antimicrob. Resist. 2013, 1, 63–69.
- Penesyan, A.; Gillings, M.; Paulsen, I. Antibiotic discovery: Combatting bacterial resistance in cells and in biofilm communities. Molecules 2015, 20, 5286–5298.
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist. 2010, 16, 91–104.
- Maisonneuve, E.; Gerdes, K. Molecular mechanisms underlying bacterial persisters. Cell 2014, 157, 539–548.
- Shaw, K.; Rather, P.; Hare, R.; Miller, G. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Mol. Biol. Rev. 1993, 57, 138–163.
- Bush, K.; Fisher, J.F. Epidemiological Expansion, Structural Studies, and Clinical Challenges of New β-Lactamases from Gram-Negative Bacteria. Annu. Rev. Microbiol. 2011, 65, 455–478.
- Hooper, D.C.; Jacoby, G.A. Mechanisms of drug resistance: Quinolone resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 12–31.
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656.
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418.
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the Antibiotic Resistome. Science 2006, 311, 374–377.
- Arcilla, M.S.; van Hattem, J.M.; Haverkate, M.R.; Bootsma, M.C.; van Genderen, P.J.; Goorhuis, A.; Grobusch, M.P.; Lashof, A.M.O.; Molhoek, N.; Schultsz, C.; et al. Import and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae by international travellers (COMBAT study): A prospective, multicentre cohort study. Lancet Infect. Dis. 2017, 17, 78–85.
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Micobiol. 2013, 11, 371–384.
- Zhang, Y.-M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Genet. 2008, 6, 222–233.
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. Nano Lett. 2012, 2, 32.
- Cui, Y.; Zhao, Y.; Tian, Y.; Zhang, W.; Lü, X.; Jiang, X. The molecular mechanism of action of bactericidal gold nanoparticles on Escherichia coli. Biomaterials 2012, 33, 2327–2333.
- Soenen, S.J.; Rivera-Gil, P.; Montenegro, J.-M.; Parak, W.J.; De Smedt, S.C.; Braeckmans, K. Cellular toxicity of inorganic nanoparticles: Common aspects and guidelines for improved nanotoxicity evaluation. Nano Today 2011, 6, 446–465.
- Jin, Y.H.; Dunlap, P.E.; McBride, S.J.; Al-Refai, H.; Bushel, P.R.; Freedman, J.H. Global transcriptome and deletome profiles of yeast exposed to transition metals. PLoS Genet. 2008, 4, e1000053.
- Feng, Q.L.; Wu, J.; Chen, G.; Cui, F.; Kim, T.; Kim, J. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mat. Res. 2000, 52, 662–668.
- Muzammil, S.; Hayat, S.; Fakhar-E-Alam, M.; Aslam, B.; Siddique, M.; Nisar, M.; Saqalein, M.; Atif, M.; Sarwar, A.; Khurshid, A. Nanoantibiotics: Future nanotechnologies to combat antibiotic resistance. Front Biosci. 2018, 10, 352–374.
- Jamaran, S.; Zarif, B.R. Synergistic effect of silver nanoparticles with neomycin or gentamicin antibiotics on mastitis-causing Staphylococcus aureus. Open J. Ecol. 2016, 6, 452–459.
- Fayaz, A.M.; Balaji, K.; Girilal, M.; Yadav, R.; Kalaichelvan, P.T.; Venketesan, R. Biogenic synthesis of silver nanoparticles and their synergistic effect with antibiotics: A study against gram-positive and gram-negative bacteria. Nanomedicine Nanotechnol. Biol. Med. 2010, 6, 103–109.
- Hassan, M.; Ismail, M.; Moharram, A.; Shoreit, A. Synergistic Effect of Biogenic Silver-nanoparticles with β lactam Cefotaxime against Resistant Staphylococcus arlettae AUMC b-163 Isolated from T3A Pharmaceutical Cleanroom, Assiut, Egypt. Am. J. Microbiol. Res. 2016, 4, 132–137.
- Hwang, I.-s.; Hwang, J.H.; Choi, H.; Kim, K.-J.; Lee, D.G. Synergistic effects between silver nanoparticles and antibiotics and the mechanisms involved. J. Med. Microbiol. 2012, 61, 1719–1726.
- Hari, N.; Thomas, T.K.; Nair, A.J. Comparative Study on the Synergistic Action of Differentially Synthesized Silver Nanoparticles with β-Cephem Antibiotics and Chloramphenicol. J. Nanosci. 2014, 2014, 1–8.
- Saha, B.; Bhattacharya, J.; Mukherjee, A.; Ghosh, A.; Santra, C.; Dasgupta, A.K.; Karmakar, P. In vitro structural and functional evaluation of gold nanoparticles conjugated antibiotics. Nanoscale Res. Lett. 2007, 2, 614–622.
- Rai, A.; Prabhune, A.; Perry, C.C. Antibiotic mediated synthesis of gold nanoparticles with potent antimicrobial activity and their application in antimicrobial coatings. J. Mater. Chem. 2010, 20, 6789–6798.
- Luo, Z.; Wu, Q.; Xue, J.; Ding, Y. Selectively enhanced antibacterial effects and ultraviolet activation of antibiotics with ZnO nanorods against Escherichia coli. J. Biomed. Nanotechnol. 2013, 9, 69–76.
- Banoee, M.; Seif, S.; Nazari, Z.E.; Jafari-Fesharaki, P.; Shahverdi, H.R.; Moballegh, A.; Moghaddam, K.M.; Shahverdi, A.R. ZnO nanoparticles enhanced antibacterial activity of ciprofloxacin against Staphylococcus aureus and Escherichia coli. J. Biomed. Mat. Res. Part B Appl. Biomater. 2010, 93, 557–561.
- Thati, V.; Roy, A.S.; Ambika Prasad, M.; Shivannavar, C.; Gaddad, S. Nanostructured zinc oxide enhances the activity of antibiotics against Staphylococcus aureus. J. Biosci. Tech. 2010, 1, 64–69.
- Roy, A.S.; Parveen, A.; Koppalkar, A.R.; Prasad, M.A. Effect of nano-titanium dioxide with different antibiotics against methicillin-resistant Staphylococcus aureus. J. Biomater. Nanobiotechnol. 2010, 1, 37–41.
- Kooti, M.; Gharineh, S.; Mehrkhah, M.; Shaker, A.; Motamedi, H. Preparation and antibacterial activity of CoFe2O4/SiO2/Ag composite impregnated with streptomycin. Chem. Eng. J. 2015, 259, 34–42.
- Khashan, K.S.; Sulaiman, G.M.; Abdulameer, F.A. Synthesis and antibacterial activity of cuo nanoparticles suspension induced by laser ablation in liquid. Arab. J. Sci. Eng. 2016, 41, 301–310.
- Patra, J.K.; Ali, M.S.; Oh, I.-G.; Baek, K.-H. Proteasome inhibitory, antioxidant, and synergistic antibacterial and anticandidal activity of green biosynthesized magnetic Fe3O4 nanoparticles using the aqueous extract of corn (Zea mays L.) ear leaves. Artif. Cells Nanomed. Biotechnol. 2017, 45, 349–356.
- Tanna, J.A.; Chaudhary, R.G.; Gandhare, N.V.; Rai, A.R.; Yerpude, S.; Juneja, H.D. Copper nanoparticles catalysed an efficient one-pot multicomponents synthesis of chromenes derivatives and its antibacterial activity. J. Exp. Nanosci. 2016, 11, 884–900.
- Tarjoman, Z.; Ganji, S.M.; Mehrabian, S. Synergistic effects of the bismuth nanoparticles along with antibiotics on PKS positive Klebsiella pneumoniae isolates from colorectal cancer patients; comparison with quinolone antibiotics. M. Res. J. Med. Med. Sci. 2015, 3, 387–393.
- Bhande, R.M.; Khobragade, C.N.; Mane, R.S.; Bhande, S. Enhanced synergism of antibiotics with zinc oxide nanoparticles against extended spectrum β-lactamase producers implicated in urinary tract infections. J. Nanopart. Res. 2013, 15, 1413.

