 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Iwona Beata Paśmionka | + 12647 word(s) | 12647 | 2021-05-28 08:06:28 | | | |
| 2 | Camila Xu | -5241 word(s) | 7406 | 2021-05-31 09:32:39 | | | | |
| 3 | Camila Xu | -5241 word(s) | 7406 | 2021-05-31 09:33:10 | | |
Video Upload Options
Soil is an important environment in which nitrogen is transformed. The nitrogen cycle in the soil depends on microorganisms, without them, nitrogen transformation processes cannot occur. They determine the continuity of the circulation of this element in the environment and ensure the renewal of its resources. The presence of microorganisms involved in nitrogen transformation in the soil, their activity and the intensity of the reactions taking place are influenced by many factors, including the type of soil, moisture, oxygenation, pH, vegetation and fertilization.
1. Introduction
Nitrogen is a widespread element in nature. It occurs in atmo-, litho-, hydro- and biosphere. It is a biogenic element, necessary for all living organisms, for their growth and development [1]. It occurs in the environment at various degrees of oxidation (from −3 to +5) [2] in molecular, mineral and organic forms. Molecular nitrogen in the N2 form is a gas, the main component of atmospheric air (78%) [3]. The mineral nitrogen is ammonium (NH4+), nitrite (NO2−) and nitrate (NO3−), while nitrogen in the organic form is part of many organic compounds, including amino acids, proteins, nucleic acids, urea, vitamins, chlorophyll, alkaloids [1][3].
This element goes to the soil with mineral and organic fertilizers, plant residues, rainfall, but also through the biological binding of atmospheric nitrogen [1]. It does not accumulate in the soil but is subject to losses as a result of volatilization to the atmosphere (ammonia, nitrogen oxides and molecular nitrogen) and leaching into waters (mainly nitrates) [4]; almost all, because about 99% of nitrogen in the soil occurs in organic form. Most nitrogen in mineral soils is found in humus horizons. Ammonium ions in the soil are bound by clay minerals and organic matter, and nitrate ions (NO3−) are easily soluble—they are found in the soil solution, they can also be eluted [1].
Nitrogen is an element that has the greatest effect on plant yielding [5]. The lack of biologically available nitrogen is often a limiting factor in the growth and production of biomass, even in environments with an appropriate climate and access to water. Therefore, numerous studies are currently carried out on the possibility of increasing the availability of this element for plants without disturbing the natural balance [2]. Nitrogen stimulates the growth of plants, makes them intensely green, regulates the use of nutrients [1]. Both deficiency and excess nitrogen are harmful to plants. The deficiency of this element is manifested by poor leaf formation and yellowing, worse reproduction and soaring. Additionally, the quantity and quality of crops are much smaller [6]. The surplus of nitrogen, on the other hand, causes excessive growth, slowing down of puberty, the tendency to lodging, and sensitivity to diseases, as well as a decline in biological value [1].
The microorganisms that participate in the transformation of nitrogen belong to bacteria, archaea and fungi [2]. The occurrence of microorganisms in the soil is closely related to vegetation. It depends on the type and species of the plant, but also the variety and even the development stage. Microbes meet many important functions for vegetation. They take part in the transformation and sharing of substances, slow down the development of other microorganisms that are often harmful to the plant, which improves plant health. The latest research indicates the possibility of effective use of various types of microbiological biostimulants containing nitrogen microorganisms that have a positive effect on plants and do not disturb soil biodiversity. The composition of microflora in the soil depends to a large extent on plant root exudations. If it were not for the activity of microorganisms, the resources of available nitrogen would be depleted. Microbes participate in the mineralization of organic compounds, thanks to which nitrogen is brought back into circulation [7].
Nitrogen in the soil undergoes many changes, most of which are carried out with the participation of microorganisms. These processes are symbiotic and non-symbiotic atmospheric nitrogen fixation, proteolysis, ammonification, nitrification, denitrification. The binding of molecular nitrogen consists of its reduction to ammonium ions, proteolysis is the hydrolytic breakdown of proteins into amino acids, ammonification is the breakdown of amino acids into ammonium ions. Nitrification is the oxidation of ammonium ions to nitrites (NO2−) and then nitrates (NO3−), while denitrification is the reduction of nitrates to nitrogen in gaseous form [8]. The currently conducted research has significantly expanded the modern knowledge on the metabolic changes occurring during the transformation of nitrogen compounds.
The natural circulation of nitrogen in nature is disturbed by human activities, including the use of nitrogen fertilizers, the burning of fossil fuels, and the release of nitrogen to soil and water along with sewage [2]. The latest scientific achievements make it possible to minimize the effects of disturbances in the nitrogen cycle.
Nitrogen belongs to the group of basic elements that are part of all organisms and therefore plays a key role in the existence of all life forms. Among other things, it builds proteins, nucleic acids and nucleotides, alkaloids, vitamins, plant hormones, chlorophyll and ATP, which is the energy carrier [1][9].
This element, applied in the right dose, affects the proper growth and development of plants, the size of the plant yield, as well as its quality. Thanks to it, the plants propagate better, limiting the dieback of lateral shoots, and thus increasing the number of ears. With the increase in the amount of nitrogen supplied, the protein content in plants increases. Nitrogen gives intensive green color to plants [1].
It is the deficit of this element in the soil that plants are the most sensitive. Nitrogen deficiency is very unfavorable to plants, because their growth and development of both aboveground and underground parts is inhibited. The root system is poorly educated, which affects the poor uptake of other micro- and macroelements [1]. The leaves fall prematurely, change their color—from light green to yellow—chlorosis may appear, and leaf blades are reduced. The fruits are small and often distorted [10].
There are also situations where the plant has an excess of nitrogen, for example as a result of using too much fertilizer. The phenomenon is also not beneficial for plants because it causes lodging and increases susceptibility to diseases. As a result of excess nitrogen, the plants later mature and their biological value deteriorates due to an inappropriate change in the composition of nitrogen compounds [1]. The latest scientific achievements indicate the possibility of using various types of biopreparations without fear of overfertilization. Nitrogen is the most yield-generating macronutrient in the cultivation of all plants. Significant importance in the transformation of its connections is assigned to microorganisms that supply plants with it in the available form of ammonium or nitrate ions [11]. Due to the gradual decrease in the content of organic matter in arable soils and the increase in their acidification resulting from the use of chemical fertilizers, biological agents supporting the development and yielding of plants are sought—in accordance with the principles of sustainable agriculture [9][12]. An interesting alternative is the use of plant growth biostimulants, significantly increasing their biomass and maintaining soil fertility. Such microorganisms include, for example, symbiotic and non-symbiotic atmospheric nitrogen-fixing bacteria [13]. Nodule bacteria are aerobic and thrive best in well-oxygenated soils, and their development stops in the absence of oxygen. They occur in soils with appropriate moisture, because in dry soils plants produce a weak root system and the degree of infection with nodule bacteria is also low. In these cases, soil inoculation with a special vaccine containing active strains of bacteria of the genus Rhizobium gives good results. Scientific studies show that, in addition to gibberellins, they also produce indole-3-acetic acid (IAA), which is one of the natural auxin hormones [14]. IAA secreted by nodule bacteria influences the action of plant auxins during root growth, by stimulating the division and elongation of plant cells, and by promoting the formation of adventitious and lateral roots [15].
The latest research shows that the positive effect of vaccines containing atmospheric nitrogen-fixing bacteria also consists in inhibiting the effects of phytopathogenic microorganisms. Currently, bacterial products are gaining more and more recognition in agricultural and horticultural production [16].
Microbiological stimulators include, among others, plant-growth promoting rhizobacteria (PGPR), mainly from the rhizosphere. Currently, several dozen PGPR strains are known. Plant growth-promoting bacteria belong to different phylogenetic groups. The most numerous group is bacteria of the genus Pseudomonas, as well as Bacillus, Enterobacter and Erwinia [14]. PGPR bacteria affect plant growth both directly and indirectly. The indirect way of influencing plants is related to the protection against the effects of phytopathogens [15]. Direct stimulation consists in supplying plants with minerals, synthesizing plant growth-promoting phytohormones, similar to plant hormones, e.g., auxins, gibberellins, cytokinins, or lowering the level of ethylene adversely affecting plant rooting. Gibberellins stimulate seed germination, stem growth, and induce flowering and flower development. They stimulate the growth of the root and root hairs. These compounds are produced, among others by Acetobacter diazotrophicus, Herbaspirillum seropedicae, Bacillus and Azospirillum. Indole-3-acetic acid (IAA) from the group of other plant hormones (auxins) is a common product synthesized by PGPR bacteria. It is probably involved in establishing bacterial contact with the plant [15]. It can also support the action of plant auxins in root growth by directly influencing the division and elongation of plant cells. Low levels of bacterial IAA stimulate root elongation, while high levels stimulate the formation of lateral and adventitious roots. Since changes in the endogenous pool of plant phytohormones under the influence of PGPR bacteria result in an increase in the root mass, the contact surface of the plant with the soil is increased, which facilitates the availability and more effective uptake of nutrients [14].
The improvement of the nutritional status of plants as a result of the use of microbiological inoculants also takes place by increasing the pool of soil nitrogen resulting from the fixation of atmospheric nitrogen by the diazotrophic bacteria. They contain nitrogenase—an enzyme that reduces atmospheric nitrogen to ammonium. Bacteria with the ability to fix atmospheric nitrogen asymbiotically are e.g., Azoarcus, Pantoea agglomerans, Azotobacter, Azospirillum, Bacillus polymyxa or Herbaspirillum [14][15]. Scientific reports show that in soil conditions favorable for bacteria, about 7–12% of total nitrogen taken up by wheat may be the nitrogen bound by Azospirillum brasilense and Azospirillum lipoferum. PGPR bacteria influence the nitrogen balance not only by the ability to fix atmospheric nitrogen, but also by stimulating the NO3− transporting systems in plants [15].
Microbiological preparations can be added to fertilizers in order to increase the effectiveness of the nutrients contained in the latter. The effectiveness of microbiological preparations depends largely on the species of plants under which they are used. Plants produce various root secretions supporting the activity of inoculated microorganisms, often constituting a substrate for the formation of biologically active substances produced by microorganisms [17][18].
2. Biological Changes of Nitrogen
2.1. Binding of Atmospheric Nitrogen
The process of biological binding of atmospheric nitrogen is carried out by microorganisms referred to as diazotrophs. They belong to the domains Archaea and Bacteria. One can distinguish among them both free-living organisms and those living in symbiosis with plants. Nitrogen binding involves the reduction of molecular nitrogen to ammonium ions. This process is a significant source of nitrogen in the soil. It is a very important part of the circulation of this element because it supplies it in the form available to plants [8].
Molecular nitrogen in the air is an unacceptable compound for plants and animals due to very low reactivity. The process of biological binding of atmospheric nitrogen leads to the formation of ammonia, i.e., nitrogen in the form available to higher organisms [19]. The process is carried out thanks to nitrogenase—an enzyme that accelerates the reduction of the N2 molecule. It runs as follows:
Oxygen is the limiting factor for the enzyme. Nitrogenase is active under anaerobic conditions or in the presence of small concentrations of O2. Higher concentrations of this element cause its inactivation [19]. Microorganisms counteract this differently. Cyanobacteria produce cells with thick walls, the so-called heterocysts to which oxygen does not reach. Aerobic bacteria change the respiratory chain or produce mucus. In the nipples, oxygen-binding leghemoglobin is produced [20].
Factors that affect the effectiveness of nitrogen binding by diazotrophs are also the content of heavy metals in the soil, pesticides and the type of fertilizer. The use of organic fertilizers has a positive effect on the growth of these microorganisms and the intensity of the binding process [21].
Research on the influence of the reaction on the effectiveness of atmospheric nitrogen binding indicates that the concentration of hydrogen ions plays a very important role in this process. Vu et al. [22] investigated the effect of pH on nitrogenase activity, and thus the effectiveness of nitrogen fixation by Acetobacter diazotrophicus—a bacterium found in tropical climate under sugarcane. These bacteria showed the highest efficiency of nitrogen binding at pH = 7, they achieved high efficiency at pH = 5 to 9, while pH = 2 and 11 were limit levels. From this it follows that the reaction plays a significant role in the efficiency of nitrogen fixation by Acetobacter diazotrophicus [19].
2.1.1. Non-Symbiotic Nitrogen Fixation
Non-symbiotic binding of atmospheric nitrogen is carried out by microorganisms that are slowly living in soil, including aerobic bacteria of the genera: Azotobacter, Azotococcus, Azospirillum, Beijerinckia, Derxia, anaerobic: Clostridium, as well as photosynthetic bacteria: Rhodobacter and cyanobacteria: Anabaena and Nostoc. These organisms are widespread in soils and water reservoirs of different climatic zones. Their habitat requirements are heterogeneous [19]. According to Papik et al. [23], 30% of nitrogen absorbed biologically is bound by non-symbiotic microorganisms.
One of the best known nitrogen-fixing bacteria are those of the genus Azotobacter. Their amount in soils in the temperate zone ranges from several to several thousand cells in one gram of soil [24], and the factor limiting their growth is the reaction. They are sensitive to acidification of the environment—they rarely occur at pH below 6 [19]. According to Batista and Dixon [25], bacteria of this type were present in 15 out of 27 soil samples collected from various places, which constituted approx. 55.5%. Most Azotobacter were observed in fertile soils, whose reaction was near neutral. Bacteria of the genus Azotobacter are sensitive to physical and chemical factors, so they can be treated as indicators of environmental changes [5].
The ability to bind atmospheric nitrogen also has many species of endophytic bacteria, for example those from the genera Acetobacter, Herbaspirillum and Azospirillum [26]. Cooperation between endophytes and plants (Table 1) is beneficial for both groups. Endophytic bacteria, in addition to nitrogen binding, also produce substances beneficial to plant roots that ensure their better development [5]. In return, they are better supplied with nutrients and protected against high oxygen concentrations that destroy nitrogenase. Additionally, endophytic bacteria play an important role in soils on which legumes are grown. They stimulate the development of papillary bacteria. Bradyrhizobium japonicum soybean suspension with Pseudomonas fluorescens caused papillation intensity. The situation was similar for the inoculation of red clover Rhizobium leguminosarum and Bacillus brevis [6].
Table 1. Examples of endophytic diazotrophs and plants with which they form associations [6].
| Bacterial Species | Plant |
|---|---|
| Azospirillum brasilense | Millet, sorghum, wheat, corn |
| Azospirillum lipoferum | Millet, wheat, corn |
| Azospirillum amazonense | Wheat |
| Azospirillum doeberainerae | Miscanthus |
| Herbaspirillum seropedicae | Wheat, sugar cane, rice |
| Herbaspirillum rubrisubalbicans | Sugar cane, rice |
| Herbaspirillum frisingense | Miscanthus |
| Acetobacter diazotrophicus | Sugar cane, sweet potatoes, wheat |
The activity of endobiotic diazotrophs increases the nitrogen resources available to plants. For this reason, these microorganisms are stimulants of plant growth [23]. The high nitrogen binding efficiency of Azospirillum bacteria, e.g., A. brasilense in association with maize, may increase yield by 18%. In contrast, Acetobacter diazotrophicus in sugarcane cultivation binds up to 150 kg N·ha−1 per year [6].
Bacteria of the genus Azospirillum are present in moderate climate soils. They live in association with plant roots, including wheat and corn. Their presence in the soil is influenced by physical and chemical factors (nitrogen content, organic matter, salinity, humidity and soil compactness), weather conditions, as well as species and plant development phase and fertilization [5].
The binding of nitrogen gas from the atmosphere to slow-living diazotrophs is low because it is carried out only during the growth of microorganisms. In addition, nitrogen does not end up in the environment immediately, and only after bacterial cells die [24]. Non-symbiotic bacteria bind 10–50 kg N·ha−1 per year [25]. Thanks to bacteria of the genus Azotobacter, which actively binds nitrogen to the soil, goes 1–5 kg N·ha−1 per year [26]. Numbers in the soil of Arthrobacter provide 5–25 kg N·ha−1 per year of this element, as well as cyanobacteria [27][28][29]. Clostridium bind 0.1–0.5 kg N·ha−1 per year [20].
Coexistence between different species of microorganisms can play an important role in non-symbiotic binding of atmospheric nitrogen. Nitrogen-fixing microorganisms in the presence of other microorganisms can absorb nitrogen in much larger quantities. Research conducted by Torres-Cruz et al. [30] proves that there is a close relationship between the nitrogen binding activity and the presence of other microorganisms in the soil. These studies have shown that the use of bacterial strains in appropriate combinations (e.g., Acetobacter diazotrophicus with Pseudomonas fluorescens) increases the nitrogenase activity up to 10-fold.
2.1.2. Symbiotic Nitrogen Fixation
Nitrogen assimilation through the symbiosis of microorganisms with plants accounts for 70–80% of the nitrogen that occurs as a result of biological binding to the global cycle [31].
Symbiotic bacteria include primarily papillary bacteria belonging to the Rhizobiaceae family. These include bacteria of the genus Rhizobium, Sinorhizobium, Mesorhizobium, Bradyrhizobium (Table 2) referred to as rhizobia and living in symbiosis with legumes (Fabaceae). Other examples of endosymbionts include actinomycetes of the genus Frankia [8].
Table 2. Species of papillary bacteria and plants with which they coexist [32].
| Bacterial Species | Plant |
|---|---|
| Rhizobium leguminosarum biowar viviae | Pea |
| Rhizobium leguminosarum biowar trifolii | Clover |
| Rhizobium leguminosarum browar phaseoli | Bean |
| Mesorhizobium lori | Pigweed |
| Sinorhizobium melilori | Lucerne, melilot |
| Azorhizobium caulinodans | Sesbania rostrata |
| Bradyrhizobium sp. | Lupine |
| Bradyrhizobium japonicum | Soya |
Actinomycetes of the genus Frankia are much less specialized diazotrophs than rhizobia. They live in symbiosis with species of angiosperm plants belonging to eight families: Betulaceae, Casuarinaceae, Myricaceae, Elaeagnaceae, Rhamnaceae, Rosaceae, Coriariaceae and Datiscaceae [8]. The most known, however, is their symbiosis with the alder [19]. The amount of nitrogen bound by actinomycetes is similar to the amount of nitrogen bound by rhizobia [8].
For an efficient nitrogen binding process to occur, the interaction of plants with microorganisms is needed. Plants must guarantee the right conditions, i.e., an anaerobic environment and energy sources needed to reduce nitrogen, while bacteria must have the ability to bind this element at an appropriate level beyond their needs.
Plant infection with bacteria is a multistage process. It begins with the mutual recognition of partners, thanks to the secretion of flavonoid compounds by plants, inhibiting the growth of other bacteria, and attracting specific species of rhizobia. The next step is the adhesion of bacteria to root hairs and the production of chitolipooligosaccharides, the so-called Nod factors, that cause root hair modifications, which initiates nipple formation. Rhizobia infect plants by forming infectious threads [8].
Root warts are formed from the cells of the primary bark of the roots. Two types of warts can be distinguished. The first ones are active meristatically, i.e., capable of continuous growth, have an elongated shape, and transport nitrogen in the form of amide compounds (e.g., asparagine, glutamine). The second have limited growth capability, are generally round, and transport nitrogen in the form of ureids (e.g., allantoin) [19]. Inside warts, the bacteria are transformed into bacteroids that have the ability to bind nitrogen. This is due to the expression of nif and fix genes that encode nitrogenase proteins [8]. Plants transport carbohydrates that are a source of energy for bacteroids through conductive beams to warts. In plant cells, leghemoglobin, a protein that protects nitrogenase complex against excessive oxygen concentration, is formed. The nitrogenase is then activated and the nitrogen is bound. Leghemoglobin makes the warts red in color. The ammonium ions formed as a result of binding are incorporated into amino acids in plant cells and then transferred beyond the warts [19]. Symbiosis of papillary bacteria with legumes provides on average 100–400 kg N·ha−1 per year [20].
In the period when the papillary bacteria do not live in symbiosis with plants, they are present in the soil as saprophytes. Their occurrence is related to both soil and climate factors as well as agrotechnical measures. The survival in the soil of symbiotic bacteria without plants with which they live in symbiosis is uneven. A significant influence on their abundance in the soil environment is the concentration of hydrogen ions and the granulometric composition—mainly the content of floatable parts. The majority of symbiotic bacteria is made up of neutral, higher content of floatable parts, liming of soil [31].
The studies of Czaban and Wróblewska [32] show that the presence of papillary bacteria in soils varies. Clover symbiotes (e.g., Rhizobium leguminosarum biovar trifolii) were present in most of the studied soils, symbiotes of lupine and bean (Rhizobium leguminosarum biovar phaseoli, Bradyrhizobium sp.) were less frequent, while alfalfa symbiotes (Sinorhizobium melilori) were found in most soils. It follows that symbionts of red clover or peas can survive longer in the soil than the lupine symbiotes.
2.2. Decomposition of Organic Nitrogen Compounds
The nitrogen contained in the organic matter is decomposed into the ammonium form as a result of the mineralization process, which consists of proteolysis and ammonification. The first stage of mineralization is proteolysis. It consists of the hydrolysis of proteins, resulting in the formation of peptides, followed by amino acids. The proteolysis of individual proteins proceeds at different rates. The degradation time for various proteins ranges from a few minutes to several hours and, in the case of structural proteins, up to several weeks or months. The process is catalyzed by proteolytic enzymes, secreted into the environment by heterotrophic microorganisms. It involves fungi, actinomyces and other bacteria (including those from the genus Proteus, Bacillus and Pseudomonas) [33]. The resulting amino acids can be taken up by microorganisms and plants, but most of them undergo a further transformation. Proteolysis also occurs in proteins other than complex nitrogen-containing organic compounds, e.g., nucleic acids, which, thanks to nucleases, hydrolyse to nucleotides and then nitrogen bases [34].
The second stage of mineralization is ammonification. This process involves converting amino acids to ammonia. Deaminization in soil occurs mainly as a result of biochemical changes due to the presence of enzymes—deaminases secreted by microorganisms [35]. According to Wang et al. [36], the breakdown of amino acids into ammonia and carbon dioxide takes 1–6 days. The ammonia is then transformed into N-NH4. This form can be taken up by plants, be in soil solution, sorption or evaporate into the atmosphere [36]. Numerous bacteria (Bacillus sp., Clostridium sp., Proteus sp., Pseudomonas sp., Serratia sp., Escherichia coli, actinomycetes) and fungi participate in the ammonification process [20]. Due to the large diversity of these microorganisms in the soil, the process takes place regardless of the conditions. Among the ammonifiers are oxygen and anaerobic organisms, preferring acidic and alkaline reaction, adapted to different temperatures and humidity [35].
Factors that influence the process of mineralization of organic nitrogen compounds in the soil include soil properties, humidity, reaction, weather conditions, season, nitrogen fertilization. Acidification of the soil influences the intensification of the ammonification process, which increases the amount of NH4+ in the soil [37].
2.3. Nitrification
2.3.1. The Morphology and Ecology of Nitrifying Bacteria and the Biochemistry of the Nitrification Process
The biological nature of the nitrification process was already established in 1877 by Schloesing and Müntz—they showed that this process involving the oxidation of ammonia to nitrites and nitrites to nitrates is carried out by autotrophic bacteria under extremely aerobic conditions. Winogradsky, on the other hand, as a result of many years of research, isolated nitrifying bacteria, which he divided into two highly specific groups of chemoautotrophic bacteria, then included in one family—Nitrobacteriaceae [38].
Nitrifiers are Gram-negative bacteria, they do not produce spores. In terms of morphology, they constitute a diverse group of rods, spherical and spiral forms. Most nitrifying bacteria produce intracytoplasmic membranes, which may be in the form of flattened lamellar structures or randomly spaced tubes. Some species have the ability to move. The mobile cells are provided with cilia [39].
Nitrifying bacteria are found in most aerobic environments in which the mineralization of organic matter takes place. They are widespread in soil, in fresh, brackish and marine waters, as well as in sewage, sediment and compost [40][41][42][43][44]. Nitrifiers occurring in sewage treatment plants tend to concentrate and grow in tight aggregates, forming structures called cysts.
According to Bergey’s Manual of Determinative Bacteriology [45][46], bacteria belonging to the Nitrobacteriaceae family were divided into 14 species belonging to nine genera (Table 3). Within this family two distinct physiological groups have been distinguished, which do not show any phylogenetic connections between them [47][48][49][50].
Table 3. The occurrence of nitrifying bacteria [45][46].
| Bacterial Species | Habitat |
|---|---|
| I. Bacteria Oxidizing NH3 To NO2− | |
| Nitrosomonas europaea | Soil, sea water, sweet water, sewage |
| Nitrosomonas cryotolerans | Marine environment |
| Nitrosococcus nitrosus | Soil, sea water, industrial sewage |
| Nitrosococcus oceanus | Sea water |
| Nitrosococcus mobilis | Brackish water |
| Nitrosococcus halophilus | Salty lagoons, salt lakes |
| Nitrosospira briensis | Soil, fresh wate |
| Nitrosolobus multiformis | Soil |
| Nitrosovibrio tenuis | Soil |
| II. Bacteria Oxidizing NO2− To NO3− | |
| Nitrobacter winogradskyi | Soil, sea water, fresh water, sewage |
| Nitrobacter hamburgensis | Soil |
| Nitrospina gracilis | Sea water |
| Nitrococcus mobilis | Sea water |
| Nitrospira marina | Soil, sea water, sediments |
The first group of nitrifiers includes bacteria oxidizing ammonia (NH3) to nitrites (NO2−), while the second group includes bacteria oxidizing nitrites (NO2−) to nitrates (NO3−) (Figure 1).
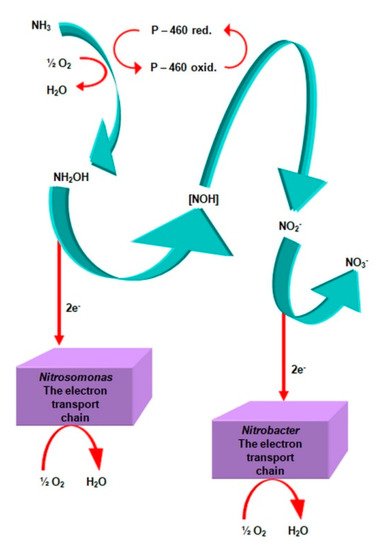
Figure 1. Electron transport reactions during the nitrification process. Compiled by Iwona B. Paśmionka.
Oxidation of ammonia takes place in two stages (Figure 2). They are involved in the nitrifying bacteria of the first phase, among which the metabolic abilities of Nitrosomonas europaea were best studied [51][52][53][54]. In the first stage, due to the attachment of the oxygen molecule, ammonia is oxidized to hydroxylamine.
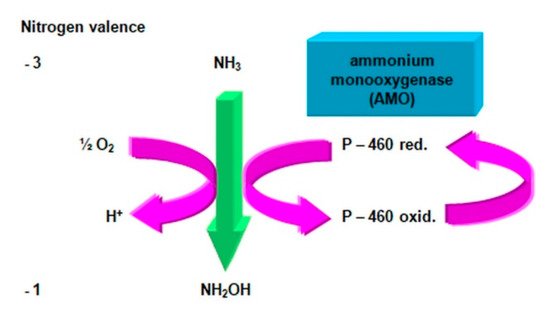
Figure 2. Oxidation of ammonia to hydroxylamine. Compiled by Iwona B. Paśmionka.
This process is catalyzed by ammonium monooxygenase (AMO), an enzyme located in the cell membrane, containing copper and cooperating with cytochrome P-460 [55][56][57].
Oxidation of ammonia to hydroxylamine is an endothermic reaction requiring energy to be supplied in an amount of ΔF = +13.4 kcal per mole of oxidized ammonia. Therefore, the first phase nitrifiers are equipped with a respiratory system of cytochromes, which allows the generation of an adequate amount of energy necessary for this transformation. It is also believed that ammonium monooxygenase may fulfill a dual function:
-
catalyzes the inclusion of one oxygen atom in the substrate (NH3);
It should be emphasized that ammonium monooxygenase tends to bind to other substrates, including those toxic to nitrifiers. These substrates in an irreversible way inactivate the enzyme. In connection with the above, ammonia oxidizing bacteria are extremely sensitive to inhibitory compounds that, even at low concentrations, make it impossible to carry out biochemical reactions necessary for life [54][58][59].
In the second stage of the first nitrification phase, hydroxylamine is oxidized to nitrite ion (Figure 3), through intermediates, which are [60]:
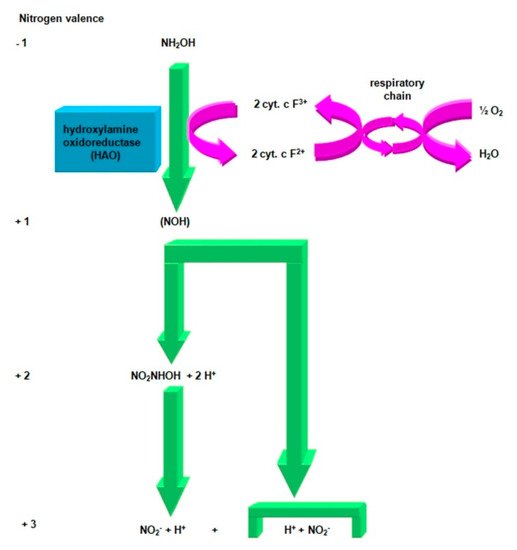
Figure 3. Oxidation of hydroxylamine to nitrite. Compiled by Iwona B. Paśmionka.
-
highly unstable nitrile (NOH) radical;
-
nitrohydroxylamine (NO2NHOH).
This reaction is catalyzed by a hydroxylamine oxidoreductase (HAO), an enzyme located in the periplasm, containing 21 c-type hemises and three P-460 centers. Hydroxylamine oxidoreductase participates in the duplex electron dehydrogenation [54]. C-type cytochromes transfer two electrons from hydroxylamine to conventional cytochrome oxidases (cytochrome aa3 or cytochrome O), and then to the final electron acceptor, i.e., oxygen. In this process, a proton pump is created, and this is the only reaction by which the bacteria responsible for the first phase of nitrification can store usable energy. Two electrons separated during oxidation (NOH) are used to regenerate the reduced form of the cofactor P-460, which allows the oxidation of the next ammonia molecule [61].
Two oxygen atoms are necessary when oxidizing ammonia. One oxygen atom in the resulting nitrite ion is derived from molecular oxygen and the other from water (Figure 4).
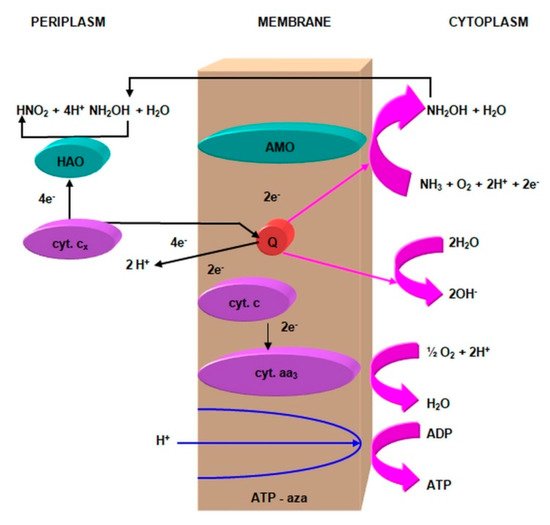
Figure 4. Oxidation of ammonia. Compiled by Iwona B. Paśmionka.
In the second nitrification phase, the nitrite ion oxidizes to the nitrate ion (Figure 5). This process is carried out by a separate group of nitrifying bacteria, with the majority of knowledge about nitrite oxidation coming from studies on Nitrobacter winogradskyi strains [62][63]. Oxidation of nitrites to nitrates is a two-electron process. The electrons are separated from the substrate connected to the enzyme. However, it is not known whether the real substrate in the second nitrification phase is the NO2− ion, undissociated nitrous acid or the hydrated form of one of them [54][58].
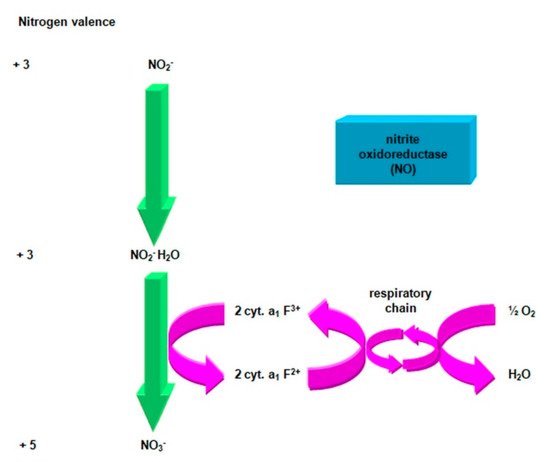
Figure 5. Oxidation of nitrate (III) to nitrate (V). Compiled by Iwona B. Paśmionka.
The oxygen necessary for the formation of nitrate ion comes from the water molecule, not atmospheric oxygen [61].
The nitrite ion is oxidized by a specific enzymatic complex, which consists of:
-
nitrite oxidase (NO), located in the cell membrane, which contains hemi c and a, in the form of cytochromes c1 and a1 and three proteins, with different molar masses, forming the molecule transporting electrons;
-
cytochrome c;
-
unknown quinone;
-
NADH dehydrogenase;
-
molybdenum center;
-
nonhemous iron and sulfur proteins [54].
In the chemolithotrophic metabolism of nitrite-oxidizing bacteria, the nitrite ion has a dual function:
-
is an electron donor for oxidative phosphorylation;
-
is an electron donor for the synthesis of NADH [41].
According to the oxidoreductive reduction potential, for the redox pair NO2−/NO3−, Eo− = + 0.42V, the electrons separated from the nitrite oxidoreductase pass through the respiratory chain to the level of cytochrome a1 to produce energy [58][59]. The classical equation of the second phase of nitrification:
is the result of a three-step reaction:
Presumably, cytochrome a1 is the basic electron acceptor for nitrites (Figure 6). According to the second reaction (b), the reduced cytochrome a1 serves as an electron donor for cytochromes c and aa3 [64].
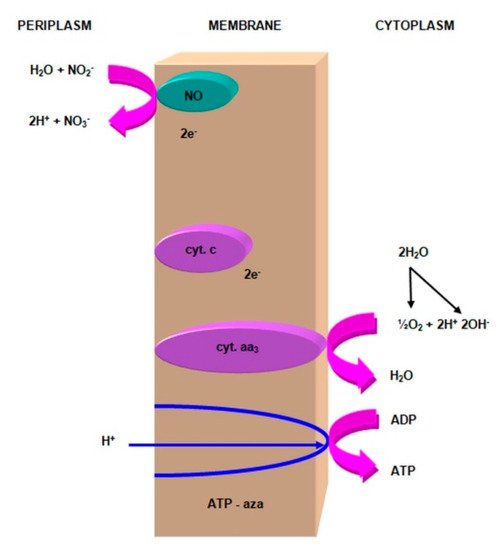
Figure 6. Oxidation of nitrite (NO2−). Compiled by Iwona B. Paśmionka.
According to Ginawi et al. [57], the formation of a reduced form of NADP in Nitrobacter winogradskyi cells requires the reverse flow of electrons, dependent on ATP. This process (requiring energy supply) appears to be catalyzed by a nitrite oxidoreductase. Therefore, the reverse electron transfer system will not be part of the conventional respiratory chain, but part of the own key enzyme complex oxidizing the nitrite ion.
This scheme considers the energetically dependent transport of hydride ion (H−), disconnected from the water molecule. This ion passes through the cytochrome c membrane, where the charge is separated and the proton separated into the periplasm. Then, two electrons from cytochrome c flow through cytochrome aa3 oxidase to molecular oxygen, which in the cytoplasm is reduced to water. The proton pump (Figure 7) is formed in the final stage of nitrite oxidation [64].
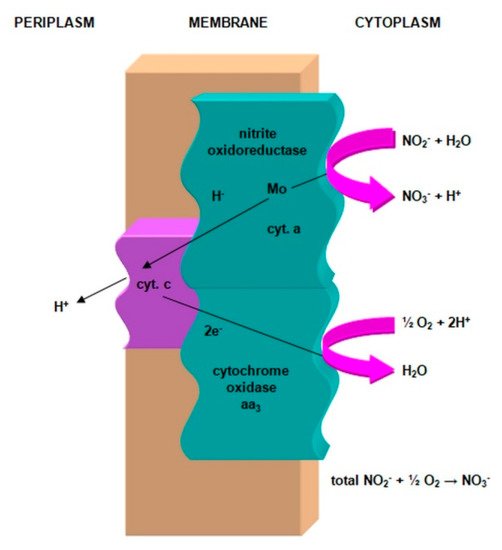
Figure 7. The action of the proton pump during the nitrite oxidation by Nitrobacter winogradskyi. Compiled by Iwona B. Paśmionka.
2.3.2. Heterotrophic Nitrification
Research conducted in recent years has shown that although most nitrifying bacteria belong to strict autotrophs, there are also single heterotrophic organisms that oxidize organic nitrogen compounds to nitrites or nitrates. Such organisms include, among others Aspergillus flavus [39], Nocardia sp., Alcaligenes sp., Agrobacter sp., Arthrobacter sp. etc. [41][54][58][59][60][65][66]. Heterotrophic nitrification plays a major role in soils with an acid reaction (e.g., forest soils). An interesting example is Thiosphaera pantotropha, a bacterium with extremely diverse matabolism, capable of both nitrification and denitrification [67].
Heterotrophs can use both organic and inorganic nitrogen compounds for nitrification. In contrast to autotrophic nitrification, the oxidation of ammonia by heterotrophs is not associated with cell growth [40].
The enzymes responsible for heterotrophic nitrification have not yet been characterized. There are doubts as to whether ammonia is first released from organic compounds and then oxidized by the same mechanisms that use conventional nitrifiers or intermediate products in the form of derivatives of organic nitrogen compounds, from which inorganic compounds are released, such as hydroxylamine, radical nitrile, nitrite [60].
The nitrification process carried out by chemoautotrophic and heterotrophic bacteria can be presented in the general scheme (Figure 8).
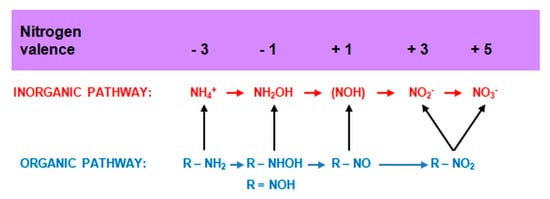
Figure 8. Nitrification pathways. Compiled by Iwona B. Paśmionka.
Nitrification is also carried out by fungi. Aspergillus flavus is the best-known species in this respect [39]. However, the process is of less importance to these organisms because they do not produce energy. The resulting intermediates can inhibit the growth and development of bacteria. The effectiveness of nitrification carried out by fungi is not high, however, in soils populated by these organisms it may play an important role [68].
2.4. Denitrification
Research on the transformation of nitrogen compounds, under the influence of microorganisms, has been conducted for over 100 years. For the first time, the concept of “denitrification” was introduced in 1882 by Gayon and Dupetit, showing the distribution of nitrate (NO3−), with the participation of microorganisms contained in wastewater, in anaerobic conditions [69].
It is now known that denitrification is a biochemical process, carried out by microorganisms, involving the reduction of nitrate (NO3−) under anaerobic or relatively anaerobic conditions. This process takes place in the soil and in the aquatic environment and may be partial or total [70].
Partial denitrification is the reduction of nitrate (NO3−) to nitrite (NO2−) or ammonia, which compounds are usually reoxidized by nitrifiers or assimilated by various microorganisms. Total denitrification is the reduction of nitrate (NO3−) to nitrogen oxides and even gas nitrogen.
Until recently, denitrification was regarded, mainly by farmers, as an undesirable phenomenon associated with high losses of nitrogen in the soil. Currently, as a result of intensification of agriculture, involving the use of high doses of mineral fertilizers and as a result of introducing municipal and industrial sewage into surface waters with a high content of nitrate (NO3−), the soil and waters are contaminated with nitrogen compounds. The use of denitrification bacteria is the most advantageous way to utilize excess nitrogen compounds in water and wastewater. The elimination of nitrate (NO3−) also leads to the suppression of the eutrophication process of surface waters, especially stagnant waters [71].
Denitrification bacteria also play an important role in the breakdown of many organic substances, such as toluene, resorcinol, aromatic heterocyclic compounds. These substances are products of the petrochemical industry and to a large extent contaminate the environment. Nitrogen compounds, mainly oxides, are also a major threat to the atmosphere. The best example of this is the possibility of exposure to nitrous oxide on the stratospheric ozone layer.
Drawing up a balance of nitrogen resources is a very difficult task, because there is a lack of sufficient data on the amount of nitrogen bound in biological processes, as well as returning to the atmosphere, also by biological means. There is also a lack of data on the amount of nitrogen bound to oxides in the processes taking place in the atmosphere. The global amount of nitrogen that gets into the soil with rainfall is around 25 million tons per year. In total, 70% of this amount is previously connected nitrogen and 30% are new nitrogen compounds resulting from the ionization process [72].
It is now known that denitrification is not able to keep up with the rate of nitrogen fixation in industrial technologies. It is also known that the sum of assimilated nitrogen exceeds the amount of nitrogen released into the atmosphere by about 10% [73].
In today’s era, eutrophication has become a major problem, both ecological and economic as well as the landscape. Enriching water with nitrogen and phosphorus is a natural process, but human activity accelerates it very fast. Currently, algal blooms follow human economic activity, wherever industrial centers or centers of intensive farming are created. The substances that cause eutrophication of water come mainly from urban centers, which, even if they are provided with well-planned wastewater treatment plants, enrich the surrounding waters with significant amounts of nitrate (NO3−) and phosphate. This is because of natural microbial degradation processes as well as conventional waste utilization methods, mineralization of organic matter into CO2, nitrate (NO3−) and phosphate, the compounds of which cause the bloom of water reservoirs. Mass algae blooms cause the water surface to be covered with sludge, filter clogging in sewage treatment plants and unpleasant odors resulting from anaerobic decomposition of dying algae. Increased levels of nitrate (NO3−) are also harmful to higher organisms (humans, animals) because it leads to the formation of carcinogenic nitrosamines. Therefore, at the moment there is a need to reduce the content of nitrate (NO3−) in various environments, by intensifying the biological denitrification processes [70][72].
2.4.1. Biochemistry of Nitrate Reduction
Denitrification is a biochemical process of reducing nitrate (NO3−) to nitrous oxide and molecular nitrogen. This process is able to carry out both anaerobic and relatively anaerobic bacteria, having the possibility of biosynthesis of appropriate reducing enzymes, thanks to which they can use nitrate (NO3−) as final electron acceptors [74].
Nitrate respiration occurs in relative anaerobes, including Micrococcus denitrificans, Pseudomonas stutzeri, Pseudomonas aeruginosa, Achromobacter liquefaciens, Achromobacter fisheri, Escherichia coli, Aerobacter aerogenes and many others.
Whereas, according to Maintinguer et al. [75], the ability to carry out denitrification includes numerous bacteria belonging to the genera:
-
Alcaligenes (A. eutrophus, A. faecalis),
-
Arthrobacter (A. citreus, A. simolex, A. terregens),
-
Bacillus (B. licheniformis, B. stearothermophilus),
-
Chromobacterium (C. lividum, C. violaceum),
-
Clostridium (C. aurantobutyricum, C. limosum, C. oroticum, C. rectum),
-
Corynebacterium (C. mycetoides),
-
Cytophaga (C. johansone),
-
Dactylosporangium (D. aurantiacum),
-
Eubacterium (E. nitritogenes),
-
Hyphomicrobium (H. vulgare),
-
Micromonospora (M. chalacea),
-
Mycobacterium (M. intercellulare),
-
Moraxella (M. kingie),
-
Nocardia (N. otididis-caviarum),
-
Paracoccus (P. denitrificans, P. halodenitrificans),
-
Propionibacterium (P. acidi-propionici),
-
Pseudomonas (P. aeruginosa, P. aureofaciens, P. fluorescens, P. stutzeri),
-
Rhizobium (R. leguminosarum, R. japonicum),
-
Rhodopseudomonas (R. capsulata, R. palustris, R. sheroides),
-
Spirillum (S. lipoferum),
-
Streptosporangium (S. pseudovulgare),
-
Thiobacillus (T. denitrificans),
-
Vibrio (V. succinogenes).
An interesting species is Thiobacillus denitrificans, which oxidizes thiosulphates, while reducing nitrate (NO3−) to nitrogen gas [76]. In this species, under aerobic conditions, the final electron acceptor is atmospheric oxygen.
It should be emphasized that many bacteria can use nitrate (NO3−) in two processes that run independently of each other:
-
assimilatory reduction of nitrate (NO3−);
-
dissimilatory reduction of nitrate (NO3−), also known as denitrification.
2.4.2. Assimilatory Reduction of Nitrate (NO3−)
As a result of the assimilatory reduction of nitrate (NO3−), nitrate (NO2−) and ammonia are formed, which microorganisms can use for many syntheses, e.g., amino acids, amines, aminosugars and other compounds. The indirect metabolite during the assimilatory reduction of nitrite (NO3−) is hydroxylamine.
This process can be shown by reaction (Figure 9):
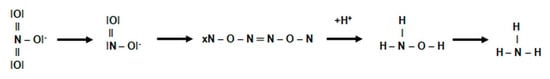
Figure 9. Assimilatory reduction of nitrate.
The process of assimilatory reduction of nitrate (NO3−) is catalyzed by three enzymes:
-
AnaR—assimilatory nitrate reductase, which occurs in the cytoplasm and works under aerobic conditions, initiating a chain of biochemical transformations of assimilated nitrate (NO3−).
-
AniR—assimilatory nitrite reductase, which catalyzes the reduction of nitrite (NO2−). The cofactor of this enzyme is a reduced nicotinamide adenic dinucleotide (NADH + H+) as a hydrogen donor. The enzyme also interacts with cytochrome c.
-
ANH2OHR—assimilatory hydroxylamine reductase, which catalyzes the reduction of hydroxylamine to ammonia in the presence of manganese (IV).
The assimilatory reduction of nitrate (NO3−) therefore requires both energy inputs and a corresponding number of cofactors and hydrogen protons. Therefore, this route must be coupled with the catabolic processes of carbohydrates. It is therefore understandable that this process is much faster under aerobic conditions.
The ability to assimilate nitrate (NO3−) reduction is possessed by bacteria of the genus Bacillus, Aerobacter, Escherichia and many others [77].
2.4.3. Dissimilatory Reduction of Nitrate (NO3−)
During denitrification, as in the assimilatory process of nitrate (NO3−) reduction, intermediate products are formed. The individual stages of denitrification can be represented through of a schematic reaction (Figure 10):
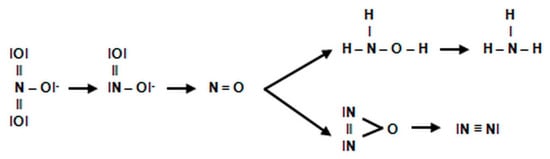
Figure 10. Stages of denitrification.
The denitrification pathway is catalyzed by the following enzymes:
-
DnaR—denitrifying nitrate reductase, which is associated with the cytoplasmic membrane of the cell and initiates chain biochemical transformations of nitrate (NO3−) during denitrification. This enzyme belongs to flavoproteids. In addition to the flavin fragment (FAD), it contains molybdenum and iron. It is active in the presence of cofactors NADH + H+ and FMN and interacts with cytochromes b and c, which transfer electrons to nitrate (NO3−). The activity of denitrifying nitrate reductase is inhibited by molecular oxygen.
The flow of hydrogen protons and electrons during the initial reduction of nitrate (NO3−) can be represented by a schematic chain of reactions (Figure 11).
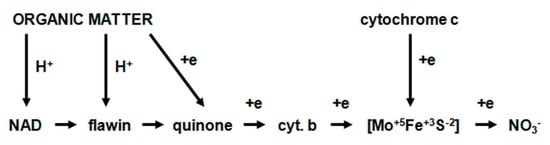
Figure 11. The flow of hydrogen protons and electrons during the initial reduction of nitrate (NO3−).
Although the assimilatory nitrate reductase and denitrifying nitrate reductase differ, both structurally and functionally, they can be produced simultaneously by certain denitrifying bacteria, e.g., Pseudomonas aeruginosa, Pseudomonas stutzeri, Micrococcus denitrificans.
- 2. DniR—denitrifying nitrite reductase. This enzyme catalyzes the reduction of nitrite (NO2−) to nitric oxide. The denitrifying nitrite reductase consists of iron, FAD and a protein called azurine, containing copper. This enzyme interacts with cytochromes c and d in the presence of the cofactor NADH + H+. The optimum activity of this reductase is at 30 °C and pH = 7.0.
- 3. DNOR—denitrifying nitrogenous oxide reductase. This enzyme catalyses the reduction of nitric oxide to nitrous oxide. It contains FAD and works with cytochromes b and c in the presence of cofactor NADH + H+.
- 4. DN2OR—denitrifying nitrous oxide reductase. This enzyme catalyzes the reduction of nitrous oxide to atmospheric nitrogen. Nitrous oxide is the penultimate stage of denitrification in many bacteria, but in some may be the final product of this process. It interacts with cytochromes b and c and probably contains copper. Denitrifying nitrous oxide reductase is extremely sensitive and deactivates after just one hour at room temperature [78].
The probable hydrogen and electron transport pathway during the total denitrification process in Paracoccus denitrificans (Figure 12) can be summarized as follows [79].
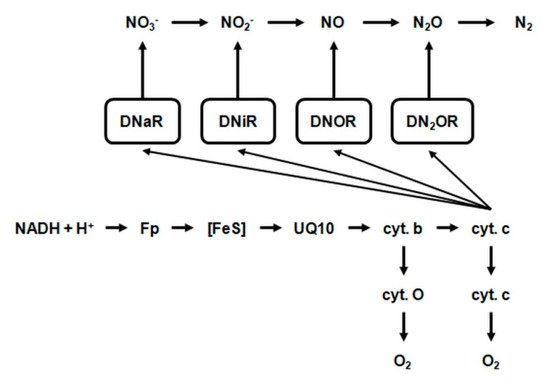
Figure 12. The course of denitrification in Paracoccus denitrificans. Fp—flavoprotein; FeS—iron-sulfur center; UQ—ubiquinone; DnaR—denitrifying nitrate reductase; DniR—denitrifying nitrite reductase; DNOR—denitrifying nitrogenous oxide reductase; DN2OR—denitrifying nitrous oxide reductase.
Probable structure of the iron-sulfur center in Paracoccus denitrificans is shown in Figure 13.
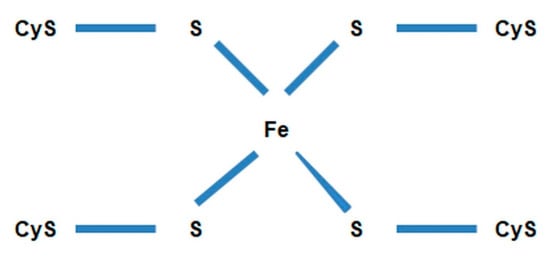
Figure 13. Structure of the iron-sulfur center in Paracoccus denitrificans. CyS—cysteine.
-
5. DNH2OHR—denitrifying hydroxylamine reductase catalyzes the reduction of hydroxylamine to ammonia. This enzyme is activated by manganese, but under anaerobic conditions, in the presence of reduced forms of NADH + H+, pyocyanin and methylene blue, as hydrogen donors, the demand for this metal is much lower.
The hydrogen flow during the reduction of hydroxylamine can be illustrated as follows (Figure 14).
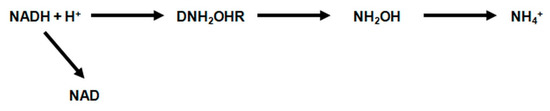
Figure 14. The hydrogen flow during the reduction of hydroxylamine.
During the reduction of nitrate (NO3−) in denitrifying bacteria there is an electron transfer system, similar to oxygen breathing. However, in the case of aerobic respiration, the enzyme transferring electrons to the final acceptor (oxygen) is cytochrome oxidase, while in nitrate respiration they are reductases. This difference affects the efficiency of oxidative phosphorylation, e.g., in Pseudomonas stutzeri, in aerobic breathing, 3 moles of ATP per mole of oxygen are formed, while in nitrate breathing only 2 moles of ATP per mole of nitrate (NO3−).
Denitrification is common among soil microorganisms. Most of them are heterotrophs and facultative anaerobes. They belong to different species, however, the largest group is Bacillus and Pseudomonas [70]. This process is also carried out by some microscopic fungi (Table 4).
Table 4. Examples of microorganisms performing denitrification [70].
| Bacteria | Fungi |
|---|---|
| Achromobacter fisheri Agrobacterium tumefaciens Alcaligenes eutrophus Azospirillum brasilense Bacillus licheniformis Micrococcus denitryficans Nitrosomonas eutropha Pseudomonas aeruginosa Thiobacillus denitryficans |
Aspergillus nidulans Fusarium oxysporum Penicillum sp. |
Low soil pH promotes the development of fungi, including those that produce mycotoxins. These are mainly fungi of Aspergillus sp., Penicillium sp. and Fusarium sp. Their metabolites adversely affect microorganisms. Some of the mycotoxins are characterized by bactericidal and fungicidal activity. These are, for example, aflatoxins, rubratoxins, dicoumarol. These toxins are harmful for atmospheric nitrogen-fixing microorganisms [80].
Denitrification is more intensive in the case of accumulation of nitrite (NO2−) and nitrate (NO3−) in the soil, oxygen deficiency, high humidity. The type of vegetation also has an impact. This is a negative process from an agricultural point of view. Total denitrification leads to nitrogen loss from the soil as a result of the volatilization of nitrogen gas and nitrogen oxides. On the other hand, partial can lead to the formation of nitrite (NO2−) toxic to plants or ammonia, which can also escape from the soil [20].
The research on denitrification that has been going on for more than 100 years has enabled us to expose many biochemical changes occurring in microorganisms during this process. Studies conducted in recent years have explained the chemistry of nitrate (NO3−) reduction, and also contributed to the understanding of the structure and function of enzymes involved in the denitrification process. Despite this, denitrification is not yet fully understood, and the activity of denitrification bacteria has not yet been fully elucidated.
References
- Yu, X.; Zhang, C. Nitrogen cycling and environmental impacts in upland agricultural soils in North China: A review. J. Integr. Agric. 2017, 16, 2848–2862.
- Martinez-Espinosa, R.M.; Cole, J.A.; Richardson, D.J.; Wartmough, N.J. Enzymology and ecology of the nitrogen cycle. Biochem. Soc. Trans. 2011, 39, 175–178.
- Purwanto, P.; Yuwariah, Y.; Sumadi, S.; Simarmata, T. Nitrogenase Activity and IAA Production of Indigenous Diazotroph and Its Effect on Rice Seedling Growth. AGRIVITA J. Agric. Sci. 2017, 39, 31–37.
- Rütting, T.; Aronsson, H.; Delin, S. Efficient use of nitrogen in agriculture. Nutr. Cycling Agroecosyst. 2018, 110, 1–5.
- Gholami, A.; Biyari, A.; Gholipoor, M.; Rahmani, H.A. Growth Promotion of Maize (Zea mays L.) by Plant-Growth-Promoting Rhizobacteria under Field Conditions. Commun. Soil. Sci. Plant Anal. 2012, 43, 1263–1272.
- Ibáñez, F.; Tonelli, M.L.; Muñoz, V.; Figueredo, M.S.; Fabra, A. Bacterial Endophytes of Plants: Diversity, Invasion Mechanisms and Effects on the Host. In Endophytes: Biology and Biotechnology. Sustainable Development and Biodiversity; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 15.
- Zhu, L.; Zhao, Y.; Zhang, W.; Zhou, H.; Chen, X.; Li, Y.; Wei, D.; Wei, Z. Roles of bacterial community in the transformation of organic nitrogen toward enhanced bioavailability during composting with different waste. Bioresour. Technol. 2019, 285, 121326.
- Garrido-Oter, R.; Nakano, R.T.; Dombrowski, N.; Ma, K.W.; McHardy, A.C.; Schulze-Lefert, P. Modular Traits of the Rhizobiales Root Microbiota and Their Evolutionary Relationship with Symbiotic Rhizobia. Cell Host Microbe 2018, 24, 155–167.
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolizm. Sci. Agric. 2016, 73, 18–23.
- Gao, C.; El-Sawah, A.M.; Ali, D.F.I.; Alhaj Hamoud, Y.; Shaghaleh, H.; Sheteiwy, M.S. The Integration of Bio and Organic Fertilizers Improve Plant Growth, Grain Yield, Quality and Metabolism of Hybrid Maize. Agronomy 2020, 10, 319.
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-Based Biostimulants Influence the Agronomical, Physiological, and Qualitative Responses of Baby Rocket Leaves under Diverse Nitrogen Conditions. Plants 2019, 8, 522.
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335.
- Chen, S.K.; Subler, S.; Edwards, C.A. Effects of agricultural biostimulants on soil microbial activity and nitrogen dynamics. Appl. Soil Ecol. 2002, 19, 249–259.
- Sangiorgio, D.; Cellini, A.; Donati, I.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing Climate Change: Application of Microbial Biostimulants to Mitigate Stress in Horticultural Crops. Agronomy 2020, 10, 794.
- De Luca, V.; De Barreda, D.G.; Lidón, A.; Lull, C. Effect of Nitrogen-fixing Microorganisms and Amino Acid-based Biostimulants on Perennial Ryegrass. ASHS 2020, 30, 280–291.
- Berg, S.; Dennis, P.G.; Paungfoo-Lonhienne, C.; Anderson, J.; Robinson, N.; Brackin, R.; Royle, A.; DiBella, L.; Schmidt, S. Effects of commercial microbial biostimulants on soil and root microbial communities and sugarcane yield. Biol. Fertil. Soils 2020, 56, 565–580.
- Chen, S.K.; Edwards, C.A.; Subler, S. The influence of two agricultural biostimulants on nitrogen transformations, microbial activity, and plant growth in soil microcosms. Soil Biol. Biochem. 2003, 35, 9–19.
- Pylak, M.; Oszust, K.; Frąc, M. Review report on the role of bioproducts, biopreparations, biostimulants and microbial inoculants in organic production of fruit. Rev. Environ. Sci. Biotechnol. 2019, 18, 597–616.
- Puri, A.; Padda, K.P.; Chanway, C.P. Nitrogen-fixation by endophytic bacteria in agricultural crops: Recent advances. In Nitrogen in Agriculture—Updates; Khan, A., Fahad, S., Eds.; InTech: Rijeka, Croatia, 2018; pp. 73–94. Available online: (accessed on 11 March 2021).
- Boliyevich, M.S.; Asrorovna, S.G.; Ugli, B.D.K. Biological Nitrogen. AJMR 2020, 9, 66–68.
- Wang, J.; Li, Q.; Shen, C.; Yang, F.; Wang, J.; Ge, Y. Significant dose effects of fertilizers on soil diazotrophic diversity, community composition, and assembly processes ina long-term paddy field fertilization experiment. Land Degrad. Dev. 2020, 1–10.
- Vu HT, L.; Yukphan, P.; Charoenyingcharoen, P.; Malimas, S.; Nguyen, L.K.; Muramatsu, Y.; Yamada, Y. Acetobacter sacchari sp. nov., for a plant growth-promoting acetic acid bacterium isolated in Vietnam. Ann. Microbiol. 2019, 69, 1155–1163.
- Papik, J.; Folkmanova, M.; Polivkova-Majorova, M.; Suman, J.; Uhlik, O. The invisible life inside plants: Deciphering the riddles of endophytic bacterial diversity. Biotechnol. Adv. 2020, 44, 107614.
- Latef, A.A.H.A.; Alhmad, M.F.A.; Kordrostami, M.; Abo–Baker, A.B.A.E.; Zakir, A. Inoculation with Azospirillum lipoferum or Azotobacter chroococcum Reinforces Maize Growth by Improving Physiological Activities under Saline Conditions. J. Plant Growth Regul. 2020, 39, 1293–1306.
- Batista, M.B.; Dixon, R. Manipulating nitrogen regulation in diazotrophic bacteria for agronomic benefit. Biochem. Soc. Trans. 2019, 47, 603–614.
- Kargapolova, K.Y.; Burygin, G.L.; Tkachenko, O.V.; Evseeva, N.V.; Pukhalskiy, Y.V.; Belimov, A.A. Effectiveness of inoculation of in vitro-grown potato microplants with rhizosphere bacteria of the genus Azospirillum. Plant Cell Tissue Organ Cult. 2020, 141, 351–359.
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants 2020, 9, 1011.
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I.M. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 2021, 12, 628379.
- Baldani, J.I.; Baldani, V.D.L. History on the biological nitrogen fixation research in graminaceous plants: Special emphasis on the Brazilian experience. An. Acad. Bras. Ciênc. 2005, 77, 549–579.
- Torres-Cruz, T.J.; Howell, A.J.; Reibold, R.H.; McHugh, T.A.; Eickhoff, M.A.; Reed, S.C. Species-specific nitrogenase activity in lichen-dominated biological soil crusts from the Colorado Plateau, USA. Plant Soil 2018, 429, 113–125.
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Banach, A.; Izak, D.; Stępniewska, Z.; Błaszczyk, M. Metagenomic Analysis of Some Potential Nitrogen-Fixing Bacteria in Arable Soils at Different Formation Processes. Microb. Ecol. 2017, 73, 162–176.
- Czaban, J.; Wróblewska, B. The Effect of Bentonite on the Survival of Azotobacter chroococcum in Sandy Soil in a Long-Term Plot Experiment. Pol. J. Environ. Stud. 2017, 26, 1–8.
- Zheng, Y.; Liang, J.; Zhao, D.L.; Meng, C.; Xu, Z.C.; Xie, Z.H.; Zhang, C.S. The Root Nodule Microbiome of Cultivated and Wild Halophytic Legumes Showed Similar Diversity but Distinct Community Structure in Yellow River Delta Saline Soils. Microorganisms 2020, 8, 207.
- Tian, J.; Wei, K.; Condron, L.M.; Chen, Z.; Xu, Z.; Feng, J.; Chen, L. Effects of elevated nitrogen and precipitation on soil organic nitrogen fractions and nitrogen-mineralizing enzymes in semi-arid steppe and abandoned cropland. Plant Soil 2017, 417, 217–229.
- Fujii, K.; Yamada, T.; Hayakawa, C.; Nakanishi, A.; Funakawa, S. Decoupling of protein depolymerization and ammonification in nitrogen mineralization of acidic forest soils. Appl. Soil Ecol. 2020, 153, 103572.
- Wang, C.; Wang, N.; Zhu, J.; Liu, Y.; Xu, X.; Niu, S.; He, N. Soil Gross N ammonification and nitrification from tropi cal to tempera te forests in ekstern China. Funct. Ecol. 2018, 32, 83–94.
- Mariano, E.; Jones, D.L.; Hill, P.W.; Trivelina, P.C.O. Mineralisation and sorption of dissolved organic nitrogen compounds in litter and soil from sugarcane fields. Soil Biol. Biochem. 2016, 103, 522–532.
- Ye, J.; An, N.; Chen, H.; Ying, Z.; Zhang, S.; Zhao, J. Performance and mechanism of carbon dioxide fixation by a newly isolated chemoautotrophic strain Paracoccus denitrificans PJ-1. Chemosphere 2020, 252, 126473.
- Li, Y.; Chapman, S.J.; Nicol, G.W.; Yao, H. Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 2018, 116, 290–301.
- Meinhardt, K.A.; Stopnisek, N.; Pannu, M.W.; Strand, S.E.; Fransen, S.C.; Casciotti, K.L.; Stahl, D.A. Ammonia-oxidizing bacteria are the primary N2O producers in an ammonia-oxidizing archaea dominated alkaline agricultural soil. Environ. Microbiol. 2018, 20, 2195–2206.
- Jin, P.; Ren, B.; Wang, X.C.; Jin, X.; Shi, X. Mechanism of microbial metabolic responses and ecological system conversion under different nitrogen conditions in sewers. Water Res. 2020, 186, 116320.
- Liu, W.; Nasry, A.A.N.B.; Zhao, J.; Laoyongxay, H.; Dai, W.; Zhao, Q. Start-up of the Simultaneous Nitrification, Anammox, and Denitrification (SNAD) Reactor and Efficacy of a Small Amount of Organic Carbon. Water Air Soil Pollut. 2019, 230, 256.
- Wang, C.; Liu, Y.; Lv, W.; Xia, S.; Han, J.; Wang, Z.; Yu, X. Enhancement of nitrogen removal by supplementing fluidized-carriers into the aerobic tank in a full-scale A2/O system. Sci. Total Environ. 2019, 660, 817–825.
- Wang, H.; He, X.; Nakhla, G.; Zhu, J.; Su, Y.K. Performance and bacterial community structure of a novel inverse fluidized bed bioreactor (IFBBR) treating synthetic municipal wastewater. Sci. Total Environ. 2020, 718, 137288.
- Williams, S.T.; Sharpe, M.E.; Holt, J.G. (Eds.) Bergey’s Manual of Determinative Bacteriology, 8th ed.; Williams & Wilkins Company: Baltimore, ML, USA, 1989; pp. 1808–1834.
- Bergey, D.H.; Holt, J.G. (Eds.) Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins Company: Baltimore, ML, USA, 1994; pp. 447–455.
- Chen, R.; Takemura, Y.; Liu, Y.; Ji, J. Using Partial Nitrification and Anammox To Remove Nitrogen from Low-Strength Wastewater by Co-immobilizing Biofilm inside a Moving Bed Bioreactor. ACS Sustain. Chem. Eng. 2019, 7, 1353–1361.
- Purkhold, U.; Pommerening-Röser, A.; Juretschko, S.; Schmid, M.C.; Koops, H.P.; Wagner, M. Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amaA sequence analysis: Implications for molecular diversity surveys. Appl. Environ. Microbiol. 2000, 66, 5368–5382.
- Cai, M.; Ng, S.K.; Lim, C.K.; Lu, H.; Jia, Y. Physiological and Metagenomic Characterizations of the Synergistic Relationships between Ammonia- and Nitrite-Oxidizing Bacteria in Freshwater Nitrification. Front. Microbiol. 2018, 9, 280.
- Ginawi, A.; Yan, Y. The Function of Ammonia Oxidizers Community in the Environment. PSM Microbiol. 2019, 4, 20–36. Available online: (accessed on 26 March 2021).
- Duddleston, K.N.; Bottomley, P.J.; Porter, A.J.; Arp, D.J. New Insights into Methyl Bromide Cooxidation by Nitrosomonas europaea Obtained by Experimenting with Moderately Low Density Cell Suspensions. Appl. Environ. Microbiol. 2000, 66, 2726–2731.
- Barker, L.K.; Giska, J.R.; Radniecki, T.S.; Semprini, L. Effects of short- and long-term exposure of silver nanoparticles and silver ions to Nitrosomonas europaea biofilms and planktonic cells. Chemosphere 2018, 206, 606–614.
- Soman, C.; Li, D.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159.
- Jeong, D.; Cho, K.; Lee, C.H.; Lee, S.; Bae, H. Effects of salinity on nitrification efficiency and bacterial community structure in a nitrifying osmotic membrane bioreaktor. Process Biochem. 2018, 73, 132–141.
- Bennett, K.; Sadler, N.C.; Wright, A.T.; Hyman, M.R. Activity-Based Protein Profiling of Ammonia Monooxygenase in Nitrosomonas europaea. Appl. Environ. Microbiol. 2016, 82, 2270–2279.
- Wright, C.L.; Schatteman, A.; Crombie, A.T.; Murrell, J.C.; Lehtovirta-Morley, L.E. Inhibition of Ammonia Monooxygenase from Ammonia-Oxidizing Archaea by Linear and Aromatic Alkynes. Appl. Environ. Microbiol. 2020, 86, 1–14.
- Ginawi, A.; Yan, Y. Molecular Techniques Applied to Investigations of Abundance of the Ammonia Oxidizing Bacteria and Ammonia Oxidizing Archaea Microorganisms in the Environment. IOSR J. Environ. Sci. Toxicol. Food Technol. 2019, 13, 19–24.
- Ren, Y.; Ngo, H.H.; Guo, W.; Wang, D.; Peng, L.; Ni, B.J. New perspectives on microbial communities and biological nitrogen removal processes in wastewater treatment systems. Bioresour. Technol. 2020, 297, 122491.
- Cole, J.A. Biodegradation of inorganic nitrogen compounds. In Biochemistry of Microbial Degradation; Ratledge, C., Ed.; Springer: Dordrecht, Germany, 1994; pp. 487–512.
- Liu, H.; Ding, Y.; Zhang, Q.; Liu, X.; Xu, J.; Li, Y.; Di, H. Heterotrophic nitrification and denitrification are the main sources of nitrous oxide in two paddy soils. Plant Soil 2019, 445, 39–53.
- Duan, P.; Shen, H.; Jiang, X.; Yan, X.; Xiong, Z. The contributions of hydroxylamine and nitrite to NO and N2O production in alkaline and acidic vegetable soils. J. Soils Sediments 2020, 20, 2903–2911.
- Ushiki, N.; Jinno, M.; Fujitani, H.; Suenaga, T.; Terada, A.; Tsuneda, S. Nitrite oxidation kinetics of two Nitrospira strains: The quest for competition and ecological niche differentiation. J. Biosci. Bioeng. 2017, 123, 581–589.
- Giguere, A.T.; Taylor, A.E.; Myrold, D.D. Nitrite-oxidizing activity responds to nitrite accumulation in soil. FEMS Microbiol. Ecol. 2018, 94, 1–9.
- Daims, H.; Lücker, S.; Wagner, M. A New Perspective on Microbes Formerly Known as Nitrite-Oxidizing Bacteria. Trends Microbiol. 2016, 24, 699–712.
- Huang, X.; Zhao, J.; Su, J.; Jia, Z.; Shi, X.; Wright, A.L. Neutrophilic bacteria are responsible for autotrophic ammonia oxidation in an acidic forest soil. Soil Biol. Biochem. 2018, 119, 83–89.
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, 1–14.
- Hu, B.; Wang, T.; Ye, J.; Zhao, J.; Yang, L.; Wu, P. Effects of carbon sources and operation modes on the performances of aerobic denitrification process and its microbial community shifts. J. Environ. Manag. 2019, 239, 299–305.
- Forsmark, B.; Nordin, A.; Maaroufi, N.I.; Lundmark, T. Low and High Nitrogen Deposition Rates in Northern Coniferous Forests Have Different Impacts on Aboveground Litter Production, Soil Respiration, and Soil Carbon Stocks. Ecosystems 2020.
- Gayon, U.; Dupetit, G. Sur la fermentation des nitrates en nitrites. C. R. Acad. Sci. 1882, 95, 664–666.
- Guo, B.; Zheng, X.; Yu, J.; Ding, H.; Pan, B.; Lu, S. Dissolved organic carbon enhances both soil N2O production and uptake. Glob. Ecol. Conserv. 2020, 24.
- Liu, C.W.; Sung, Y.; Chen, B.C.; Lai, H.Y. Effects of Nitrogen Fertilizers on the Growth and Nitrate Content of Lettuce (Lactuca sativa L.). Int. J. Environ. Res. Public Health 2014, 11, 4427–4440.
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition-Current Knowledge and Future Directions. Front. Plant Sci. 2017, 19.
- Dey, S.; Awata, T.; Mitsushita, J.; Zhang, D.; Kasai, T.; Matsuura, N.; Katayama, A. Promotion of biological nitrogen fixation activity of an anaerobic consortium using humin as an extracellular electron mediator. Sci. Rep. 2021, 11.
- Tiso, M.; Schechter, A.N. Nitrate Reduction to Nitrite, Nitric Oxide and Ammonia by Gut Bacteria under Physiological Conditions. PLoS ONE 2015.
- Maintinguer, S.I.; Sakamoto, I.K.; Adorno, M.A.T.; Varesche, M.B.A. Evaluation of the microbial diversity of denitrifying bacteria in batch reactor. Braz. J. Chem. Eng. 2013, 30.
- Beristain-Cardoso, R.; Sierra-Alvarez, R.; Rowlette, P.; Razo-Flores, E. Sulfide oxidation under chemolithoautotrophic denitrifying conditions. Biotechnol. Bioeng. 2006, 95, 11481157.
- Sparacino-Watkins, C.; Stolz, J.F.; Basu, P. Nitrate and periplasmic nitrate reductases. Chem. Soc. Rev. 2014, 43, 676–706.
- Ona-Nguema, G.; Guerbois, D.; Pallud, C.; Brest, J.; Abdelmoula, M.; Morin, G. Biogenic Fe(II-III) Hydroxycarbonate Green Rust Enhances Nitrate Removal and Decreases Ammonium Selectivity during Heterotrophic Denitrification. Minerals 2020, 10, 818.
- Olaya-Abril, A.; Hidalgo-Carrillo, J.; Luque-Almagro, V.M.; Fuentes-Almagro, C.; Urbano, F.J.; Moreno-Vivián, C.; Richardson, D.J.; Roldán, M.D. Exploring the Denitrification Proteome of Paracoccus denitrificans PD1222. Front. Microbiol. 2018, 9.
- Natywa, M.; Selwet, M.; Maciejewski, T. Effect of some agrotechnical factors on the number and activity soil microorganisms. Fragm. Agron. 2014, 31, 56–63.


