 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Juan Carlos Corona | + 3287 word(s) | 3287 | 2021-01-04 02:29:42 | | | |
| 2 | Peter Tang | -96 word(s) | 3191 | 2021-03-07 11:47:48 | | |
Video Upload Options
Quercetin is a flavonoid compound present in a wide variety of vegetables and fruit, such as onion, asparagus, red leaf lettuce, cilantro, lovage, dill, capers, apples, and berries. Quercetin represents the highest percentage of total flavonoid intake and is the most important component of flavonol subclass, often the base of other flavonoids. Thus, quercetin has been demonstrated to exert neuroprotective effects in several neurodegenerative disorders as well as antioxidant, anti-inflammatory, anti-cancer, anti-obesity, anti-viral and anti-microbial properties, and cardioprotective and hepatoprotective activities.
1. Introduction
Flavonoids belong to a large group of natural polyphenolic phytochemicals which have been shown to produce several effects such as antioxidative and anti-inflammatory [1][2] and several studies have highlighted the potential beneficial role of flavonoids in numerous neurodegenerative diseases [3][4][5]. There are several subclasses of flavonoids which include the flavones (such as luteolin, rutin, chrysin, baicalin and oroxylin A), flavanones (such as naringenin and hesperidin), isoflavones (such as daidzein and genistein), proanthocyanidins (such as procyanidins), flavanols (such as catechin and epicatechin) and flavonols (such as kaempferol, myricetin and quercetin).
2. Antioxidant Effects of Quercetin
The antioxidant effects of quercetin are elicited owing to the presence of several hydroxyl groups and the basic flavonol skeleton. Quercetin is a potent scavenger of reactive oxygen species (ROS), including superoxide, peroxynitrite and hydrogen peroxide (H2O2), which is also a good lipid peroxidation inhibitor [6][7][8]. One of the antioxidant effects of quercetin depends on the level of glutathione (GSH). Thus, high GSH level promotes the formation of the 6-glutathionyl-Qu (GSQ) complex, which enhances the antioxidant effect, while a low level of GSH increases the extent of cellular damage [9][10].
The effects of quercetin against oxidative stress have been widely demonstrated in diverse conditions both in vitro and in vivo. Accordingly, it was shown that quercetin reduces the H2O2-mediated oxidative stress in yeast mutant cells [11]. Oxidative stress plays a role in the pathophysiology of mental diseases, such as depression or anxiety. Repeated predator stress exposure to mice produced freezing, anxiety-like and depressive-like behaviors. Quercetin showed a protective effect against depression and could alleviate the fear of traumatic events in these mice [12]. Treatment with quercetin showed a protective effect against the oxidative stress produced by cadmium exposure in rats via decrement of the malondialdehyde (MDA) content and an increment in the levels of antioxidant enzymes, superoxide dismutase (SOD), glutathione peroxidase (GPx) and catalase (CAT) [13]. Quercetin protected against doxorubicin-induced cardiomyopathy in rats by increasing the levels of antioxidant defense molecules such as the nuclear factor erythroid 2-related factor 2 (Nrf2), which is a regulator of cellular defense against oxidative stress as well as via the restoration of histological and biochemical defects [14]. Lipopolysaccharide-induced intestinal oxidative stress exerted in broiler chickens and quercetin could significantly inhibit oxidative stress and up-regulate the SOD and GPx levels. Moreover, quercetin relieved mitochondria damage and up-regulated mitochondrial DNA copy number-related gene expression. Furthermore, quercetin promoted Nrf2 activation and increased the gene expression level of heme oxygenase-1 (HO-1), NAD(P)H quinone dehydrogenase 1 (NQO1), and manganese (Mn) SOD2 [15]. Furthermore, quercetin protected against oxidative stress induced by bisphenol-A in rat cardiac mitochondria, considering the improved mitochondrial membrane potential (∆Ψm), GSH level and CAT activity [16]. In addition, quercetin reduced the generation of ROS and nitric oxide (NO) induced by cigarette smoke exposure both in vitro and in vivo, as well as reduced the levels of oxidative stress, leukocyte level and histological pattern changes of the pulmonary parenchyma [17]. Finally, it was demonstrated that quercetin could ameliorate diabetic encephalopathy in mice as a result of reduction in the learning and memory dysfunction, reduced fasting blood glucose and increased insulin sensitivity; in addition, quercetin inhibited oxidative stress and ameliorated neurodegeneration. Moreover, quercetin activated SIRT1, which is an enzyme that deacetylates proteins, which contribute to cellular regulation and inhibited the expression of the endoplasmic reticulum (ER) stress-related proteins [18].
3. Central Nervous System (CNS) Tumors
CNS tumors are a heterogeneous group of neoplasms representing the primary cause of death among children and adolescents. Among all brain tumors, glioblastoma, medulloblastoma, and ependymoma are the most common ones in pediatric populations [19][20][21]. Gliomas are tumors that originate from the glial cells (the supporting cells of CNS) and exists in diverse types based on the involved cell type, for example, astrocytoma, glioblastoma, ependymoma, oligodendroglioma and oligoastrocytoma [19]. Despite the conventional therapy including surgical resection followed by chemotherapy (mainly with temozolomide) and radiotherapy, the effectiveness of this treatment approach is extremely limited and the prognosis is poor with a median survival of 1 year after the therapy. In addition, the most common side-effects of this therapy include cognitive and endocrine dysfunctions, as well as secondary malignancies [20]. Therefore, new natural therapeutic strategies are warranted for use either alone or in combination with other pharmacological agents for the treatment of CNS tumors. Until date, multiple studies have demonstrated the antitumor effect of quercetin on different types of cancer, including breast, esophageal, colorectal, stomach, prostate, lung, ovarian, melanoma and leukemia [22][23][24][25][26]. In addition, quercetin has been reported to induce angiogenesis, the inhibition of proliferation, metastasis, chemoresistance as well as apoptosis both in vitro and in vivo [27].
4. Autism Spectrum Disorder (ASD)
ASD is a heterogeneous and complex neurodevelopmental condition characterized by significant deficiencies in social interaction, communication and repetitive patterns of behavior [28]. People with ASD spend less time engaged in social interaction when compared with non-ASD individuals. Notably, children with ASD do not attribute sufficient value to potential social interactions and favor other environmental stimuli that seems more valuable to them. The worldwide population prevalence of ASD is approximately 1%, and its onset occurs during childhood before the 3 years of age. Autism affects boys more than girls and it has high comorbidity correlation with other neurological disorders [29][30]. The etiology of ASD involves genetic factors or is associated with Rett Syndrome, Fragile X and Down Syndrome with de novo mutations and also environmental factors (exposure to toxins, neurotoxic metals and smoking) as well as certain types of medications during embryonic development, maternal stress, infections during pregnancy and metabolic- and immune- related nutritional factors [31][32][33][34]. The most common treatment in children and young adults with ASD include the use of antipsychotics and the medications used for ADHD and antidepressants [35][36][37]. However, there is evidence of inconsistent efficacy and significant side-effects for most of these pharmacological interventions [38]. In consequence, there is presently a huge interest in the search for alternative ASD treatments, with natural compounds with demonstrated neuroprotective potential via their antioxidant properties and tolerable side-effects [39].
4.1. Protective Effects of Diverse Compounds That Include Quercetin in ASD
It has been reported that oxidative stress in combination with genetic factors and inflammation could be involved in the pathophysiology of ASD [40][41]. Accordingly, the formulation NeuroProtek that contains the flavone luteolin and the flavonoids quercetin and rutin, was applied in 37 children with ASD. The liposomal formulation was found to be safe and well-tolerated, and it showed a positive impact through reduction of brain and gut inflammations [42]. An open-label pilot study of a formulation containing quercetin, luteolin, and the quercetin glycoside rutin was found to effectively to reduce the ASD symptoms with no major adverse effects recorded [43]. Moreover, the induction of developmental hypothyroidism can be used as a model of ASD and can disrupt hippocampal neurogenesis. A diet containing α-lipoic acid as an antioxidant and α-Glycosyl isoquercitrin (AGIQ), which is a mixture of quercetin glycoside consisting of isoquercitrin and its α-glucosylated derivatives along with >10 additional linear glucose moieties, possesses antioxidant effects. The AGIQ-recovered expression of some antioxidant enzyme genes such as NQO1 and thioredoxin 1 (Txn1) in the developmental hypothyroidism rats also restored of NeuN-positive post-mitotic granule cells, parvalbumin and somatostatin-positive interneurons and both antioxidants recovered expression of GABAergic interneuron-related gene orthodenticle homeobox 2 (Otx2) and also AGIQ-recovered expression of glutamate ionotropic receptor AMPA type subunit 3 (Gria3), thereby reversing the disruptive neurogenesis through compensatory responses [44]. Recently, in an experimental model of autism induced by valproic acid during the gestational period, the prenatal treatment with quercetin prevented the behavioral changes and also the treatment with quercetin prevented alterations in the total thiol content as well as changes in the activities of SOD, CAT and glutathione-S-transferase (GST) enzymes in the hippocampus, which in turn prevented the alterations in the CAT and GPx activity in the cerebellum to thereby prevent an increase in the level of ROS, nitrite and thiobarbituric acid reactive substances (TBARS) levels in the striatum and that in the nitrite and CAT alterations in the cerebral cortex [45]. Thus, compounds that contain quercetin can improve the antioxidant defense mechanism. However, more research is warranted to support the efficacy of quercetin alone or in combination with other flavonoids as a possible treatment option for ASD. The outcomes of the protective effects of quercetin in ASD are summarized in Table 1.
Table 1. Summary of protective effects of quercetin in CNS tumors, ASD and ADHD.
|
Type of Study in CNS Tumor |
Effects |
References |
|
Human glioblastoma and rat glioma cell lines |
Reduced cell proliferation and increased antioxidant system |
|
|
Rat glioma and human glioblastoma cell lines |
Induced cell death due to increased oxidative stress and activation of caspases |
|
|
Glioblastoma cell lines |
Anti-inflammatory activity by inhibition of the STAT signaling pathway |
[54] |
|
Glioblastoma and astrocytoma cell lines |
In combination with other compounds induced apoptosis |
|
|
Mouse model glioblastoma and cell line |
Induced autophagy by LC3-I processing and dose-dependency |
[52] |
|
Rat glioma model |
Increased tumor volume and reduced T lymphocyte infiltration and proliferation |
[59] |
|
Medulloblastoma cell lines and mouse model |
Decreased cell migration and growth tumor and increased survival |
|
|
Type of Study in ASD |
Effects |
References |
|
Children |
Safe, well-tolerated and with a positive impact through reduction of brain and gut inflammations |
[42] |
|
Children |
In an open-label pilot study, it effectively reduced symptoms without any adverse effects |
[43] |
|
Developmental hypothyroidism rat model |
Recovered expression of NQO1 and Txn1, restored NeuN-positive granule cells, parvalbumin and somatostatin-positive interneurons and recovered the expressions of Otx2 and Gria3 |
[44] |
|
Prenatal model in rats induced by valproic acid |
Prevented behavioral changes, alterations in total thiol content and changes of SOD, CAT and GST in the hippocampus, prevented the alterations of CAT and GPx in the cerebellum, prevented the increase of ROS, nitrite and TBARS levels in the striatum and prevented nitrite and CAT alterations in the cerebral cortex |
[45] |
|
Type of Study in ADHD |
Effects |
References |
|
Children and adolescents |
In a randomized controlled trial, it showed clinical benefits and tolerable side-effects |
[62] |
|
Children and adolescents |
In a randomized double-blind controlled trial of 8 weeks, it did not improve symptoms |
[63] |
|
Adolescents |
A preliminary study improved some symptoms in patients |
[64] |
|
SH-SY5Y cells |
Increased ATP levels |
[65] |
|
SHR model |
Reduced plasma MDA levels, aortic superoxide production and also improved NO-dependent acetylcholine relaxation, inhibited eNOS phosphorylation and reduced the blood pressure |
[66] |
|
SHR model |
Reduced oxidative stress |
[67] |
|
Amphetamine-induced unilateral rotations in rats |
Reduced rotations and also attenuated the rotenone-induced loss in striatal dopamine, up-regulated mitochondrial complex-I activity and increased CAT and SOD |
[68] |
|
SHR model and H9C2 cells |
Prevented cardiac hypertrophy by suppressing AP1 transcription activity and by increasing activation of PPARγ, also the ultrastructural damage of mitochondria and myofibrils were attenuated |
[69] |
|
MPH-induced hyperlocomotion in mice |
Blocked hyperlocomotion and an increase in lipid peroxidation levels in the striatum and prefrontal cortex regions |
[70] |
5. Attention-Deficit/Hyperactivity Disorder (ADHD)
ADHD is the most prevalent neuropsychiatric disorder, with a worldwide prevalence in children of 7.2% [71][72][73]. The characteristic symptoms of ADHD include hyperactivity, lack of attention and impulsivity [74]. In around 50% of children and adolescents diagnosed with ADHD, the symptoms persist throughout the adult life as well [72][75]. The symptoms of ADHD cause problems in personal, scholar, social, or work performance resulting in the consequences of isolation, lower socioeconomic status and increased risk of substance abuse in adolescence, as well as changes of development of comorbidity and antisocial and delinquent behavior [72][75]. Pharmacological treatment with psychostimulants and non-psychostimulants for this condition include the medications aimed at improving the symptoms of ADHD. Methylphenidate (MPH) and amphetamines increase extracellular dopamine and norepinephrine release in the hippocampus, prefrontal cortex, and striatum, which in turn improve neurotransmitter imbalance and symptoms [76][77][78]. The therapy with atomoxetine, which is a selective norepinephrine reuptake inhibitor, increases extracellular dopamine and norepinephrine release in the cerebellum, prefrontal cortex, hypothalamus and hippocampus, resulting in behavioral improvement [78][79][80]. Nevertheless, psychostimulants induce side-effects such as insomnia, appetite loss, headache, abdomen pain, sleep disturbance and anxiety [4][81]. In addition, non-psychostimulants can produce nausea, diarrhea, somnolence, vomiting, appetite loss, fatigue, dizziness, and changes in the cardiovascular events [4][82]. Extensive studies have suggested that the pathophysiology of ADHD is associated with oxidative stress [74][83][84][85]. Therefore, there is an increasing interest in the search for alternative treatments for ADHD, including the application of bioactive natural compounds owing to their antioxidant properties and considering that these alternative treatments options may have minimal side-effects. The neuroprotective mechanisms of quercetin in pediatric neurological diseases are summarized in Figure 1.
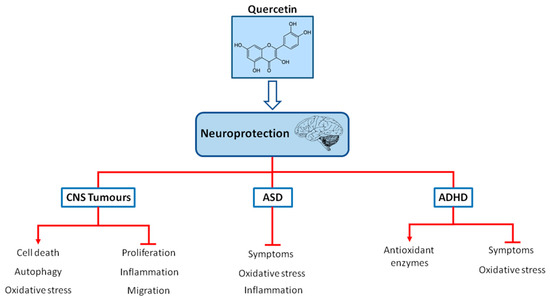
Figure 1. Chemical structure of quercetin and neuroprotection in pediatric neurological diseases (CNS tumors, ASD and ADHD). Quercetin may act as a neuroprotector in pediatric neurological diseases via the regulation of oxidative stress, inflammation, proliferation and improving symptoms and also via increasing antioxidant defenses, autophagy or cell death.
5.1. Protective Effects of Diverse Compounds That Include Quercetin in ADHD
As indicated earlier, accumulating evidence indicate that oxidative stress is involved in the pathophysiology of ADHD [74][83][84][85] and also that the administration of MPH can induce oxidative stress in neurons and thereby neurodegeneration in the cerebral cortex and the hippocampus of animals [86]. Passionflower, which is commonly known as Passiflora incarnata, contains quercetin and other ingredients. In a randomized controlled trial with passionflower in children and adolescents with ADHD, significant clinical benefit and a tolerable side-effect profile was achieved as result of the advantages of passionflower as compared with MPH [62]. St. John’s wort (Hypericum perforatum extract) that contains quercetin among other flavonoids, was used in a randomized double-blinded controlled trial of 8 weeks-duration, but it showed no improvement in the ADHD symptoms [63]. Conversely, a preliminary study demonstrated that treatment with St. John’s wort improved some symptoms in ADHD patients [64]. In vitro, the treatment with St. John’s wort could significantly increase the ATP levels in SH-SY5Y cells [65]. The outcomes of the protective effects of quercetin in ADHD are summarized in Table 1.
The spontaneously hypertensive rat (SHR) is presently used as a validated animal model of ADHD [87]. It was demonstrated that the treatment of SHR with quercetin could reduce its plasma MDA levels and aortic superoxide production as well as improve NO-dependent acetylcholine relaxation, which inhibited endothelial NO synthase (eNOS) phosphorylation and reduced the blood pressure [66]. It was also observed that an increase in oxidative stress in SHR, could be reversed by the treatment with quercetin [67]. Moreover, treatment with quercetin in rats showed significant reduction in the amphetamine-induced unilateral rotations, attenuation of rotenone-induced loss in striatal dopamine, up-regulation of the mitochondrial complex-I activity and increase in the CAT and SOD levels [68]. Quercetin prevented cardiac hypertrophy via suppression of the activator protein 1 (AP1) transcription activity and promotion of the activation of peroxisome proliferator-activated receptor γ (PPARγ). Moreover, the ultrastructural damage of mitochondria and myofibrils in both the SHR and H9C2 cells were found to be attenuated [69]. It was previously demonstrated that PPARγ activation has neuroprotective and antioxidant effects [88]. Finally, chronic treatment with quercetin blocked MPH-induced hyperlocomotion and also blocked the increase in lipid peroxidation levels in the striatum and prefrontal cortex regions [70]. Thus, compounds that contain quercetin could improve the neuroprotection through the activation of antioxidant pathways and by its powerful scavenging properties. Nevertheless, further studies are warranted to verify the efficacy, effects, and dosages of quercetin either alone or in combination with other flavonoids as a possible treatment alternative agent against oxidative stress in ADHD.
6. Protective Effects of Other Flavonoids in Pediatric Neurological Diseases
There are increasing data with flavonoids to verify their efficacy and potential for CNS tumors treatment. Furthermore, it was demonstrated that flavonoids combined with anticancer drugs led to the enhanced anticancer effect. Thus, flavanols such as epigallocatechin gallate which is a constituent of green tea, alone or in combination with temozolomide inhibited neurosphere formation and cell migration of glioma stem-like cells and the treatment whit epigallocatechin gallate, also affected both migration and adhesion of medulloblastoma cells [89][90]. Besides, the flavone chrysin and the combination with cisplatin, induced apoptosis, cell cycle arrest and ∆Ψm loss in human glioma cells [91]. The flavonoid luteolin significantly inhibited glioma cell proliferation, induced apoptosis via MAPK and caspase activation and promote autophagy [92][93]. Moreover, luteolin induced ER stress and mitochondrial dysfunction leading to cell death in glioblastoma cell lines and in an animal model [94]. The combination of the flavonoids luteolin and silibinin effectively blocked angiogenesis and survival pathways leading to induction of apoptosis [95]. Also, the same combination of flavonoids, induced inhibition of growth of glioblastoma cells by the induction of apoptosis and the inhibition of invasion and migration [96].
Alternative approaches with flavonoids are on continuous research to confirm their efficacy and to understand its potential in ASD treatment. The green tea extract (Camellia sinensis) is an important source of flavonols, such as catechins, epicatechin, epigallocatechin and epicatechin-3-gallate and flavonol derivatives such as kaempferol, quercetin and myricetin [97]. Thus, the green tea extract treatment demonstrated amelioration of behavioral and oxidative stress aberrations in an animal model of valproate-induced autism [98]. The treatment of co-ultramicronized palmitoylethanolamide and the flavonoid luteolin in a murine model of autism was efficient in ameliorating social and non-social symptoms via modulation of TNFα and IL-1β immunoreactivity, reduction of GFAP, NF-κB and increased neurogenesis and neuroplasticity in the hippocampus [99]. Moreover, consumption of epigallocatechin-3-gallate, the major compound of catechin in green tea can reverse the behavioural alterations in the sodium valproate-induced autism rat model possibly due to antioxidant effects [100]. Naringenin is a flavanone abundantly found in oranges, grapefruit, and tomato skin. The administration orally for 29 days of naringenin, significantly restored behavioral and biochemical deficits in ASD phenotype in rats induced by propanoic acid [101]. Ginkgo biloba leaves, contains flavonoids (quercetin, kaempferol, and isorhamnetin), terpenoids, and ginkgolic acid. In an observational study of three patients treated with Ginkgo biloba extract, improved aberrant behavior and symptoms of autism [102]. In a double-blind placebo-controlled trial, ginkgo biloba extract was used in patients with autism and the results demonstrated ginkgo biloba no shown significant improvement in the treated group; however, ginkgo biloba was relatively safe and well-tolerated [103].
A growing interest in alternative treatments for ADHD include the research with diverse flavonoids due to their antioxidant properties and because they have minimal side-effects [4]. Baicalin, a major flavonoid isolated from Scutellaria baicalensis Georgi, has antioxidative properties. Thus, baicalin regulated the core symptoms of ADHD and also, improved LDH activity and the synaptosomal ATPase via regulating the AC/cAMP/PKA signaling pathway in the SHR [104][105]. Pycnogenol has antioxidants effects and is extracted from French maritime pine bark (Pinus pinaster), the main ingredients are procyanidins which are a class of flavonoids and phenolic acids. In a randomized, double-blind, placebo-controlled trials, pycnogenol normalized total antioxidant status, catecholamine concentration, reduced oxidative stress, improved hyperactivity and attention in children with ADHD [106][107]. Moreover, the treatment with pycnogenol improved attention, visual-motor coordination, concentration and also reduced significantly the hyperactivity in children with ADHD [108]. Oroxylin A is a flavonoid found in plants Scutellaria baicalensis, Scutellaria lateriflora and the Oroxylum indicum tree. Oroxylin A has activity as a dopamine reuptake inhibitor and is an antagonist of the GABA-A receptor and also has antioxidant effects. Treatment with oroxylin A and a derivate of oroxylin A, improved ADHD-like behaviors in the SHR [109][110].
References
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115.
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042.
- Maher, P. The Potential of Flavonoids for the Treatment of Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3056.
- Corona, J.C. Natural Compounds for the Management of Parkinson’s Disease and Attention-Deficit/Hyperactivity Disorder. Biomed. Res. Int. 2018, 2018, 4067597.
- Khan, H.; Tundis, R.; Ullah, H.; Aschner, M.; Belwal, T.; Mirzaei, H.; Akkol, E.K. Flavonoids targeting NRF2 in neurodegenerative disorders. Food Chem. Toxicol. 2020, 146, 111817.
- Batiha, G.E.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374.
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271.
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119.
- Suganthy, N.; Devi, K.P.; Nabavi, S.F.; Braidy, N.; Nabavi, S.M. Bioactive effects of quercetin in the central nervous system: Focusing on the mechanisms of actions. Biomed. Pharm. 2016, 84, 892–908.
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and cancer chemoprevention. Evid. Based Complement. Altern. Med. 2011, 2011, 591356.
- Alugoju, P.; Periyasamy, L.; Dyavaiah, M. Quercetin enhances stress resistance in Saccharomyces cerevisiae tel1 mutant cells to different stressors. J. Food Sci. Technol. 2018, 55, 1455–1466.
- Anggreini, P.; Ardianto, C.; Rahmadi, M.; Khotib, J. Quercetin attenuates acute predator stress exposure-evoked innate fear and behavioral perturbation. J. Basic Clin. Physiol. Pharm. 2019, 30.
- Wang, J.; Zhu, H.; Wang, K.; Yang, Z.; Liu, Z. Protective effect of quercetin on rat testes against cadmium toxicity by alleviating oxidative stress and autophagy. Environ. Sci. Pollut. Res. Int. 2020.
- Sharma, A.; Parikh, M.; Shah, H.; Gandhi, T. Modulation of Nrf2 by quercetin in doxorubicin-treated rats. Heliyon 2020, 6, e03803.
- Sun, L.; Xu, G.; Dong, Y.; Li, M.; Yang, L.; Lu, W. Quercetin Protects Against Lipopolysaccharide-Induced Intestinal Oxidative Stress in Broiler Chickens through Activation of Nrf2 Pathway. Molecules 2020, 25, 1053.
- Vanani, A.R.; Mahdavinia, M.; Shirani, M.; Alizadeh, S.; Dehghani, M.A. Protective effects of quercetin against oxidative stress induced by bisphenol-A in rat cardiac mitochondria. Environ. Sci. Pollut. Res. Int. 2020, 27, 15093–15102.
- da Silva Araujo, N.P.; de Matos, N.A.; Leticia Antunes Mota, S.; Farias de Souza, A.B.; Dantas Cangussu, S.; Cunha Alvim de Menezes, R.; Silva Bezerra, F. Quercetin Attenuates Acute Lung Injury Caused by Cigarette Smoke Both In Vitro and In Vivo. COPD 2020, 17, 205–214.
- Hu, T.; Shi, J.J.; Fang, J.; Wang, Q.; Chen, Y.B.; Zhang, S.J. Quercetin ameliorates diabetic encephalopathy through SIRT1/ER stress pathway in db/db mice. Aging (Albany NY) 2020, 12, 7015–7029.
- Cacciotti, C.; Fleming, A.; Ramaswamy, V. Advances in the molecular classification of pediatric brain tumors: A guide to the galaxy. J. Pathol. 2020, 251, 249–261.
- Udaka, Y.T.; Packer, R.J. Pediatric Brain Tumors. Neurol. Clin. 2018, 36, 533–556.
- Tan, A.C.; Ashley, D.M.; Lopez, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312.
- Darband, S.G.; Kaviani, M.; Yousefi, B.; Sadighparvar, S.; Pakdel, F.G.; Attari, J.A.; Mohebbi, I.; Naderi, S.; Majidinia, M. Quercetin: A functional dietary flavonoid with potential chemo-preventive properties in colorectal cancer. J. Cell Physiol. 2018, 233, 6544–6560.
- Sharmila, G.; Bhat, F.A.; Arunkumar, R.; Elumalai, P.; Raja Singh, P.; Senthilkumar, K.; Arunakaran, J. Chemopreventive effect of quercetin, a natural dietary flavonoid on prostate cancer in in vivo model. Clin. Nutr. 2014, 33, 718–726.
- Davoodvandi, A.; Shabani Varkani, M.; Clark, C.C.T.; Jafarnejad, S. Quercetin as an anticancer agent: Focus on esophageal cancer. J. Food Biochem. 2020, 44, e13374.
- Harris, Z.; Donovan, M.G.; Branco, G.M.; Limesand, K.H.; Burd, R. Quercetin as an Emerging Anti-Melanoma Agent: A Four-Focus Area Therapeutic Development Strategy. Front. Nutr. 2016, 3, 48.
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174.
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130.
- American Psychological Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Publishing, Inc.: Washington, DC, USA, 2013.
- Pan, P.Y.; Bolte, S.; Kaur, P.; Jamil, S.; Jonsson, U. Neurological disorders in autism: A systematic review and meta-analysis. Autism 2020, 1362361320951370.
- Lyall, K.; Croen, L.; Daniels, J.; Fallin, M.D.; Ladd-Acosta, C.; Lee, B.K.; Park, B.Y.; Snyder, N.W.; Schendel, D.; Volk, H.; et al. The Changing Epidemiology of Autism Spectrum Disorders. Annu. Rev. Public Health 2017, 38, 81–102.
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental risk factors for autism: An evidence-based review of systematic reviews and meta-analyses. Mol. Autism 2017, 8, 13.
- Ijomone, O.M.; Olung, N.F.; Akingbade, G.T.; Okoh, C.O.A.; Aschner, M. Environmental influence on neurodevelopmental disorders: Potential association of heavy metal exposure and autism. J. Trace Elem. Med. Biol. 2020, 62, 126638.
- Cheroni, C.; Caporale, N.; Testa, G. Autism spectrum disorder at the crossroad between genes and environment: Contributions, convergences, and interactions in ASD developmental pathophysiology. Mol. Autism 2020, 11, 69.
- Yousaf, A.; Waltes, R.; Haslinger, D.; Klauck, S.M.; Duketis, E.; Sachse, M.; Voran, A.; Biscaldi, M.; Schulte-Ruther, M.; Cichon, S.; et al. Quantitative genome-wide association study of six phenotypic subdomains identifies novel genome-wide significant variants in autism spectrum disorder. Transl. Psychiatry 2020, 10, 215.
- Lamy, M.; Pedapati, E.V.; Dominick, K.L.; Wink, L.K.; Erickson, C.A. Recent Advances in the Pharmacological Management of Behavioral Disturbances Associated with Autism Spectrum Disorder in Children and Adolescents. Paediatr. Drugs 2020.
- Hurwitz, R.; Blackmore, R.; Hazell, P.; Williams, K.; Woolfenden, S. Tricyclic antidepressants for autism spectrum disorders (ASD) in children and adolescents. Cochrane Database Syst. Rev. 2012.
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current Enlightenment About Etiology and Pharmacological Treatment of Autism Spectrum Disorder. Front. Neurosci. 2018, 12, 304.
- Whitehouse, A.J. Complementary and alternative medicine for autism spectrum disorders: Rationale, safety and efficacy. J. Paediatr. Child. Health 2013, 49, E438–E442.
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. Polyphenols as food bioactive compounds in the context of Autism Spectrum Disorders: A critical mini-review. Neurosci. Biobehav. Rev. 2019, 102, 290–298.
- Osredkar, J.; Gosar, D.; Macek, J.; Kumer, K.; Fabjan, T.; Finderle, P.; Sterpin, S.; Zupan, M.; Jekovec Vrhovsek, M. Urinary Markers of Oxidative Stress in Children with Autism Spectrum Disorder (ASD). Antioxidants 2019, 8, 187.
- Carpita, B.; Muti, D.; Dell’Osso, L. Oxidative Stress, Maternal Diabetes, and Autism Spectrum Disorders. Oxid Med. Cell Longev. 2018, 2018, 3717215.
- Theoharides, T.C.; Asadi, S.; Panagiotidou, S. A case series of a luteolin formulation (NeuroProtek(R)) in children with autism spectrum disorders. Int. J. Immunopathol. Pharm. 2012, 25, 317–323.
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clin. Ther. 2013, 35, 592–602.
- Tanaka, T.; Masubuchi, Y.; Okada, R.; Nakajima, K.; Nakamura, K.; Masuda, S.; Nakahara, J.; Maronpot, R.R.; Yoshida, T.; Koyanagi, M.; et al. Ameliorating effect of postweaning exposure to antioxidant on disruption of hippocampal neurogenesis induced by developmental hypothyroidism in rats. J. Toxicol. Sci. 2019, 44, 357–372.
- de Mattos, B.D.S.; Soares, M.S.P.; Spohr, L.; Pedra, N.S.; Teixeira, F.C.; de Souza, A.A.; Stefanello, F.M.; Baldissarelli, J.; Gamaro, G.D.; Spanevello, R.M. Quercetin prevents alterations of behavioral parameters, delta-aminolevulinic dehydratase activity, and oxidative damage in brain of rats in a prenatal model of autism. Int. J. Dev. Neurosci. 2020.
- Ersoz, M.; Erdemir, A.; Derman, S.; Arasoglu, T.; Mansuroglu, B. Quercetin-loaded nanoparticles enhance cytotoxicity and antioxidant activity on C6 glioma cells. Pharm. Dev. Technol. 2020, 25, 757–766.
- Cholia, R.P.; Kumari, S.; Kumar, S.; Kaur, M.; Kumar, R.; Dhiman, M.; Mantha, A.K. An in vitro study ascertaining the role of H2O2 and glucose oxidase in modulation of antioxidant potential and cancer cell survival mechanisms in glioblastoma U-87 MG cells. Metab. Brain Dis. 2017, 32, 1705–1716.
- Klauser, E.; Gulden, M.; Maser, E.; Seibert, S.; Seibert, H. Additivity, antagonism, and synergy in arsenic trioxide-induced growth inhibition of C6 glioma cells: Effects of genistein, quercetin and buthionine-sulfoximine. Food Chem. Toxicol. 2014, 67, 212–221.
- Jang, E.; Kim, I.Y.; Kim, H.; Lee, D.M.; Seo, D.Y.; Lee, J.A.; Choi, K.S.; Kim, E. Quercetin and chloroquine synergistically kill glioma cells by inducing organelle stress and disrupting Ca(2+) homeostasis. Biochem. Pharm. 2020, 178, 114098.
- Wang, G.; Wang, J.J.; Yang, G.Y.; Du, S.M.; Zeng, N.; Li, D.S.; Li, R.M.; Chen, J.Y.; Feng, J.B.; Yuan, S.H.; et al. Effects of quercetin nanoliposomes on C6 glioma cells through induction of type III programmed cell death. Int. J. Nanomed. 2012, 7, 271–280.
- Wang, G.; Wang, J.; Luo, J.; Wang, L.; Chen, X.; Zhang, L.; Jiang, S. PEG2000-DPSE-coated quercetin nanoparticles remarkably enhanced anticancer effects through induced programed cell death on C6 glioma cells. J. Biomed. Mater. Res. A 2013, 101, 3076–3085.
- Kim, H.; Moon, J.Y.; Ahn, K.S.; Cho, S.K. Quercetin induces mitochondrial mediated apoptosis and protective autophagy in human glioblastoma U373MG cells. Oxid. Med. Cell Longev. 2013, 2013, 596496.
- Taylor, M.A.; Khathayer, F.; Ray, S.K. Quercetin and Sodium Butyrate Synergistically Increase Apoptosis in Rat C6 and Human T98G Glioblastoma Cells Through Inhibition of Autophagy. Neurochem. Res. 2019, 44, 1715–1725.
- Michaud-Levesque, J.; Bousquet-Gagnon, N.; Beliveau, R. Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp. Cell Res. 2012, 318, 925–935.
- Jakubowicz-Gil, J.; Langner, E.; Badziul, D.; Wertel, I.; Rzeski, W. Apoptosis induction in human glioblastoma multiforme T98G cells upon temozolomide and quercetin treatment. Tumour. Biol. 2013, 34, 2367–2378.
- Sang, D.P.; Li, R.J.; Lan, Q. Quercetin sensitizes human glioblastoma cells to temozolomide in vitro via inhibition of Hsp27. Acta Pharm. Sin. 2014, 35, 832–838.
- Badziul, D.; Jakubowicz-Gil, J.; Langner, E.; Rzeski, W.; Glowniak, K.; Gawron, A. The effect of quercetin and imperatorin on programmed cell death induction in T98G cells in vitro. Pharm. Rep. 2014, 66, 292–300.
- Jakubowicz-Gil, J.; Langner, E.; Badziul, D.; Wertel, I.; Rzeski, W. Quercetin and sorafenib as a novel and effective couple in programmed cell death induction in human gliomas. Neurotox Res. 2014, 26, 64–77.
- Zamin, L.L.; Filippi-Chiela, E.C.; Vargas, J.; Demartini, D.R.; Meurer, L.; Souza, A.P.; Bonorino, C.; Salbego, C.; Lenz, G. Quercetin promotes glioma growth in a rat model. Food Chem. Toxicol. 2014, 63, 205–211.
- Lagerweij, T.; Hiddingh, L.; Biesmans, D.; Crommentuijn, M.H.; Cloos, J.; Li, X.N.; Kogiso, M.; Tannous, B.A.; Vandertop, W.P.; Noske, D.P.; et al. A chemical screen for medulloblastoma identifies quercetin as a putative radiosensitizer. Oncotarget 2016, 7, 35776–35788.
- Labbe, D.; Provencal, M.; Lamy, S.; Boivin, D.; Gingras, D.; Beliveau, R. The flavonols quercetin, kaempferol, and myricetin inhibit hepatocyte growth factor-induced medulloblastoma cell migration. J. Nutr. 2009, 139, 646–652.
- Akhondzadeh, S.; Mohammadi, M.-R.; Momeni, F. Passiflora incarnata in the teartment of attention-deficit hyperactivity disorder in children and adolescents. Therapy 2005, 2, 609–614.
- Weber, W.; Vander Stoep, A.; McCarty, R.L.; Weiss, N.S.; Biederman, J.; McClellan, J. Hypericum perforatum (St John’s wort) for attention-deficit/hyperactivity disorder in children and adolescents: A randomized controlled trial. JAMA 2008, 299, 2633–2641.
- Niederhofer, H., St. John’s wort may improve some symptoms of attention-deficit hyperactivity disorder. Nat. Prod. Res. 2010, 24, 203–205.
- Schmidt, A.J.; Krieg, J.C.; Hemmeter, U.M.; Kircher, T.; Schulz, E.; Clement, H.W.; Heiser, P. Impact of plant extracts tested in attention-deficit/hyperactivity disorder treatment on cell survival and energy metabolism in human neuroblastoma SH-SY5Y cells. Phytother. Res. 2010, 24, 1549–1553.
- Romero, M.; Jimenez, R.; Hurtado, B.; Moreno, J.M.; Rodriguez-Gomez, I.; Lopez-Sepulveda, R.; Zarzuelo, A.; Perez-Vizcaino, F.; Tamargo, J.; Vargas, F.; et al. Lack of beneficial metabolic effects of quercetin in adult spontaneously hypertensive rats. Eur. J. Pharm. 2010, 627, 242–250.
- Monteiro, M.M.; Franca-Silva, M.S.; Alves, N.F.; Porpino, S.K.; Braga, V.A. Quercetin improves baroreflex sensitivity in spontaneously hypertensive rats. Molecules 2012, 17, 12997–13008.
- Karuppagounder, S.S.; Madathil, S.K.; Pandey, M.; Haobam, R.; Rajamma, U.; Mohanakumar, K.P. Quercetin up-regulates mitochondrial complex-I activity to protect against programmed cell death in rotenone model of Parkinson’s disease in rats. Neuroscience 2013, 236, 136–148.
- Yan, L.; Zhang, J.D.; Wang, B.; Lv, Y.J.; Jiang, H.; Liu, G.L.; Qiao, Y.; Ren, M.; Guo, X.F. Quercetin inhibits left ventricular hypertrophy in spontaneously hypertensive rats and inhibits angiotensin II-induced H9C2 cells hypertrophy by enhancing PPAR-gamma expression and suppressing AP-1 activity. PLoS ONE 2013, 8, e72548.
- Kanazawa, L.K.; Vecchia, D.D.; Wendler, E.M.; Hocayen, P.A.; Beirao, P.S., Jr.; de Melo, M.L.; Dos Reis Livero, F.A.; Corso, C.R.; Stipp, M.C.; Acco, A.; et al. Effects of acute and chronic quercetin administration on methylphenidate-induced hyperlocomotion and oxidative stress. Life Sci. 2017, 171, 1–8.
- Polanczyk, G.; de Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The worldwide prevalence of ADHD: A systematic review and metaregression analysis. Am. J. Psychiatry 2007, 164, 942–948.
- Faraone, S.V.; Asherson, P.; Banaschewski, T.; Biederman, J.; Buitelaar, J.K.; Ramos-Quiroga, J.A.; Rohde, L.A.; Sonuga-Barke, E.J.; Tannock, R.; Franke, B. Attention-deficit/hyperactivity disorder. Nat. Rev. Dis Primers 2015, 1, 15020.
- Sayal, K.; Prasad, V.; Daley, D.; Ford, T.; Coghill, D. ADHD in children and young people: Prevalence, care pathways, and service provision. Lancet Psychiatry 2018, 5, 175–186.
- Alvarez-Arellano, L.; Gonzalez-Garcia, N.; Salazar-Garcia, M.; Corona, J.C. Antioxidants as a Potential Target against Inflammation and Oxidative Stress in Attention-Deficit/Hyperactivity Disorder. Antioxidants 2020, 9, 176.
- Wilens, T.E.; Faraone, S.V.; Biederman, J. Attention-deficit/hyperactivity disorder in adults. JAMA 2004, 292, 619–623.
- Wolraich, M.L.; Chan, E.; Froehlich, T.; Lynch, R.L.; Bax, A.; Redwine, S.T.; Ihyembe, D.; Hagan, J.F., Jr. ADHD Diagnosis and Treatment Guidelines: A Historical Perspective. Pediatrics 2019, 144, e20191682.
- Briars, L.; Todd, T. A Review of Pharmacological Management of Attention-Deficit/Hyperactivity Disorder. J. Pediatr. Pharm. 2016, 21, 192–206.
- Koda, K.; Ago, Y.; Cong, Y.; Kita, Y.; Takuma, K.; Matsuda, T. Effects of acute and chronic administration of atomoxetine and methylphenidate on extracellular levels of noradrenaline, dopamine and serotonin in the prefrontal cortex and striatum of mice. J. Neurochem. 2010, 114, 259–270.
- Bymaster, F.P.; Katner, J.S.; Nelson, D.L.; Hemrick-Luecke, S.K.; Threlkeld, P.G.; Heiligenstein, J.H.; Morin, S.M.; Gehlert, D.R.; Perry, K.W. Atomoxetine increases extracellular levels of norepinephrine and dopamine in prefrontal cortex of rat: A potential mechanism for efficacy in attention deficit/hyperactivity disorder. Neuropsychopharmacology 2002, 27, 699–711.
- Swanson, C.J.; Perry, K.W.; Koch-Krueger, S.; Katner, J.; Svensson, K.A.; Bymaster, F.P. Effect of the attention deficit/hyperactivity disorder drug atomoxetine on extracellular concentrations of norepinephrine and dopamine in several brain regions of the rat. Neuropharmacology 2006, 50, 755–760.
- Clemow, D.B. Misuse of Methylphenidate. Curr. Top. Behav. Neurosci. 2017, 34, 99–124.
- Reed, V.A.; Buitelaar, J.K.; Anand, E.; Day, K.A.; Treuer, T.; Upadhyaya, H.P.; Coghill, D.R.; Kryzhanovskaya, L.A.; Savill, N.C. The Safety of Atomoxetine for the Treatment of Children and Adolescents with Attention-Deficit/Hyperactivity Disorder: A Comprehensive Review of Over a Decade of Research. CNS Drugs 2016, 30, 603–628.
- Lopresti, A.L. Oxidative and nitrosative stress in ADHD: Possible causes and the potential of antioxidant-targeted therapies. Atten. Defic. Hyperact. Disord. 2015, 7, 237–247.
- Joseph, N.; Zhang-James, Y.; Perl, A.; Faraone, S.V. Oxidative Stress and ADHD: A Meta-Analysis. J. Attent. Disord. 2015, 19, 915–924.
- Corona, J.C. Role of Oxidative Stress and Neuroinflammation in Attention-Deficit/Hyperactivity Disorder. Antioxidants 2020, 9, 1039.
- Motaghinejad, M.; Motevalian, M.; Shabab, B.; Fatima, S. Effects of acute doses of methylphenidate on inflammation and oxidative stress in isolated hippocampus and cerebral cortex of adult rats. J. Neural. Transm. (Vienna) 2017, 124, 121–131.
- Meneses, A.; Perez-Garcia, G.; Ponce-Lopez, T.; Tellez, R.; Gallegos-Cari, A.; Castillo, C. Spontaneously hypertensive rat (SHR) as an animal model for ADHD: A short overview. Rev. Neurosci. 2011, 22, 365–371.
- Corona, J.C.; Duchen, M.R. PPARgamma as a therapeutic target to rescue mitochondrial function in neurological disease. Free Radic. Biol. Med. 2016, 100, 153–163.
- Zhang, Y.; Wang, S.X.; Ma, J.W.; Li, H.Y.; Ye, J.C.; Xie, S.M.; Du, B.; Zhong, X.Y. EGCG inhibits properties of glioma stem-like cells and synergizes with temozolomide through downregulation of P-glycoprotein inhibition. J. Neurooncol. 2015, 121, 41–52.
- Pilorget, A.; Berthet, V.; Luis, J.; Moghrabi, A.; Annabi, B.; Beliveau, R. Medulloblastoma cell invasion is inhibited by green tea (-)epigallocatechin-3-gallate. J. Cell Biochem. 2003, 90, 745–755.
- Jia, W.Z.; Zhao, J.C.; Sun, X.L.; Yao, Z.G.; Wu, H.L.; Xi, Z.Q. Additive anticancer effects of chrysin and low dose cisplatin in human malignant glioma cell (U87) proliferation and evaluation of the mechanistic pathway. J. BUON 2015, 20, 1327–1336.
- You, Y.; Wang, R.; Shao, N.; Zhi, F.; Yang, Y. Luteolin suppresses tumor proliferation through inducing apoptosis and autophagy via MAPK activation in glioma. ONCO Targets 2019, 12, 2383–2396.
- Anson, D.M.; Wilcox, R.M.; Huseman, E.D.; Stump, T.A.; Paris, R.L.; Darkwah, B.O.; Lin, S.; Adegoke, A.O.; Gryka, R.J.; Jean-Louis, D.S.; et al. Luteolin Decreases Epidermal Growth Factor Receptor-Mediated Cell Proliferation and Induces Apoptosis in Glioblastoma Cell Lines. Basic Clin. Pharm. Toxicol. 2018, 123, 678–686.
- Wang, Q.; Wang, H.; Jia, Y.; Pan, H.; Ding, H. Luteolin induces apoptosis by ROS/ER stress and mitochondrial dysfunction in gliomablastoma. Cancer Chemother. Pharm. 2017, 79, 1031–1041.
- Chakrabarti, M.; Ray, S.K. Synergistic anti-tumor actions of luteolin and silibinin prevented cell migration and invasion and induced apoptosis in glioblastoma SNB19 cells and glioblastoma stem cells. Brain Res. 2015, 1629, 85–93.
- Chakrabarti, M.; Ray, S.K. Anti-tumor activities of luteolin and silibinin in glioblastoma cells: Overexpression of miR-7-1-3p augmented luteolin and silibinin to inhibit autophagy and induce apoptosis in glioblastoma in vivo. Apoptosis 2016, 21, 312–328.
- Cabrera, C.; Artacho, R.; Gimenez, R. Beneficial effects of green tea—A review. J. Am. Coll Nutr. 2006, 25, 79–99.
- Banji, D.; Banji, O.J.; Abbagoni, S.; Hayath, M.S.; Kambam, S.; Chiluka, V.L. Amelioration of behavioral aberrations and oxidative markers by green tea extract in valproate induced autism in animals. Brain Res. 2011, 1410, 141–151.
- Bertolino, B.; Crupi, R.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Beneficial Effects of Co-Ultramicronized Palmitoylethanolamide/Luteolin in a Mouse Model of Autism and in a Case Report of Autism. CNS Neurosci. 2017, 23, 87–98.
- Kumaravel, P.; Melchias, G.; Vasanth, N.; Manivasagam, T. Epigallocatechin Gallate Attenuates Behavioral Defects in Sodium Valproate Induced Autism Rat Model. Res. J. Pharm. Tech. 2017, 10, 1477–1480.
- Bhandari, R.; Paliwal, J.K.; Kuhad, A. Naringenin and its nanocarriers as potential phytotherapy for autism spectrum disorders. J. Funct. Foods 2018, 47, 361–375.
- Niederhofer, H. First preliminary results of an observation of Ginkgo Biloba treating patients with autistic disorder. Phytother. Res. 2009, 23, 1645–1646.
- Hasanzadeh, E.; Mohammadi, M.R.; Ghanizadeh, A.; Rezazadeh, S.A.; Tabrizi, M.; Rezaei, F.; Akhondzadeh, S. A double-blind placebo controlled trial of Ginkgo biloba added to risperidone in patients with autistic disorders. Child. Psychiatry Hum. Dev. 2012, 43, 674–682.
- Zhou, R.Y.; Wang, J.J.; You, Y.; Sun, J.C.; Song, Y.C.; Yuan, H.X.; Han, X.M. Effect of baicalin on ATPase and LDH and its regulatory effect on the AC/cAMP/PKA signaling pathway in rats with attention deficit hyperactivity disorder. Zhongguo Dang Dai Er Ke Za Zhi 2017, 19, 576–582.
- Zhou, R.Y.; Han, X.M.; Wang, J.J.; Yuan, H.X.; Sun, J.C.; You, Y.; Song, Y.C. Effect of baicalin on behavioral characteristics of rats with attention deficit hyperactivity disorder. Zhongguo Dang Dai Er Ke Za Zhi 2017, 19, 930–937.
- Chovanova, Z.; Muchova, J.; Sivonova, M.; Dvorakova, M.; Zitnanova, I.; Waczulikova, I.; Trebaticka, J.; Skodacek, I.; Durackova, Z. Effect of polyphenolic extract, Pycnogenol, on the level of 8-oxoguanine in children suffering from attention deficit/hyperactivity disorder. Free Radic Res. 2006, 40, 1003–1010.
- Dvorakova, M.; Sivonova, M.; Trebaticka, J.; Skodacek, I.; Waczulikova, I.; Muchova, J.; Durackova, Z. The effect of polyphenolic extract from pine bark, Pycnogenol on the level of glutathione in children suffering from attention deficit hyperactivity disorder (ADHD). Redox Rep. 2006, 11, 163–172.
- Trebaticka, J.; Kopasova, S.; Hradecna, Z.; Cinovsky, K.; Skodacek, I.; Suba, J.; Muchova, J.; Zitnanova, I.; Waczulikova, I.; Rohdewald, P.; et al. Treatment of ADHD with French maritime pine bark extract, Pycnogenol. Eur. Child. Adolesc. Psychiatry 2006, 15, 329–335.
- Yoon, S.Y.; dela Pena, I.; Kim, S.M.; Woo, T.S.; Shin, C.Y.; Son, K.H.; Park, H.; Lee, Y.S.; Ryu, J.H.; Jin, M.; et al. Oroxylin A improves attention deficit hyperactivity disorder-like behaviors in the spontaneously hypertensive rat and inhibits reuptake of dopamine in vitro. Arch. Pharm. Res. 2013, 36, 134–140.
- dela Pena, I.C.; Young Yoon, S.; Kim, Y.; Park, H.; Man Kim, K.; Hoon Ryu, J.; Young Shin, C.; Hoon Cheong, J. 5,7-Dihydroxy-6-methoxy-4′-phenoxyflavone, a derivative of oroxylin A improves attention-deficit/hyperactivity disorder (ADHD)-like behaviors in spontaneously hypertensive rats. Eur. J. Pharm. 2013, 715, 337–344.


