 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Fulvio Massaro | + 3242 word(s) | 3242 | 2021-01-07 02:14:36 | | | |
| 2 | Vivi Li | Meta information modification | 3242 | 2021-01-13 11:05:17 | | |
Video Upload Options
Aging of bone marrow is a complex process that is involved in the development of many diseases, including hematologic cancers. The results obtained in this field of research, year after year, underline the important role of cross-talk between hematopoietic stem cells and their close environment. In bone marrow, mesenchymal stromal cells (MSCs) are a major player in cell-to-cell communication, presenting a wide range of functionalities, sometimes opposite, depending on the environmental conditions. Although these cells are actively studied for their therapeutic properties, their role in tumor progression remains unclear. One of the reasons for this is that the aging of MSCs has a direct impact on their behavior and on hematopoiesis. In addition, tumor progression is accompanied by dynamic remodeling of the bone marrow niche that may interfere with MSC functions.
1. Introduction
Cancers and aging are closely linked [1]. Indeed, in most organisms, aging is accompanied by multiple alterations at the cellular, tissue and systemic levels. All of these alterations provide fertile ground for the development and progression of tumors, as evidenced by the shared hallmarks of aging and cancers [2][3]. Although the intrinsic processes leading to cell transformation from a normal cell into a tumor cell are now well known, it is also commonly accepted that the microenvironment surrounding cells and the interactions between malignant cells and this microenvironment play crucial roles in tumor development and growth. Malignant hemopathies represent no exception: they comprise a wide collection of disorders, all originating from cells of the bone marrow (BM) and the lymphatic system and accounting for almost 230,000 new cases every year in Europe [4].
The homeostasis and maintenance of BM cells and the immune system require continuous renewal of all types of blood cells. This function is ensured in the BM by hematopoietic stem cells (HSCs) that can differentiate into myeloid progenitors, giving rise to erythrocytes, platelets, granulocytes, and monocytes, or into lymphoid progenitors, giving rise to B lymphocytes, T lymphocytes and NK cells. The function and regulation of HSCs are supported by their close environment, the BM niche [5][6][7]. A growing number of studies point to a clear link between aging, remodeling of the BM microenvironment and impairment of hematopoiesis, leading, among other things, to hematologic cancers [8][9]. The BM niche is a complex and dynamic network that is not yet fully understood and is regulated by a wide number of cell types: endothelial cells, mesenchymal stromal cells (MSCs), perivascular stromal cells, osteoblasts, sympathetic neurons, nonmyelinating Schwann cells, adipocytes and regulatory T cells.
MSCs are multipotent nonhematopoietic cells able to differentiate into osteoblasts, chondrocytes, adipocytes and fibroblasts [10][11][12]. They also secrete a wide variety of compounds, such as growth factors, antiapoptotic factors, angiogenic factors and several cytokines, and thus contribute to the regenerative process, wound healing, hematopoietic support and regulation of the immune response [13][14]. MSCs also produce a large amount of extracellular vesicles (EVs), small vesicles playing a major role in cell-to-cell communication. EVs transport different elements, such as proteins, lipids and microRNAs (miRNAs), to target cells and are involved in many biological functions of MSCs [15][16][17]. It is now known that the aging of MSCs alters their EV production and has a direct impact on their functions and differentiation capacities [18][19]. Aging is also associated with an increased incidence of hematologic malignancies such as chronic and acute leukemias, non-Hodgkin lymphomas and plasma cell disorders: the mean age at diagnosis is 65–70 years, and the incidence typically increases in groups of older subjects [4][20]. Due to their functions, MSCs are important actors in the tumoral microenvironment, but their exact role remains ambiguous. Indeed, different studies carried out to date show both a protumoral and an antitumoral function of MSCs, as reviewed by Galland and Stamenkovic in The Journal of Pathology [21]. Although the prominent role of MSCs in vivo seems to be participation in tumor progression, further studies will be necessary to obtain a deep understanding of their exact role inside the tumoral microenvironment.
2. The Role of BM-MSCs in BM and Hematopoiesis Alterations during Aging
As mentioned above, during the aging process, changes in HSCs and hematopoiesis disturbances occur. More precisely, the number of lymphoid progenitors decreases to the benefit of myeloid progenitors that increase but lose some of their functions [21][22]. The result is an alteration of the immune system, leading to an increased susceptibility to infections and to the development of autoimmune diseases and cancers. The aging of HSCs is due in part to cell-intrinsic factors, as reviewed by Mejia-Ramirez and Florian in Haematologica [23], but also by external signals from the aging microenvironment of which BM-MSCs are a part. In this section, we will discuss the main features of BM-MSC aging and its consequences on hematopoiesis and the inflammatory state of BM through the modifications of BM-MSC secretome, the imbalance of their immunomodulation properties and the imbalance between osteogenesis and adipogenesis leading to progressive replacement of bone by fat (Figure 1). We will also briefly examine the spatial and functional heterogeneity of BM-MSCs inside the BM niche and its change during aging.
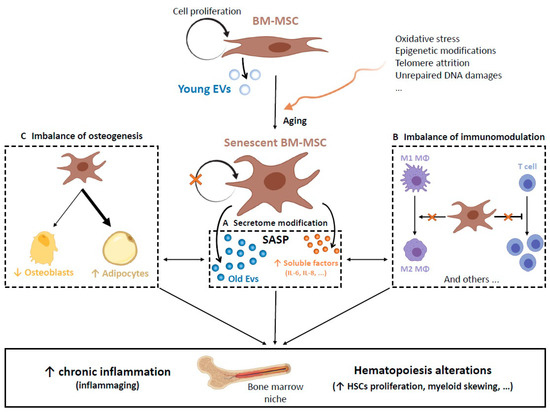
Figure 1. With aging, several factors induce the senescence of bone marrow mesenchymal stromal cells (BM-MSCs) that accumulate inside the bone marrow (BM) niche. (A) The senescent BM-MSCs adopt the senescent-associated secretory phenotype (SASP) enriched in particular with proinflammatory cytokines, and their extracellular vesicle (EV) morphology and content are strongly modified (see Section 2.1). (B) The immunomodulatory properties of aged BM-MSCs are impaired. For example, their ability to promote macrophage (MΦ) polarization into the M2 phenotype and their capacity to inhibit T lymphocyte proliferation are reduced (see Section 2.2). (C) An imbalance between osteogenesis and adipogenesis occurs, leading to a progressive replacement of bone by fat (see Section 2.3). All of these processes are closely interconnected and can lead to the establishment of low-grade chronic inflammation and hematopoiesis alterations (Figure created with BioRender).
2.1. Epigenetic and Secretome Modifications Associated with BM-MSC Aging
MSCs are multipotent cells with proliferative properties. However, similar to any normal cell, they can only undergo a limited number of cell divisions before entering a senescent state. Cellular senescence and its related cell cycle arrest were observed for the first time by Hayflick in long-term in vitro culture of human fibroblasts [24]. Since then, a wide variety of factors causing MSC senescence have also been described, such as oxidative stress [25], telomere attrition occurring during in vitro expansion [26] or unrepaired DNA damages [27]. Accumulation of senescent cells was also observed in several aged tissues, as it was well illustrated in a recent study evaluating the expression of p16 and p21, two markers of senescence, in organs from young or old donors [28]. An increased level of p21 was also observed in BM-MSCs from elderly people, suggesting that senescent BM-MSCs accumulate with physiological aging [29]. Nevertheless, some experiments studying MSC senescence do not use cells form elderly donors but rather in vitro stress-induced-senescence conditions such as long-term culture expansion or senescence induced by gamma irradiation. It is therefore necessary to remain cautious when comparing data concerning in vitro senescence with physiological aging.
Several pathways and actors implicated in MSC senescent cell cycle arrest have been identified: the well-established p53/p21 and p16/pRB pathways, as well as the AKT/mTOR pathway [30], JAK/STAT pathway [31], mitogen activated protein kinase p38MAP [32] and fibroblast growth factor FGF21 [33].
Senescence is accompanied by many cellular modifications at morphological and functional levels: the cells become larger and resistant to apoptosis, while the autophagy process decreases, and they are also subject to genetic and epigenetic modifications [34][35]. Epigenetic modifications are key components of the BM niche homeostasis and can contribute to age- and disease-associated MSC alterations. Modifications of MSC DNA methylation patterns and hypermethylated and hypomethylated CpG sites in several genomic loci have been observed upon aging and replicative senescence [36][37]. Some epigenetic regulators have been identified to participate in MSCs aging. The expression and activation of Sirt1, a NAD-dependent histone deacetylase, decrease with age and modify MSC proliferation and differentiation [38][39]. Interestingly, miR-199b-5p, which is predicted to target Sirt1, is deregulated in old BM-MSCs [40]. MSCs deficient in Sirt6, another histone deacetylase, displayed accelerated cellular senescence, dysregulated redox metabolism and increased sensitivity to oxidative stress [41]. In HSCs, identification of somatic mutations in the epigenetic regulators DNMT3, TET2 and ASXL1 is associated with an increased risk of developing hematologic cancers [42]. These mutations can occur as people age and their identification in healthy people is known as clonal hematopoiesis of indeterminate potential (CHIP). These three epigenetic regulators seem to be involved in MSC aging. In human umbilical cord blood-derived MSCs (UC-MSCs), the inhibition of DNMT1 and DNMT3b induces cellular senescence [43]. In a mouse model used to study age-related skeletal diseases, the expression of TET2 resulted decreased [44]. In addition, it has been shown in mice that a loss of ASXL1 or TET2 impairs BM-MSCs fate and their ability to support hematopoiesis [45][46]. Taken together, these two observations suggest that epigenetic modifications of BM-MSCs occurring during aging can contribute to hematopoiesis disturbances.
The alterations associated with BM-MSC senescence also lead to a deep modification of their secretome, making them adopt a new phenotype called the senescent-associated secretory phenotype (SASP) [34] (Figure 1A). This SASP is characterized by increased secretion of growth factors, proangiogenic factors, extracellular matrix remodeling factors and especially proinflammatory cytokines such as IL-1β, IL-6 and IL-8 [47][48][49][50][51]. It is now well known that EVs contribute greatly to the SASP and that senescence of MSCs has a strong impact on them: their secretion is increased while their size is reduced and their content is modified, especially in terms of miRNA [18][52]. For example, the activation of AKT in aged BM-MSCs leads to increased partitioning of miR-17 and miR-34a in EVs, which, upon transfer to HSCs, cause functional impairment via downregulation of autophagy-related genes [53]. Terlecki-Zaniewicz and colleagues suggested that EVs of dermal fibroblasts and their miRNAs act as cargo for novel members of the SASP that are selectively secreted or retained in cellular senescence [54]. Although there are no similar experimental data on MSCs, it is reasonable to assume this may also be applicable to them. Robbins suggested that senescent cell-derived EVs could function as pro-geronic factors [55]. The SASP participates in the establishment of the low-grade and chronic inflammation state observed during aging, called inflammaging [56][57].
In addition, proinflammatory cytokines induce the expression of other cytokines by BM-MSCs [58]. It has been reported that BM-MSCs secrete a significant amount of IL-6 in response to TNF-α and IFN-γ [59] and that this cytokine is secreted at higher levels by aged BM-MSCs [50]. IL-6 is a proinflammatory cytokine implicated in inflammaging or promoting tumor growth and metastasis formation. The serum of elderly patients with chronic disease or cancer usually contains more IL-6 than the serum of young healthy people [60]. The MSCs contribute thus, by this cytokine release, to the “inflammaging” process well described by Fulop and others in age-related diseases [61]. With TGF-β, TNF-α and GM-CSF, IL-6 also promotes the differentiation of HSCs towards the myeloid lineage [50][62]. Interestingly, a recent analysis of the secretome of adipose tissue-derived MSCs (AT-MSCs) showed that GM-CSF is more highly secreted by senescent MSCs obtained by successive passaging [63]. Other proinflammatory cytokines secreted by BM-MSCs could impair hematopoiesis. Indeed, although IFN-γ promotes stem cell factor (SCF) expression in mouse BM-MSCs, an important factor for the support of hematopoiesis, chronic exposure of BM-MSCs to this cytokine leads to a decrease in the total number of BM-MSCs, diminishing their hematopoietic support [64]. In a recent paper, Gnani and colleagues showed a clonogenic impairment of young hematopoietic stem and progenitor cells (HSPCs) due to the activation of a proinflammatory program when they are exposed to compounds secreted by aged BM-MSCs [51].
2.2. Imbalance between Pro- and Anti-Inflammatory Functions
In addition to their role as multipotent progenitor cells, MSCs are also endogenous regulators of inflammation capable of immunosuppressive or proinflammatory functions depending on environmental conditions [65]. Thus, they express several Toll-like receptors (TLRs) whose activation influences their immunologic properties. In normal nonsenescent MSCs, the activation of TLR3 or the presence of IFN-γ and TNF-α polarizes the MSCs towards an anti-inflammatory state that is able to negatively regulate the proliferation of T lymphocytes and NK cells through the secretion of nitric oxide synthase (NOS) and prostaglandin E2 (PGE2), respectively [66]. MSCs also promote the polarization of macrophages into the alternatively activated anti-inflammatory type 2 state (M2) at the expense of the classical proinflammatory type 1 state (M1) [67].
These observations suggest that within the aging and inflamed microenvironment of the BM, BM-MSCs may influence other immune actors to counteract inflammation. However, the studies described in literature to date do not point in this direction. For example, it has been shown that gamma-irradiated senescent BM-MSCs showed a lower capacity to migrate in response to proinflammatory signals and, at least in part, a lower inhibitory capacity towards T lymphocytes [68]. In addition, the priming of UC-MSCs with IFN-γ and TNF-α induces the phosphorylation of p38MAP kinase only in aged MSCs, which could in turn negatively regulate the COX2/PGE2 pathway and explain at least partially the reduction in the immunomodulatory capacity of aged MSCs [69]. Other studies have highlighted the impact of MSC aging on macrophage polarization. In mice, indirect coculture of macrophages with BM-MSCs using transwells induces differential gene expression in macrophages depending on the aging state of BM-MSCs. Thus, TNF-α and iNOS, two markers of M1 proinflammatory macrophages, are upregulated after coculture with aged BM-MSCs compared to coculture with young BM-MSCs, which instead induce the expression of IL-10 and ARG1, two markers of the M2 state [70]. More recently, in a lung injury murine model, Huang and colleagues showed that injections of EVs secreted by young human BM-MSCs but not those secreted by the oldest BM-MSCs are able to polarize macrophages towards the M2 state, reducing the severity of lung injury [71]. These differences between old and young EVs might be due to their miRNA content.
Taken together, these results suggest that the aging of MSCs impairs their ability to adopt an immunosuppressive phenotype in response to environmental stimulation (Figure 1B). Interestingly, TLR signaling involved in MSC immune polarization is also implicated in the proliferation of HSCs towards the myeloid lineage and in the migration of monocytes [72][73].
2.3. Imbalance between Osteogenesis and Adipogenesis
Bone tissue is a dynamic tissue undergoing constant remodeling throughout its lifetime. Its homeostasis is maintained by two complementary processes: the formation of new bone by osteoblasts and the resorption of old and damaged tissues by osteoclasts. BM-MSCs play an important role in this balance by being recruited at the bone-resorptive site through TGF-β1 signaling and by differentiating into osteoblasts [74]. However, during aging, bone resorption increases, and the bone density of the organism progressively decreases, leading to osteoporosis and increasing the risk of fractures [75]. The age-related changes in MSC differentiation potential have been studied by several groups in mice and humans. Although conflicting results have been reported, one cause for the imbalance between bone and adipose tissue occurring with aging could be due to a gradual loss of the ability of BM-MSCs to differentiate into osteoblasts, favoring differentiation into adipocytes (Figure 1C). A study using senescent BM-MSCs obtained after long-term culture showed an increased osteogenic differentiation potential after several passages [76]. However, other studies comparing BM-MSCs harvested from young and old donors have shown both a maintenance [77][78] or a decrease in the osteogenic differentiation of oldest BM-MSCs [29][79][80][81]. In a recent study, authors analyzed the transcriptional profile of freshly isolated BM-MSCs from young and old donors and showed the upregulation of genes implicated in the peroxisome proliferator-activated receptor (PPAR) signaling in the oldest group, suggesting a reinforcement of pro-adipogenic microenvironment with aging [82].
Several factors are implicated in the control of BM-MSC differentiation: RUNX2 and SP7 promote osteogenesis [83][84], while CEBPα, CEBPβ, CEBPγ and PPARγ promote adipogenesis [85]. There is a growing body of data highlighting the age-dependent control of these factors. In mice, it has been shown that FOXP1, a transcription factor interacting with CEBPβ, is downregulated during aging [86]. Similarly, CBFβ and MAF, two cofactors of RUNX2, are also downregulated with increasing age [87][88], while PPARγ is upregulated [89]. All of these signaling pathway modifications promote adipogenesis. The miRNA content of BM-MSCs and their EVs also seem to be involved in the imbalance between osteogenesis and adipogenesis. Indeed, it has been shown that aging and oxidative stress can alter the miRNA cargo of EVs, which in turn causes the suppression of cellular proliferation and osteogenic differentiation of BM stromal cells [90]. It has also been reported that miR-31a-5p level rises in aged BM-MSCs and appears to be involved in increasing adipogenesis and decreasing osteogenesis [91]. The decrease in osteogenic differentiation by BM-MSCs is accompanied by a reduced level of osteopontin secretion, which is known to negatively regulate the self-renewal of HSCs [92][93].
Adipocytes in BM impair hematopoiesis by diminishing the differentiation of hematopoietic progenitors towards the B lymphocyte lineage [94]. In a recent paper, Aguilar-Navarro et al. observed an increase of adipocytes in BM of elderly people associated with an increase of maturing myeloid cells and they proposed a contributive role for adipocytes in myeloid skewing [95]. Another study conducted on mice has been shown that aging is associated with the expansion of adipogenic potential of a stem cell-like subpopulation in the BM which, in turn, altered hematopoiesis through an excessive production of Dipeptidyl peptidase-4 [96].
By increasing the number of adipocytes inside the BM, the aging of BM-MSCs could also indirectly impact the inflammatory state of the BM niche. It is indeed well known today that adipose tissue participates in the production of a large amount of soluble factors and cytokines and that aging and metabolic diseases, like obesity, are correlated with an increase of its proinflammatory cytokine secretion [97].
2.4. Functional and Spatial Heterogeneity of BM-MSCs
MSCs represent a complex cell population characterised by specific localisation and functional heterogeneity that may be essential to their biological role. Several surface markers can be used to identify the different subpopulations of MSCs [98]. In BM, CD271 antigen can be used to identify a subset of BM-MSCs able to inhibit the proliferation of allogenic T-lymphocytes and presenting lympho-hematopoietic engraftment-promoting properties [99]. Most of HSCs are located in intimate cell-cell contact with these CD271+ MSCs [100]. A low or negative expression of platelet derived growth factor receptor alpha (PDGRF-α) by CD271+ MSCs is correlated with expression of key-genes for HSC supportive function and this expression is modulated according to the different phases of development of the organism [101]. CD271+ cells can be further divided in two cell subgroups with different localisation depending on the expression of CD146. CD146+ status defines MSC population located in the perivascular spaces while CD146− cells are found in the endosteal region [102]. These populations have different degree of maturity: CD146− MSCs are more mature whereas CD146+ cells retain plasticity. Their distribution varies with age: Maijenburg et al. showed a predominance of CD146+ subset in pediatric and fetal BM and suggested that variation in MSC subpopulations is a dynamic process that can change MSC functions during aging of the BM [103].
Other studies using a new method of single cell transcriptional analysis showed age-related changes in BM-MSCs composition. Duscher et al. identified an age-related depletion of a subpopulation characterized by a pro-vascular transcriptional profile [104]. More recently, Khong and colleagues identified a unique quiescent subpopulation exclusively present in MSCs from young donors and showed that this subpopulation was characterized by a higher expression of genes involved in tissue regeneration [105].
It has also been described the existence of two populations of MSCs with neural crest or mesoderm embryonic origins and particularly the neural crest has been proposed as a source of MSCs with specialized hematopoietic stem cell niche function [106]. Embryonic origin has also been shown to play an essential role in the age-related decrease in the functional capacities of BM-MSCs [107].
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA A Cancer J. Clin. 2020, 70, 7–30.
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217.
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674.
- Rodriguez-Abreu, D.; Bordoni, A.; Zucca, E. Epidemiology of hematological malignancies. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2007, 18 (Suppl. 1), i3–i8.
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334.
- Taichman, R.S.; Emerson, S.G. Human osteoblasts support hematopoiesis through the production of granulocyte colony-stimulating factor. J. Exp. Med. 1994, 179, 1677–1682.
- Zhang, J.; Niu, C.; Ye, L.; Huang, H.; He, X.; Tong, W.G.; Ross, J.; Haug, J.; Johnson, T.; Feng, J.Q.; et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003, 425, 836–841.
- Geiger, H.; Denkinger, M.; Schirmbeck, R. Hematopoietic stem cell aging. Curr. Opin. Immunol. 2014, 29, 86–92.
- Hellmich, C.; Moore, J.A.; Bowles, K.M.; Rushworth, S.A. Bone Marrow Senescence and the Microenvironment of Hematological Malignancies. Front. Oncol. 2020, 10, 230.
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317.
- Direkze, N.C.; Hodivala-Dilke, K.; Jeffery, R.; Hunt, T.; Poulsom, R.; Oukrif, D.; Alison, M.R.; Wright, N.A. Bone marrow contribution to tumor-associated myofibroblasts and fibroblasts. Cancer Res. 2004, 64, 8492–8495.
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science (N. Y.) 1999, 284, 143–147.
- Glenn, J.D.; Whartenby, K.A. Mesenchymal stem cells: Emerging mechanisms of immunomodulation and therapy. World J. Stem Cells 2014, 6, 526–539.
- Majumdar, M.K.; Thiede, M.A.; Haynesworth, S.E.; Bruder, S.P.; Gerson, S.L. Human marrow-derived mesenchymal stem cells (MSCs) express hematopoietic cytokines and support long-term hematopoiesis when differentiated toward stromal and osteogenic lineages. J. Hematotherapy Stem Cell Res. 2000, 9, 841–848.
- Xie, H.; Sun, L.; Zhang, L.; Liu, T.; Chen, L.; Zhao, A.; Lei, Q.; Gao, F.; Zou, P.; Li, Q.; et al. Mesenchymal Stem Cell-Derived Microvesicles Support Ex Vivo Expansion of Cord Blood-Derived CD34(+) Cells. Stem Cells Int. 2016, 2016, 6493241.
- Mardpour, S.; Hamidieh, A.A.; Taleahmad, S.; Sharifzad, F.; Taghikhani, A.; Baharvand, H. Interaction between mesenchymal stromal cell-derived extracellular vesicles and immune cells by distinct protein content. J. Cell. Physiol. 2019, 234, 8249–8258.
- Toh, W.S.; Zhang, B.; Lai, R.C.; Lim, S.K. Immune regulatory targets of mesenchymal stromal cell exosomes/small extracellular vesicles in tissue regeneration. Cytotherapy 2018, 20, 1419–1426.
- Fafián-Labora, J.A.; Morente-López, M.; Arufe, M.C. Effect of aging on behaviour of mesenchymal stem cells. World J. Stem Cells 2019, 11, 337–346.
- Lei, Q.; Liu, T.; Gao, F.; Xie, H.; Sun, L.; Zhao, A.; Ren, W.; Guo, H.; Zhang, L.; Wang, H.; et al. Microvesicles as Potential Biomarkers for the Identification of Senescence in Human Mesenchymal Stem Cells. Theranostics 2017, 7, 2673–2689.
- Bron, D.; Ades, L.; Fulop, T.; Goede, V. Aging and malignant hemopathies. Haematologica 2015, 100, 571–574.
- Ergen, A.V.; Boles, N.C.; Goodell, M.A. Rantes/Ccl5 influences hematopoietic stem cell subtypes and causes myeloid skewing. Blood 2012, 119, 2500–2509.
- Berliner, N. Anemia in the elderly. Trans. Am. Clin. Climatol. Assoc. 2013, 124, 230–237.
- Mejia-Ramirez, E.; Florian, M.C. Understanding intrinsic hematopoietic stem cell aging. Haematologica 2020, 105, 22–37.
- Hayflick, L. THE LIMITED IN VITRO LIFETIME OF HUMAN DIPLOID CELL STRAINS. Exp. Cell Res. 1965, 37, 614–636.
- Choo, K.B.; Tai, L.; Hymavathee, K.S.; Wong, C.Y.; Nguyen, P.N.; Huang, C.J.; Cheong, S.K.; Kamarul, T. Oxidative stress-induced premature senescence in Wharton’s jelly-derived mesenchymal stem cells. Int. J. Med Sci. 2014, 11, 1201–1207.
- Baxter, M.A.; Wynn, R.F.; Jowitt, S.N.; Wraith, J.E.; Fairbairn, L.J.; Bellantuono, I. Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells (Dayt. Ohio) 2004, 22, 675–682.
- Cmielova, J.; Havelek, R.; Soukup, T.; Jiroutová, A.; Visek, B.; Suchánek, J.; Vavrova, J.; Mokry, J.; Muthna, D.; Bruckova, L.; et al. Gamma radiation induces senescence in human adult mesenchymal stem cells from bone marrow and periodontal ligaments. Int. J. Radiat. Biol. 2012, 88, 393–404.
- Idda, M.L.; McClusky, W.G.; Lodde, V.; Munk, R.; Abdelmohsen, K.; Rossi, M.; Gorospe, M. Survey of senescent cell markers with age in human tissues. Aging 2020, 12, 4052–4066.
- Zhou, S.; Greenberger, J.S.; Epperly, M.W.; Goff, J.P.; Adler, C.; Leboff, M.S.; Glowacki, J. Age-related intrinsic changes in human bone-marrow-derived mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell 2008, 7, 335–343.
- Gharibi, B.; Farzadi, S.; Ghuman, M.; Hughes, F.J. Inhibition of Akt/mTOR attenuates age-related changes in mesenchymal stem cells. Stem Cells (Dayt. Ohio) 2014, 32, 2256–2266.
- Ji, J.; Wu, Y.; Meng, Y.; Zhang, L.; Feng, G.; Xia, Y.; Xue, W.; Zhao, S.; Gu, Z.; Shao, X. JAK-STAT signaling mediates the senescence of bone marrow-mesenchymal stem cells from systemic lupus erythematosus patients. Acta Biochim. Biophys. Sin. 2017, 49, 208–215.
- Griukova, A.; Deryabin, P.; Sirotkina, M.; Shatrova, A.; Nikolsky, N.; Borodkina, A. P38 MAPK inhibition prevents polybrene-induced senescence of human mesenchymal stem cells during viral transduction. PLoS ONE 2018, 13, e0209606.
- Li, X.; Hong, Y.; He, H.; Jiang, G.; You, W.; Liang, X.; Fu, Q.; Han, S.; Lian, Q.; Zhang, Y. FGF21 Mediates Mesenchymal Stem Cell Senescence via Regulation of Mitochondrial Dynamics. Oxidative Med. Cell. Longev. 2019, 2019, 4915149.
- Neri, S.; Borzì, R.M. Molecular Mechanisms Contributing to Mesenchymal Stromal Cell Aging. Biomolecules 2020, 10, 340.
- Cakouros, D.; Gronthos, S. Epigenetic Regulation of Bone Marrow Stem Cell Aging: Revealing Epigenetic Signatures associated with Hematopoietic and Mesenchymal Stem Cell Aging. Aging Dis. 2019, 10, 174–189.
- Bork, S.; Pfister, S.; Witt, H.; Horn, P.; Korn, B.; Ho, A.D.; Wagner, W. DNA methylation pattern changes upon long-term culture and aging of human mesenchymal stromal cells. Aging Cell 2010, 9, 54–63.
- Fernández, A.F.; Bayón, G.F.; Urdinguio, R.G.; Toraño, E.G.; García, M.G.; Carella, A.; Petrus-Reurer, S.; Ferrero, C.; Martinez-Camblor, P.; Cubillo, I.; et al. H3K4me1 marks DNA regions hypomethylated during aging in human stem and differentiated cells. Genome Res. 2015, 25, 27–40.
- Yuan, H.F.; Zhai, C.; Yan, X.L.; Zhao, D.D.; Wang, J.X.; Zeng, Q.; Chen, L.; Nan, X.; He, L.J.; Li, S.T.; et al. SIRT1 is required for long-term growth of human mesenchymal stem cells. J. Mol. Med. (Berl. Ger.) 2012, 90, 389–400.
- Simic, P.; Zainabadi, K.; Bell, E.; Sykes, D.B.; Saez, B.; Lotinun, S.; Baron, R.; Scadden, D.; Schipani, E.; Guarente, L. SIRT1 regulates differentiation of mesenchymal stem cells by deacetylating β-catenin. EMBO Mol. Med. 2013, 5, 430–440.
- Peffers, M.J.; Collins, J.; Fang, Y.; Goljanek-Whysall, K.; Rushton, M.; Loughlin, J.; Proctor, C.; Clegg, P.D. Age-related changes in mesenchymal stem cells identified using a multi-omics approach. Eur. Cells Mater. 2016, 31, 136–159.
- Pan, H.; Guan, D.; Liu, X.; Li, J.; Wang, L.; Wu, J.; Zhou, J.; Zhang, W.; Ren, R.; Zhang, W.; et al. SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2. Cell Res. 2016, 26, 190–205.
- Kurosawa, S.; Iwama, A. Aging and leukemic evolution of hematopoietic stem cells under various stress conditions. Inflamm. Regen. 2020, 40, 29.
- So, A.Y.; Jung, J.W.; Lee, S.; Kim, H.S.; Kang, K.S. DNA methyltransferase controls stem cell aging by regulating BMI1 and EZH2 through microRNAs. PLoS ONE 2011, 6, e19503.
- Yang, R.; Yu, T.; Kou, X.; Gao, X.; Chen, C.; Liu, D.; Zhou, Y.; Shi, S. Tet1 and Tet2 maintain mesenchymal stem cell homeostasis via demethylation of the P2rX7 promoter. Nat. Commun. 2018, 9, 2143.
- Li, R.; Zhou, Y.; Cao, Z.; Liu, L.; Wang, J.; Chen, Z.; Xing, W.; Chen, S.; Bai, J.; Yuan, W.; et al. TET2 Loss Dysregulates the Behavior of Bone Marrow Mesenchymal Stromal Cells and Accelerates Tet2(−/−)-Driven Myeloid Malignancy Progression. Stem Cell Rep. 2018, 10, 166–179.
- Zhang, P.; Chen, Z.; Li, R.; Guo, Y.; Shi, H.; Bai, J.; Yang, H.; Sheng, M.; Li, Z.; Li, Z.; et al. Loss of ASXL1 in the bone marrow niche dysregulates hematopoietic stem and progenitor cell fates. Cell Discov. 2018, 4, 4.
- Malaquin, N.; Martinez, A.; Rodier, F. Keeping the senescence secretome under control: Molecular reins on the senescence-associated secretory phenotype. Exp. Gerontol. 2016, 82, 39–49.
- Mavrogonatou, E.; Pratsinis, H.; Papadopoulou, A.; Karamanos, N.K.; Kletsas, D. Extracellular matrix alterations in senescent cells and their significance in tissue homeostasis. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 75–76, 27–42.
- Lee, B.C.; Yu, K.R. Impact of mesenchymal stem cell senescence on inflammaging. BMB Rep. 2020, 53, 65–73.
- O’Hagan-Wong, K.; Nadeau, S.; Carrier-Leclerc, A.; Apablaza, F.; Hamdy, R.; Shum-Tim, D.; Rodier, F.; Colmegna, I. Increased IL-6 secretion by aged human mesenchymal stromal cells disrupts hematopoietic stem and progenitor cells’ homeostasis. Oncotarget 2016, 7, 13285–13296.
- Gnani, D.; Crippa, S.; Della Volpe, L.; Rossella, V.; Conti, A.; Lettera, E.; Rivis, S.; Ometti, M.; Fraschini, G.; Bernardo, M.E.; et al. An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program. Aging Cell 2019, 18, e12933.
- Urbanelli, L.; Buratta, S.; Sagini, K.; Tancini, B.; Emiliani, C. Extracellular Vesicles as New Players in Cellular Senescence. Int. J. Mol. Sci. 2016, 17, 1408.
- Kulkarni, R.; Bajaj, M.; Ghode, S.; Jalnapurkar, S.; Limaye, L.; Kale, V.P. Intercellular Transfer of Microvesicles from Young Mesenchymal Stromal Cells Rejuvenates Aged Murine Hematopoietic Stem Cells. Stem Cells (Dayt. Ohio) 2018, 36, 420–433.
- Terlecki-Zaniewicz, L.; Lämmermann, I.; Latreille, J.; Bobbili, M.R.; Pils, V.; Schosserer, M.; Weinmüllner, R.; Dellago, H.; Skalicky, S.; Pum, D.; et al. Small extracellular vesicles and their miRNA cargo are anti-apoptotic members of the senescence-associated secretory phenotype. Aging 2018, 10, 1103–1132.
- Robbins, P.D. Extracellular vesicles and aging. Stem Cell Investig. 2017, 4, 98.
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. Ser. ABiol. Sci. Med Sci. 2014, 69 (Suppl. 1), S4–S9.
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front. Immunol. 2018, 9, 586.
- Najar, M.; Ouhaddi, Y.; Bouhtit, F.; Melki, R.; Afif, H.; Boukhatem, N.; Merimi, M.; Fahmi, H. Empowering the immune fate of bone marrow mesenchymal stromal cells: Gene and protein changes. Inflamm. Res. 2019, 68, 167–176.
- Romieu-Mourez, R.; François, M.; Boivin, M.N.; Bouchentouf, M.; Spaner, D.E.; Galipeau, J. Cytokine modulation of TLR expression and activation in mesenchymal stromal cells leads to a proinflammatory phenotype. J. Immunol. 2009, 182, 7963–7973.
- Minciullo, P.L.; Catalano, A.; Mandraffino, G.; Casciaro, M.; Crucitti, A.; Maltese, G.; Morabito, N.; Lasco, A.; Gangemi, S.; Basile, G. Inflammaging and Anti-Inflammaging: The Role of Cytokines in Extreme Longevity. Arch. Immunol. Ther. Exp. 2016, 64, 111–126.
- Fulop, T.; Witkowski, J.M.; Olivieri, F.; Larbi, A. The integration of inflammaging in age-related diseases. Semin. Immunol. 2018, 40, 17–35.
- Zhao, J.L.; Ma, C.; O’Connell, R.M.; Mehta, A.; DiLoreto, R.; Heath, J.R.; Baltimore, D. Conversion of danger signals into cytokine signals by hematopoietic stem and progenitor cells for regulation of stress-induced hematopoiesis. Cell Stem Cell 2014, 14, 445–459.
- Ratushnyy, A.; Ezdakova, M.; Buravkova, L. Secretome of Senescent Adipose-Derived Mesenchymal Stem Cells Negatively Regulates Angiogenesis. Int. J. Mol. Sci. 2020, 21, 1802.
- Goedhart, M.; Cornelissen, A.S.; Kuijk, C.; Geerman, S.; Kleijer, M.; van Buul, J.D.; Huveneers, S.; Raaijmakers, M.; Young, H.A.; Wolkers, M.C.; et al. Interferon-Gamma Impairs Maintenance and Alters Hematopoietic Support of Bone Marrow Mesenchymal Stromal Cells. Stem Cells Dev. 2018, 27, 579–589.
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A new mesenchymal stem cell (MSC) paradigm: Polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE 2010, 5, e10088.
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008, 2, 141–150.
- Kim, J.; Hematti, P. Mesenchymal stem cell-educated macrophages: A novel type of alternatively activated macrophages. Exp. Hematol. 2009, 37, 1445–1453.
- Sepúlveda, J.C.; Tomé, M.; Fernández, M.E.; Delgado, M.; Campisi, J.; Bernad, A.; González, M.A. Cell senescence abrogates the therapeutic potential of human mesenchymal stem cells in the lethal endotoxemia model. Stem Cells (Dayt. Ohio) 2014, 32, 1865–1877.
- Yu, K.R.; Lee, J.Y.; Kim, H.S.; Hong, I.S.; Choi, S.W.; Seo, Y.; Kang, I.; Kim, J.J.; Lee, B.C.; Lee, S.; et al. A p38 MAPK-mediated alteration of COX-2/PGE2 regulates immunomodulatory properties in human mesenchymal stem cell aging. PLoS ONE 2014, 9, e102426.
- Yin, Y.; Wu, R.X.; He, X.T.; Xu, X.Y.; Wang, J.; Chen, F.M. Influences of age-related changes in mesenchymal stem cells on macrophages during in-vitro culture. Stem Cell Res. Ther. 2017, 8, 153.
- Huang, R.; Qin, C.; Wang, J.; Hu, Y.; Zheng, G.; Qiu, G.; Ge, M.; Tao, H.; Shu, Q.; Xu, J. Differential effects of extracellular vesicles from aging and young mesenchymal stem cells in acute lung injury. Aging 2019, 11, 7996–8014.
- Ziegler, P.; Boettcher, S.; Takizawa, H.; Manz, M.G.; Brümmendorf, T.H. LPS-stimulated human bone marrow stroma cells support myeloid cell development and progenitor cell maintenance. Ann. Hematol. 2016, 95, 173–178.
- Shi, C.; Jia, T.; Mendez-Ferrer, S.; Hohl, T.M.; Serbina, N.V.; Lipuma, L.; Leiner, I.; Li, M.O.; Frenette, P.S.; Pamer, E.G. Bone marrow mesenchymal stem and progenitor cells induce monocyte emigration in response to circulating toll-like receptor ligands. Immunity 2011, 34, 590–601.
- Granero-Moltó, F.; Weis, J.A.; Miga, M.I.; Landis, B.; Myers, T.J.; O’Rear, L.; Longobardi, L.; Jansen, E.D.; Mortlock, D.P.; Spagnoli, A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells (Dayt. Ohio) 2009, 27, 1887–1898.
- Coipeau, P.; Rosset, P.; Langonne, A.; Gaillard, J.; Delorme, B.; Rico, A.; Domenech, J.; Charbord, P.; Sensebe, L. Impaired differentiation potential of human trabecular bone mesenchymal stromal cells from elderly patients. Cytotherapy 2009, 11, 584–594.
- Wagner, W.; Horn, P.; Castoldi, M.; Diehlmann, A.; Bork, S.; Saffrich, R.; Benes, V.; Blake, J.; Pfister, S.; Eckstein, V.; et al. Replicative senescence of mesenchymal stem cells: A continuous and organized process. PLoS ONE 2008, 3, e2213.
- Justesen, J.; Stenderup, K.; Eriksen, E.F.; Kassem, M. Maintenance of osteoblastic and adipocytic differentiation potential with age and osteoporosis in human marrow stromal cell cultures. Calcif. Tissue Int. 2002, 71, 36–44.
- Fickert, S.; Schröter-Bobsin, U.; Gross, A.F.; Hempel, U.; Wojciechowski, C.; Rentsch, C.; Corbeil, D.; Günther, K.P. Human mesenchymal stem cell proliferation and osteogenic differentiation during long-term ex vivo cultivation is not age dependent. J. Bone Miner. Metab. 2011, 29, 224–235.
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173.
- Zaim, M.; Karaman, S.; Cetin, G.; Isik, S. Donor age and long-term culture affect differentiation and proliferation of human bone marrow mesenchymal stem cells. Ann. Hematol. 2012, 91, 1175–1186.
- Zhang, W.; Ou, G.; Hamrick, M.; Hill, W.; Borke, J.; Wenger, K.; Chutkan, N.; Yu, J.; Mi, Q.S.; Isales, C.M.; et al. Age-related changes in the osteogenic differentiation potential of mouse bone marrow stromal cells. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2008, 23, 1118–1128.
- Alameda, D.; Saez, B.; Lara-Astiaso, D.; Sarvide, S.; Lasa, M.; Alignani, D.; Rodriguez, I.; Garate, S.; Vilas, A.; Paiva, B.; et al. Characterization of freshly isolated mesenchymal stromal cells from healthy and multiple myeloma bone marrow: Transcriptional modulation of the microenvironment. Haematologica 2020, 105.
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell. Biochem. 2006, 99, 1233–1239.
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29.
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952.
- Li, H.; Liu, P.; Xu, S.; Li, Y.; Dekker, J.D.; Li, B.; Fan, Y.; Zhang, Z.; Hong, Y.; Yang, G.; et al. FOXP1 controls mesenchymal stem cell commitment and senescence during skeletal aging. J. Clin. Investig. 2017, 127, 1241–1253.
- Wu, M.; Wang, Y.; Shao, J.Z.; Wang, J.; Chen, W.; Li, Y.P. Cbfβ governs osteoblast-adipocyte lineage commitment through enhancing β-catenin signaling and suppressing adipogenesis gene expression. Proc. Natl. Acad. Sci. USA 2017, 114, 10119–10124.
- Nishikawa, K.; Nakashima, T.; Takeda, S.; Isogai, M.; Hamada, M.; Kimura, A.; Kodama, T.; Yamaguchi, A.; Owen, M.J.; Takahashi, S.; et al. Maf promotes osteoblast differentiation in mice by mediating the age-related switch in mesenchymal cell differentiation. J. Clin. Investig. 2010, 120, 3455–3465.
- Moerman, E.J.; Teng, K.; Lipschitz, D.A.; Lecka-Czernik, B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: The role of PPAR-gamma2 transcription factor and TGF-beta/BMP signaling pathways. Aging Cell 2004, 3, 379–389.
- Davis, C.; Dukes, A.; Drewry, M.; Helwa, I.; Johnson, M.H.; Isales, C.M.; Hill, W.D.; Liu, Y.; Shi, X.; Fulzele, S.; et al. MicroRNA-183-5p Increases with Age in Bone-Derived Extracellular Vesicles, Suppresses Bone Marrow Stromal (Stem) Cell Proliferation, and Induces Stem Cell Senescence. Tissue Eng. Part A 2017, 23, 1231–1240.
- Xu, R.; Shen, X.; Si, Y.; Fu, Y.; Zhu, W.; Xiao, T.; Fu, Z.; Zhang, P.; Cheng, J.; Jiang, H. MicroRNA-31a-5p from aging BMSCs links bone formation and resorption in the aged bone marrow microenvironment. Aging Cell 2018, 17, e12794.
- Guidi, N.; Sacma, M.; Ständker, L.; Soller, K.; Marka, G.; Eiwen, K.; Weiss, J.M.; Kirchhoff, F.; Weil, T.; Cancelas, J.A.; et al. Osteopontin attenuates aging-associated phenotypes of hematopoietic stem cells. EMBO J. 2017, 36, 840–853.
- Stier, S.; Ko, Y.; Forkert, R.; Lutz, C.; Neuhaus, T.; Grünewald, E.; Cheng, T.; Dombkowski, D.; Calvi, L.M.; Rittling, S.R.; et al. Osteopontin is a hematopoietic stem cell niche component that negatively regulates stem cell pool size. J. Exp. Med. 2005, 201, 1781–1791.
- Kennedy, D.E.; Knight, K.L. Inhibition of B Lymphopoiesis by Adipocytes and IL-1-Producing Myeloid-Derived Suppressor Cells. J. Immunol. 2015, 195, 2666–2674.
- Aguilar-Navarro, A.G.; Meza-León, B.; Gratzinger, D.; Juárez-Aguilar, F.G.; Chang, Q.; Ornatsky, O.; Tsui, H.; Esquivel-Gómez, R.; Hernández-Ramírez, A.; Xie, S.Z.; et al. Human Aging Alters the Spatial Organization between CD34+ Hematopoietic Cells and Adipocytes in Bone Marrow. Stem Cell Rep. 2020, 15, 317–325.
- Ambrosi, T.H.; Scialdone, A.; Graja, A.; Gohlke, S.; Jank, A.M.; Bocian, C.; Woelk, L.; Fan, H.; Logan, D.W.; Schürmann, A.; et al. Adipocyte Accumulation in the Bone Marrow during Obesity and Aging Impairs Stem Cell-Based Hematopoietic and Bone Regeneration. Cell Stem Cell 2017, 20, 771–784.e776.
- Mau, T.; Yung, R. Adipose tissue inflammation in aging. Exp. Gerontol. 2018, 105, 27–31.
- Tormin, A.; Brune, J.C.; Olsson, E.; Valcich, J.; Neuman, U.; Olofsson, T.; Jacobsen, S.E.; Scheding, S. Characterization of bone marrow-derived mesenchymal stromal cells (MSC) based on gene expression profiling of functionally defined MSC subsets. Cytotherapy 2009, 11, 114–128.
- Kuçi, S.; Kuçi, Z.; Kreyenberg, H.; Deak, E.; Pütsch, K.; Huenecke, S.; Amara, C.; Koller, S.; Rettinger, E.; Grez, M.; et al. CD271 antigen defines a subset of multipotent stromal cells with immunosuppressive and lymphohematopoietic engraftment-promoting properties. Haematologica 2010, 95, 651–659.
- Flores-Figueroa, E.; Varma, S.; Montgomery, K.; Greenberg, P.L.; Gratzinger, D. Distinctive contact between CD34+ hematopoietic progenitors and CXCL12+ CD271+ mesenchymal stromal cells in benign and myelodysplastic bone marrow. Lab. Investig. 2012, 92, 1330–1341.
- Li, H.; Ghazanfari, R.; Zacharaki, D.; Ditzel, N.; Isern, J.; Ekblom, M.; Méndez-Ferrer, S.; Kassem, M.; Scheding, S. Low/negative expression of PDGFR-α identifies the candidate primary mesenchymal stromal cells in adult human bone marrow. Stem Cell Rep. 2014, 3, 965–974.
- Tormin, A.; Li, O.; Brune, J.C.; Walsh, S.; Schütz, B.; Ehinger, M.; Ditzel, N.; Kassem, M.; Scheding, S. CD146 expression on primary nonhematopoietic bone marrow stem cells is correlated with in situ localization. Blood 2011, 117, 5067–5077.
- Maijenburg, M.W.; Kleijer, M.; Vermeul, K.; Mul, E.P.; van Alphen, F.P.; van der Schoot, C.E.; Voermans, C. The composition of the mesenchymal stromal cell compartment in human bone marrow changes during development and aging. Haematologica 2012, 97, 179–183.
- Duscher, D.; Rennert, R.C.; Januszyk, M.; Anghel, E.; Maan, Z.N.; Whittam, A.J.; Perez, M.G.; Kosaraju, R.; Hu, M.S.; Walmsley, G.G.; et al. Aging disrupts cell subpopulation dynamics and diminishes the function of mesenchymal stem cells. Sci. Rep. 2014, 4, 7144.
- Khong, S.M.L.; Lee, M.; Kosaric, N.; Khong, D.M.; Dong, Y.; Hopfner, U.; Aitzetmüller, M.M.; Duscher, D.; Schäfer, R.; Gurtner, G.C. Single-Cell Transcriptomics of Human Mesenchymal Stem Cells Reveal Age-Related Cellular Subpopulation Depletion and Impaired Regenerative Function. Stem Cells (Dayt. Ohio) 2019, 37, 240–246.
- Isern, J.; García-García, A.; Martín, A.M.; Arranz, L.; Martín-Pérez, D.; Torroja, C.; Sánchez-Cabo, F.; Méndez-Ferrer, S. The neural crest is a source of mesenchymal stem cells with specialized hematopoietic stem cell niche function. eLife 2014, 3, e03696.
- Wang, X.; Zou, X.; Zhao, J.; Wu, X.; Lingling, E.; Feng, L.; Wang, D.; Zhang, G.; Xing, H.; Liu, H. Site-Specific Characteristics of Bone Marrow Mesenchymal Stromal Cells Modify the Effect of Aging on the Skeleton. Rejuvenation Res. 2016, 19, 351–361.


