 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Marc Edwards | + 2310 word(s) | 2310 | 2020-11-26 07:06:03 | | | |
| 2 | Vicky Zhou | -51 word(s) | 2259 | 2020-12-22 06:47:11 | | |
Video Upload Options
The growth of Legionella pneumophila and other opportunistic pathogens (OPs) in drinking water premise plumbing poses an increasing public health concern. Premise plumbing is constructed of a variety of materials, creating complex environments that vary chemically, microbiologically, spatially, and temporally in a manner likely to influence survival and growth of OPs. Plastic pipes can leach organic carbon, but demonstrate a lower disinfectant demand and fewer water chemistry interactions. Iron pipes may provide OPs with nutrients directly or indirectly, exhibiting a high disinfectant demand and potential to form scales with high surface areas suitable for biofilm colonization. While copper pipes are known for their antimicrobial properties, evidence of their efficacy for OP control is inconsistent. Under some as yet not fully understood circumstances, copper’s interactions with premise plumbing water chemistry and resident microbes can encourage growth of OPs. Plumbing design, configuration, and operation can be manipulated to control such interactions and health outcomes. Influences of pipe materials on OP physiology should also be considered, including the possibility of influencing virulence and antibiotic resistance. In conclusion, all known pipe materials have a potential to either stimulate or inhibit OP growth, depending on the circumstances.
1. Introduction
Legionnaires’ Disease is the “leading cause of reportable waterborne illness” in the United States [1][2], with 52,000–70,000 cases per year [1][3][4], 8000–18,000 hospitalizations [5], an overall mortality rate of 15% [4], and high healthcare and legal costs [2][6][7][8]. Bacteria belonging to the genus Legionella are the causative agent of Legionnaires’ disease and Pontiac Fever, which infect the human respiratory system via inhalation or aspiration. Legionella is classified as “opportunistic” because it preferentially infects those with underlying illnesses or weakened immune systems [4][8][9]. To date more than 60 Legionella species have been identified [10], with Legionella pneumophila being the species most commonly attributed to human disease [11]. Legionella can be found even in “the most aggressively treated drinking water” [12]. Studies have confirmed that potable water is a key source of infection [1][4][13][14][15][16][17] for both hospital- and community-acquired cases [18][19][20]. Other opportunistic pathogens (OPs) such as nontuberculous mycobacteria (NTM), Pseudomonas aeruginosa, and Acanthamoebae, can similarly be transmitted via tap water and tend to infect individuals belonging to certain risk groups [8].
To infect humans, Legionella and other OPs must be present in tap water at the point of use. While Legionella can occasionally survive drinking water treatment and be transported through the main water distribution system, the primary environment for Legionella proliferation to numbers needed to infect humans generally occurs in building or “premise” plumbing [21][22]. Premise plumbing includes the service pipe that connects buildings to the water main, in addition to the full array of components comprising cold and hot portions of a building’s potable water system [8]. Premise plumbing is characterized by high surface area to volume ratios, longer stagnation times, low disinfectant residual, areas with excess sediment and scale, chemically and biologically reactive plumbing materials, and water with relatively warm temperatures. Such conditions can create ideal micro- and macro-environmental niches for growth of various OPs [1][8][23].
Premise plumbing is a key conduit for human exposure via showering, handwashing, and other applications that create airborne aerosols [24]. Legionella has been detected in faucets, showerheads, decorative fountains, grocery store mist systems, ice machines, and cooling towers [13][14][16][25]. Larger buildings with more complex plumbing systems are more likely to create physicochemical conditions suited for Legionella proliferation, but it is also often detectable in water mains and residences with simple conventional hot and cold water plumbing systems [17][26][27]. A Centre for Disease Control (CDC) summary of Legionnaires’ Disease potable water outbreak investigations from 2000–2014, concluded that 85% of the cases had “deficiencies” in water system maintenance within buildings as a contributing factor [28] and that water chemistry flowing into buildings is one, but not the only, predictor of Legionella incidence [29][30].
The mechanisms by which premise plumbing influences L. pneumophila and other OPs, as well as the broader premise plumbing microbiome, are varied and complex (Figure 1). The influent water chemistry has been found to influence Legionella, and also strongly shape the plumbing microbiome, especially through the delivery of growth-promoting nutrients, growth-inhibiting disinfectants, and influent microorganisms [31][32][33][34]. The ecological interactions among microorganisms in biofilms of building plumbing systems can also help overcome barriers to growth from low nutrient levels and disinfectants [24][35][36]. Conversely, other interactions, such as competition, exclusion, predation, or inactivation of symbiotic organisms, may inhibit the growth of OPs [37]. The selective pressures in premise plumbing might also alter the physiologies of resident microbes in a manner that influences infectivity [38]. All these phenomena are further complicated by the fact that premise plumbing configurations, hydraulics, temperature, and water use patterns including velocity, flow or stagnation events, all differ significantly from building to building. In particular, there is strong variability due to occupancy, building size, water heater design, water saving devices, storage and other factors [39][40]. Thus, while there are many overarching similarities, every premise plumbing system is at least as variable as the occupants’ unique water use patterns and habits.
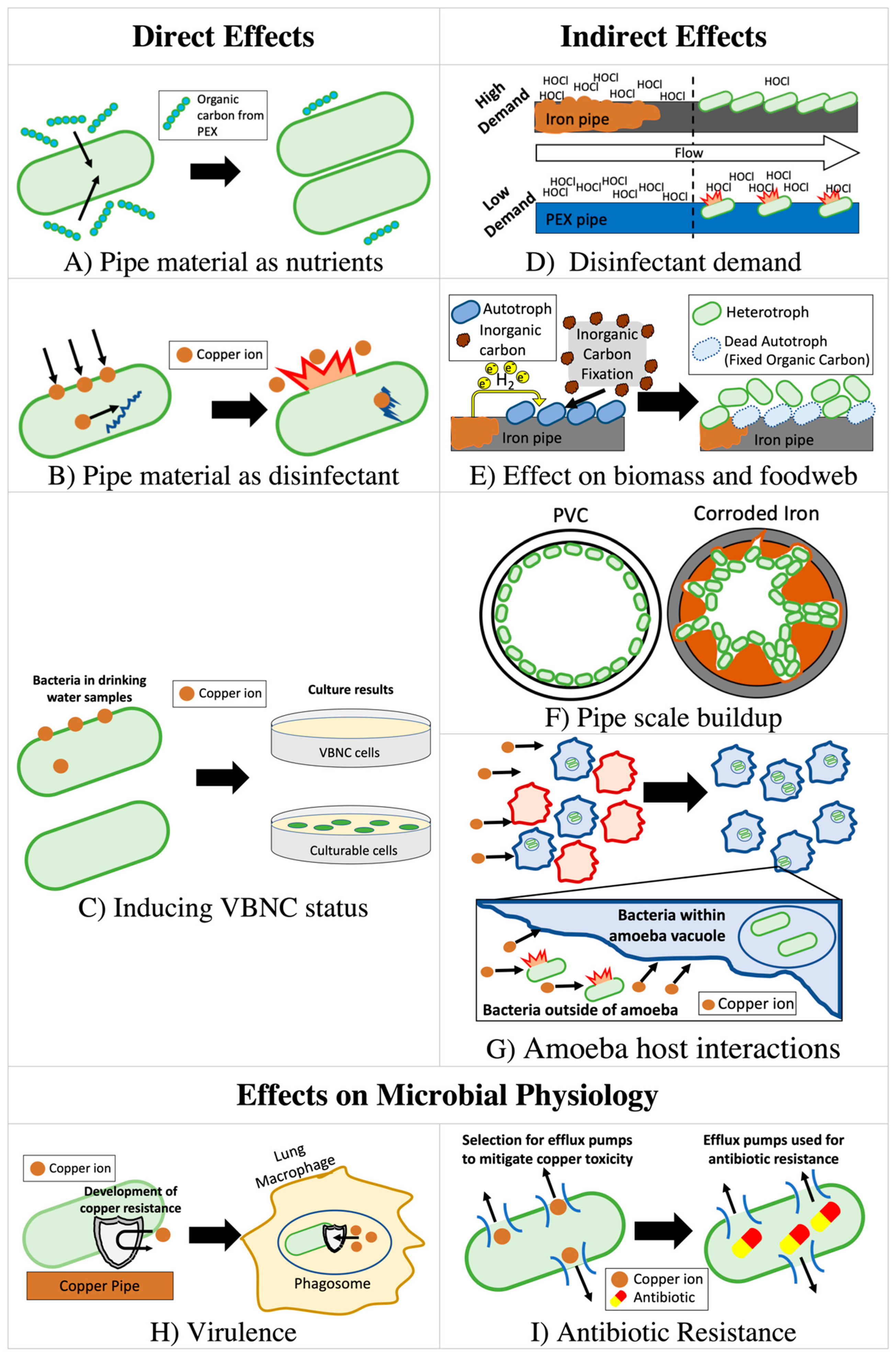
The type of pipe material can also strongly influence the relationship between premise plumbing materials and OPs through both direct effects (interaction with chemical species released from pipe) and indirect effects (secondary consequences of released material from pipes) by altering the level of nutrients, disinfectants, and microbial biomass (Figure 1). Selection of pipe material can therefore strongly affect chemistry, biological stability [41], and microbiome composition [42] of the drinking water.
2. Mediating Role of Microbiome and other Microbiological Considerations
2.1. The Role of Pipe Material in Shaping the Premise Plumbing Microbiome and Resident Amoeba Host Organisms
Interactions between OPs and the microbial communities surrounding them are key to OP proliferation and are likely influenced by pipe materials. OPs can be parasitic to free-living amoebae that first prey upon them in drinking water biofilms, before they reproduce inside and eventually kill the host organism [24]. In fact, there is some doubt that Legionella actually reproduces significantly in drinking water outside of an amoeba host [43]. Amoebae can also protect OPs from disinfectants and provide access to nutrients. For example, Legionella exclusively use amino acids, which are abundant in amoeba vacuoles, as a carbon source [44][45][46][47][48][49][50]. Thus, although poorly studied, any factor altering growth of key host amoebae (including Acanthamoeba, Vermamoeba, and Naegleria) is expected to indirectly affect growth of OPs, including L. pneumophila, P. aeruginosa, and NTM [51][44][45][46][47][48][52][53][54][55]. In one experiment, copper coupons were found to host more Acanthamoeba polyphaga than PVC coupons [56], possibly because copper hosts less diverse eukaryotic communities [57][58] and limits competition for A. polyphaga. As a result, L. pneumophila grew and shed to the bulk waters in higher numbers on these copper coupons than on PVC coupons if co-inoculated with A. polyphaga 116[59].
Interbacterial interactions may also influence the growth of OPs. Broadly speaking, OPs benefit from the biofilm community through access to nutrients and protection from disinfectants [24][35][36][60]. Some studies have identified correlations between specific taxa and OPs in premise plumbing [61], cooling towers [62] and drinking water distribution systems [63]. However, the significance of these correlations to premise plumbing material selection is not well understood, as most studies examining differences in bacterial communities focus on very broad measures of community structure [64][65][57][66][58][67][68][69]. Certain waterborne bacteria are known to produce toxins that inhibit L. pneumophila growth [66][70] or exude other compounds that have secondary bacteriostatic effects on Legionella [71]. Intra-bacterial inhibition also may be mediated through amoebae by reducing host uptake [72][73] or killing the host population [74][75][76]. More research is needed to elucidate how the broad ecological differences resulting from pipe material influence these interactions. Integration of metagenomic or meta-transcriptomic analyses targeting the production of bacteriocins or other toxins with known effects on OPs could elucidate the ecological effects of taxonomic shifts resulting from pipe material. Interrupting OP-amoeba endosymbiosis through the enrichment of preferential non-OP amoeba prey [72][73] has been suggested as a probiotic means of controlling OPs [77], and pipe material could be explored as a means of enrichment of these taxa.
2.2. Variation in Copper Tolerance Among Species and Strains
Strain-to-strain differences in intrinsic tolerance of copper, acclimation to copper concentrations with time through induction of the appropriate genes, or acquisition of copper resistance via mutation or horizontal gene transfer in premise plumbing might explain some of the discrepancies in variable outcomes of copper on OPs (Table 2). Legionella [78] and other OPs [79] may acclimate to high copper levels through the expression of copper detoxification or efflux systems. Bedard et al. [78] reported four-fold differences in the copper tolerance of environmentally-isolated L. pneumophila strains, noting that more resistant strains showed increased copper ATPase copA expression, speculating that their increased tolerance may also be a result of higher biofilm production. Strikingly, Williams et al. [79] showed that, during exposure to 95 mg/L of copper over 6 h in liquid culture, culturable A. baumannii levels (CFU/mL) could increase by 2-logs or decrease by 2-logs, depending on the strain. The authors identified putative copper detoxification and efflux systems within the genome of the most resistant isolate and identified specific genes that were upregulated in response to copper exposure.
2.3. Confounding Effects of VBNC Bacteria
The discovery of VBNC bacteria has complicated prior understanding for all OP control strategies, including copper. Virtually all prior work relied on culture methods to determine copper’s efficacy for killing OPs [80][81][82][83][84][85][86][87][88][89][90], but some microbes rendered not culturable might remain viable and still infect host amoebae or humans [91][92][93][94][95]. The existence of VBNC pathogens in premise plumbing has been demonstrated by comparing culture-based numbers with those enumerated via fluorescence (e.g., live/dead) and molecular-based (e.g., quantitative polymerase chain reaction) monitoring methods [96].
Bench-scale studies examining copper’s antimicrobial efficacy have found discrepancies between culture-based and molecular-based numbers of L. pneumophila [97][59] that are also suggestive of a copper-induced VBNC state. Similar discrepancies have been noted for P. aeruginosa, Stenotrophomonas maltophilia, and M. avium [98][87][99][100][101]. Evidence of copper-induced VBNC activity is particularly strong in the case of P. aeruginosa, where one study applied multiple non-culture-based measures of viability [99][100]. Furthermore, VBNC P. aeruginosa have been shown to partially recover infectivity after removal of copper from solution [100][101]. To understand how VBNC bacteria contribute to OP infections, additional studies are needed to delineate the premise plumbing conditions more precisely that induce VBNC status and to confirm the range of functionality maintained in this state. A primary challenge in achieving this is that there are currently no reliable methods for confidently enumerating VBNC bacteria.
2.4. Virulence
The premise plumbing environment exhibits several features that could possibly contribute to the virulence of resident OPs. Wargo [38] describes features of drinking water plumbing that could prime OPs to infect cystic fibrosis patients, although the interactions described in this review could also pose risk to otherwise immunocompromised individuals. Such features that are relevant to pipe material include [38]:
- Elevated copper levels, selecting for resistance to copper overload within macrophage phagosomes, a component of the innate immune response [102].
- Elevated iron levels, influencing interactions between iron homeostasis and virulence.
- Exposure to lipids, which are generally not well removed by drinking water treatment, priming OPs for lipid-rich environments within hosts. Accumulation of phospholipid fatty acids has been shown to be greater in the biofilms of polyethylene pipes than copper pipes, though these lipids were putatively associated with bacteria [103].
- Low DO levels, selecting for OPs capable of survival in low DO regions of the biofilm in infected host tissue.
- Exposure to eukaryotic predation, selecting for resistance to the host’s immune response (e.g., lung macrophages) or enhanced virulence.
2.5. Antibiotic Resistance and Tolerance
Copper, among other heavy metals has been shown to exert selection pressure, leading to enhanced survival of antibiotic resistant bacteria. In fact, heavy-metal-associated co-selection and cross-selection has been proposed to be as much of a concern for environmental propagation of antibiotic resistance as antibiotics themselves [104]. Increases in antibiotic resistance genes at the community scale have been identified after long-term copper exposure in soil [105][106][107][108], sediment [109], and drinking water [109]. Bench-scale tests using bacterial isolates from biofilters [110] and wastewater [111] inoculated into growth media have shown that a selective or inductive effect of copper can take places within hours. However, these studies were performed with copper concentrations 5–77 times greater than the 1.3 mg/L US EPA copper action level and similarly in exceedance of the Chinese Standard for Drinking Water Quality of 1 mg/L [112] and WHO Guideline for Drinking-Water Quality of 2 mg/L [113]. Thus, these concentrations may not be representative of potable water systems. One study examining antibiotic resistant and sensitive strains of Staphylococcus aureus showed that the more antibiotic resistant strain survived longer in a copper container [114]. As discussed above, copper may also better support Acanthamoeba than other materials, while in one study L. pneumophila grown within A. polyphaga demonstrated increased tolerance to all antibiotics tested (rifampin, ciprofloxacin, and erythromycin) compared to those grown in culture media [115]. The role of copper plumbing and other pipe materials in these emerging areas of research is worthy of further investigation.
References
- National Academies of Sciences, Engineering, and Medicine. Management of Legionella in Water Systems; The National Academies Press: Washington, DC, USA, 2019; ISBN 978-0-309-49947-7.
- Ashbolt, N.J. Microbial Contamination of Drinking Water and Human Health from Community Water Systems. Curr. Environ. Health Rep. 2015, 2, 95.
- United States Occupational Safety & Health Administration Legionnaires’ Disease: Facts and Frequently Asked Questions. Available online: https://www.osha.gov/dts/osta/otm/legionnaires/faq.html (accessed on 1 July 2020).
- United States Occupational Safety & Health Administration Legionnaires’ Disease. Available online: https://www.osha.gov/dts/osta/otm/otm_iii/otm_iii_7.html (accessed on 1 July 2020).
- CDC Legionella (Legionnaires’ Disease and Pontiac Fever) Surveillance and Reporting. Available online: http://www.cdc.gov/legionella/surv-reporting.html (accessed on 1 July 2020).
- Singh, R.; Hamilton, K.A.; Rasheduzzaman, M.; Yang, Z.; Kar, S.; Fasnacht, A.; Masters, S.V.; Gurian, P.L. Managing Water Quality in Premise Plumbing: Subject Matter Experts’ Perspectives and a Systematic Review of Guidance Documents. Water 2020, 12, 347.
- McCoy, W.F. Preventing Legionellosis; IWA Publishing: London, UK, 2005; ISBN 1843390949.
- Falkinham, J.; Pruden, A.; Edwards, M. Opportunistic Premise Plumbing Pathogens: Increasingly Important Pathogens in Drinking Water. Pathogens 2015, 4, 373.
- Centers for Disease Control Legionella (Legionnaires’ Disease and Pontiac Fever). Available online: https://www.cdc.gov/legionella/index.html (accessed on 1 July 2020).
- Gomez-Valero, L.; Rusniok, C.; Rolando, M.; Neou, M.; Dervins-Ravault, D.; Demirtas, J.; Rouy, Z.; Moore, R.J.; Chen, H.; Petty, N.K.; et al. Comparative analyses of Legionella species identifies genetic features of strains causing Legionnaires’ disease. Genome Biol. 2014, 15, 505.
- Association of Water Technologies Legionella 2019: A Position Statement and Guidance Document. Available online: https://www.awt.org/pub/035C2942-03BE-3BFF-08C3-4C686FB7395C (accessed on 15 November 2020).
- Allen, M.J.; Edberg, S.C.; Clancy, J.L.; Hrudey, S.E. Drinking water microbial myths. Crit. Rev. Microbiol. 2015, 41, 366–373.
- Barrabeig, I.; Rovira, A.; Garcia, M.; Oliva, J.M.; Vilamala, A.; Ferrer, M.D.; SabriÀ, M.; DomÍNguez, A. Outbreak of Legionnaires’ disease associated with a supermarket mist machine. Epidemiol. Infect. 2010, 138, 1823–1828.
- Mahoney, F.J.; Hoge, C.W.; Farley, T.A.; Barbaree, J.M.; Breiman, R.F.; Benson, R.F.; McFarland, L.M. Communitywide Outbreak of Legionnaires’ Disease Associated with a Grocery Store Mist Machine. J. Infect. Dis. 1992, 165, 736–739.
- Stout, J.E.; Yu, V.L.; Yee, Y.C.; Vaccarello, S.; Diven, W.; Lee, T.C. Legionella pneumophila in residential water supplies: Environmental surveillance with clinical assessment for Legionnaires’ disease. Epidemiol. Infect. 1992, 109, 49–57.
- Stout, J.E.; Victor, L.Y.; Muraca, P. Isolation of Legionella pneumophila from the cold water of hospital ice machines: Implications for origin and transmission of the organism. Infect. Control Hosp. Epidemiol. 1985, 6, 141–146.
- Rhoads, W.J.; Bradley, T.N.; Mantha, A.; Buttling, L.; Keane, T.; Pruden, A.; Edwards, M.A. Residential water heater cleaning and occurrence of Legionella in Flint, MI. Water Res. 2020, 115439.
- Gobin, I.; Newton, P.R.; Hartland, E.L.; Newton, H.J. Infections caused by nonpneumophila species of Legionella. Rev. Med. Microbiol. 2009, 20, 1–11.
- Joseph, C.A. Legionnaires’ disease in Europe 2000–2002. Epidemiol. Infect. 2004, 132, 417–424.
- Stout, J.E.; Yu, V.L.; Muraca, P.; Joly, J.; Troup, N.; Tompkins, L.S. Potable water as a cause of sporadic cases of community-acquired legionnaires’ disease. N. Engl. J. Med. 1992, 326, 151–155.
- Fliermans, C.B.; Cherry, W.B.; Orrison, L.H.; Smith, S.J.; Tison, D.L.; Pope, D.H. Ecological distribution of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 41, 9–16.
- Lee, G.F.; Jones-Lee, A. Public Health Significance of Waterborne Pathogens in Domestic Water Supplies and Reclaimed Water. In Report to State of California Environmental Protection Agency Comparative Risk Project; California Environmental Protection Agency: Berkeley, CA, USA, 1993.
- Borella, P.; Montagna, M.T.; Romano-Spica, V.; Stampi, S.; Stancanelli, G.; Triassi, M.; Neglia, R.; Marchesi, I.; Fantuzzi, G.; Tatò, D.; et al. Legionella Infection Risk from Domestic Hot Water. Emerg. Infect. Dis. 2004, 10, 457–464.
- Falkinham III, J.O. Common Features of Opportunistic Premise Plumbing Pathogens. Int. J. Environ. Res. Public Health 2015, 12, 4533–4545.
- Parr, A.; Whitney, E.A.; Berkelman, R.L. Legionellosis on the Rise: A Review of Guidelines for Prevention in the United States. J. public Health Manag. Pract. 2015, 21, E17–E26.
- Rhoads, W.J.; Garner, E.; Ji, P.; Zhu, N.; Parks, J.; Schwake, D.O.; Pruden, A.; Edwards, M.A. Distribution System Operational Deficiencies Coincide with Reported Legionnaires’ Disease Clusters in Flint, Michigan. Environ. Sci. Technol. 2017, 51, 11986–11995.
- Mercante, J.W.; Winchell, J.M. Current and Emerging Legionella Diagnostics for Laboratory and Outbreak Investigations. Clin. Microbiol. Rev. 2015, 28, 95–133.
- Garrison, L.E. Vital Signs: Deficiencies in Environmental Control Identified in Outbreaks of Legionnaires’ Disease—North America, 2000–2014. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 576–584.
- Pierre, D.; Baron, J.L.; Ma, X.; Sidari, F.P.; Wagener, M.M.; Stout, J.E. Water Quality as a Predictor of Legionella Positivity of Building Water Systems. Pathogens 2019, 8, 295.
- Perrin, Y.; Bouchon, D.; Delafont, V.; Moulin, L.; Héchard, Y. Microbiome of drinking water: A full-scale spatio-temporal study to monitor water quality in the Paris distribution system. Water Res. 2019, 149, 375–385.
- Liu, G.; Zhang, Y.; Knibbe, W.-J.; Feng, C.; Liu, W.; Medema, G.; van der Meer, W. Potential impacts of changing supply-water quality on drinking water distribution: A review. Water Res. 2017, 116, 135–148.
- Pinto, A.J.; Xi, C.; Raskin, L. Bacterial community structure in the drinking water microbiome is governed by filtration processes. Environ. Sci. Technol. 2012, 46, 8851–8859.
- Wang, H.; Masters, S.; Falkinham, J.O.; Edwards, M.A.; Pruden, A. Distribution System Water Quality Affects Responses of Opportunistic Pathogen Gene Markers in Household Water Heaters. Environ. Sci. Technol. 2015, 49, 8416–8424.
- Baron, J.L.; Vikram, A.; Duda, S.; Stout, J.E.; Bibby, K. Shift in the Microbial Ecology of a Hospital Hot Water System following the Introduction of an On-Site Monochloramine Disinfection System. PLoS ONE 2014, 9.
- Schwering, M.; Song, J.; Louie, M.; Turner, R.J.; Ceri, H. Multi-species biofilms defined from drinking water microorganisms provide increased protection against chlorine disinfection. Biofouling 2013, 29, 917–928.
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413.
- Wang, H.; Edwards, M.A.; Falkinham, J.O.; Pruden, A. Probiotic Approach to Pathogen Control in Premise Plumbing Systems? A Review. Environ. Sci. Technol. 2013, 47, 10117–10128.
- Wargo, M.J. Is the Potable Water System an Advantageous Preinfection Niche for Bacteria Colonizing the Cystic Fibrosis Lung? MBio 2019, 10, e00883-19.
- Rhoads, J.W.; Pearce, A.; Pruden, A.; Edwards, M.A. Anticipating the Effects of Green Buildings on Water Quality and Infrastructure. J. Am. Water Works Assoc. 2015, 107, 50.
- Liu, G.; Tao, Y.; Zhang, Y.; Lut, M.; Knibbe, W.-J.; van der Wielen, P.; Liu, W.; Medema, G.; van der Meer, W. Hotspots for selected metal elements and microbes accumulation and the corresponding water quality deterioration potential in an unchlorinated drinking water distribution system. Water Res. 2017, 124, 435–445.
- Proctor, C.R.; Hammes, F. Drinking water microbiology - from measurement to management. Curr. Opin. Biotechnol. 2015, 33, 87–94.
- Neu, L.; Hammes, F. Feeding the Building Plumbing Microbiome: The Importance of Synthetic Polymeric Materials for Biofilm Formation and Management. Water 2020, 12, 1774.
- Ewann, F.; Hoffman, P.S. Cysteine Metabolism in Legionella pneumophila: Characterization of an L-Cystine-Utilizing Mutant. Appl. Environ. Microbiol. 2006, 72, 3993–4000.
- Barker, J.; Brown, M.R.; Collier, P.J.; Farrell, I.; Gilbert, P. Relationship between Legionella pneumophila and Acanthamoeba polyphaga: Physiological status and susceptibility to chemical inactivation. Appl. Environ. Microbiol. 1992, 58, 2420–2425.
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Abu Kwaik, Y. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277.
- Adékambi, T.; Ben Salah, S.; Khlif, M.; Raoult, D.; Drancourt, M. Survival of Environmental Mycobacteria in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2006, 72, 5974.
- Cervero-Aragó, S.; Rodríguez-Martínez, S.; Puertas-Bennasar, A.; Araujo, R.M. Effect of Common Drinking Water Disinfectants, Chlorine and Heat, on Free Legionella and Amoebae-Associated Legionella. PLoS ONE 2015, 10, e0134726.
- Marciano-Cabral, F.; Jamerson, M.; Kaneshiro, E.S. Free-living amoebae, Legionella and Mycobacterium in tap water supplied by a municipal drinking water utility in the USA. J. Water Health 2009, 8, 71–82.
- Shaheen, M.; Scott, C.; Ashbolt, N.J. Long-term persistence of infectious Legionella with free-living amoebae in drinking water biofilms. Int. J. Hyg. Environ. Health 2019, 222, 678–686.
- Richards, A.M.; Von Dwingelo, J.E.; Price, C.T.; Abu Kwaik, Y. Cellular microbiology and molecular ecology of Legionella–amoeba interaction. Virulence 2013, 4, 307–314.
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S.; Bermudez, L.E. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 1997, 65, 3759–3767.
- González-Cabaleiro, R.; Curtis, T.P.; Ofiţeru, I.D. Bioenergetics analysis of ammonia-oxidizing bacteria and the estimation of their maximum growth yield. Water Res. 2019, 154, 238–245.
- Flannery, B.; Gelling, L.B.; Vugia, D.J.; Weintraub, J.M.; Salerno, J.J.; Conroy, M.J.; Stevens, V.A.; Rose, C.E.; Moore, M.R.; Fields, B.S.; et al. Reducing Legionella colonization of water systems with monochloramine. Emerg. Infect. Dis. 2006, 12, 588.
- Michel, R.; Burghardt, H.; Bergmann, H. Acanthamoeba, naturally intracellularly infected with Pseudomonas aeruginosa, after their isolation from a microbiologically contaminated drinking water system in a hospital. Zent. Hyg. Umweltmed. 1995, 196, 532–544.
- Ji, P.; Rhoads, W.J.; Edwards, M.A.; Pruden, A. Impact of water heater temperature setting and water use frequency on the building plumbing microbiome. Isme J. 2017, 11, 1318–1330.
- Buse, H.Y.; Lu, J.; Lu, X.; Mou, X.; Ashbolt, N.J. Microbial diversities (16S and 18S rRNA gene pyrosequencing) and environmental pathogens within drinking water biofilms grown on the common premise plumbing materials unplasticized polyvinylchloride and copper. FEMS Microbiol. Ecol. 2014, 88, 280–295.
- Lu, J.; Buse, H.Y.; Gomez-Alvarez, V.; Struewing, I.; Santo Domingo, J.; Ashbolt, N.J. Impact of drinking water conditions and copper materials on downstream biofilm microbial communities and Legionella pneumophila colonization. J. Appl. Microbiol. 2014, 117, 905–918.
- Falkinham, J.O.; Hilborn, E.D.; Arduino, M.J.; Pruden, A.; Edwards, M.A. Epidemiology and Ecology of Opportunistic Premise Plumbing Pathogens: Legionella pneumophila, Mycobacterium avium, and Pseudomonas aeruginosa. Environ. Health Perspect. 2015, 123, 749–758.
- Buse, H.Y.; Lu, J.; Struewing, I.T.; Ashbolt, N.J. Preferential colonization and release of Legionella pneumophila from mature drinking water biofilms grown on copper versus unplasticized polyvinylchloride coupons. Int. J. Hyg. Environ. Health 2014, 217, 219–225.
- Ma, X.; Pierre, D.; Bibby, K.; Stout, J.E. Bacterial community structure correlates with Legionella pneumophila colonization of New York City high rise building premises plumbing systems. Environ. Sci. Water Res. Technol. 2020, 6, 1324–1335.
- Paranjape, K.; Bédard, É.; Whyte, L.G.; Ronholm, J.; Prévost, M.; Faucher, S.P. Presence of Legionella spp. in cooling towers: The role of microbial diversity, Pseudomonas, and continuous chlorine application. Water Res. 2020, 169, 115252.
- Garner, E.; McLain, J.; Bowers, J.; Engelthaler, D.M.; Edwards, M.A.; Pruden, A. Microbial Ecology and Water Chemistry Impact Regrowth of Opportunistic Pathogens in Full-Scale Reclaimed Water Distribution Systems. Environ. Sci. Technol. 2018, 52, 9056–9068.
- Ji, P.; Parks, J.; Edwards, M.A.; Pruden, A. Impact of Water Chemistry, Pipe Material and Stagnation on the Building Plumbing Microbiome. PLoS ONE 2015, 10, e0141087.
- Zhang, C.; Li, C.; Zheng, X.; Zhao, J.; He, G.; Zhang, T. Effect of pipe materials on chlorine decay, trihalomethanes formation, and bacterial communities in pilot-scale water distribution systems. Int. J. Environ. Sci. Technol. 2017, 14, 85–94.
- Proctor, C.R.; Dai, D.; Edwards, M.A.; Pruden, A. Interactive effects of temperature, organic carbon, and pipe material on microbiota composition and Legionella pneumophila in hot water plumbing systems. Microbiome 2017, 5, 130.
- Vikesland, P.J.; Valentine, R.L. Reaction Pathways Involved in the Reduction of Monochloramine by Ferrous Iron. Environ. Sci. Technol. 2000, 34, 83–90.
- Lin, W.; Yu, Z.; Chen, X.; Liu, R.; Zhang, H. Molecular characterization of natural biofilms from household taps with different materials: PVC, stainless steel, and cast iron in drinking water distribution system. Appl. Microbiol. Biotechnol. 2012, 97.
- Proctor, C.R.; Gächter, M.; Kötzsch, S.; Rölli, F.; Sigrist, R.; Walser, J.-C.; Hammes, F. Biofilms in shower hoses–choice of pipe material influences bacterial growth and communities. Environ. Sci. Water Res. Technol. 2016, 2, 670–682.
- Corre, M.-H.; Delafont, V.; Legrand, A.; Berjeaud, J.-M.; Verdon, J. Exploiting the Richness of Environmental Waterborne Bacterial Species to Find Natural Legionella pneumophila Competitors. Front. Microbiol. 2019, 9, 3360.
- Kimura, S.; Tateda, K.; Ishii, Y.; Horikawa, M.; Miyairi, S.; Gotoh, N.; Ishiguro, M.; Yamaguchi, K. Pseudomonas aeruginosa Las quorum sensing autoinducer suppresses growth and biofilm production in Legionella species. Microbiology 2009, 155, 1934–1939.
- Declerck, P.; Behets, J.; Delaedt, Y.; Margineanu, A.; Lammertyn, E.; Ollevier, F. Impact of Non-Legionella Bacteria on the Uptake and Intracellular Replication of Legionella pneumophila in Acanthamoeba castellanii and Naegleria lovaniensis. Microb. Ecol. 2005, 50, 536–549.
- Berry, D.; Horn, M.; Xi, C.; Raskin, L. Mycobacterium avium Infections of Acanthamoeba Strains: Host Strain Variability, Grazing-Acquired Infections, and Altered Dynamics of Inactivation with Monochloramine. Appl. Environ. Microbiol. 2010, 76, 6685–6688.
- Weitere, M.; Bergfeld, T.; Rice, S.A.; Matz, C.; Kjelleberg, S. Grazing resistance of Pseudomonas aeruginosa biofilms depends on type of protective mechanism, developmental stage and protozoan feeding mode. Environ. Microbiol. 2005, 7, 1593–1601.
- Pukatzki, S.; Kessin, R.H.; Mekalanos, J.J. The human pathogen Pseudomonas aeruginosa utilizes conserved virulence pathways to infect the social amoeba Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA 2002, 99, 3159.
- Matz, C.; Moreno, A.M.; Alhede, M.; Manefield, M.; Hauser, A.R.; Givskov, M.; Kjelleberg, S. Pseudomonas aeruginosa uses type III secretion system to kill biofilm-associated amoebae. ISME J. 2008, 2, 843–852.
- Steinberg, C.; Laurent, J.; Edel-Hermann, V.; Barbezant, M.; Sixt, N.; Dalle, F.; Aho, S.; Bonnin, A.; Hartemann, P.; Sautour, M. Adaptation of Fusarium oxysporum and Fusarium dimerum to the specific aquatic environment provided by the water systems of hospitals. Water Res. 2015, 76, 53–65.
- Petitjean, M.; Martak, D.; Silvant, A.; Bertrand, X.; Valot, B.; Hocquet, D. Genomic characterization of a local epidemic Pseudomonas aeruginosa reveals specific features of the widespread clone ST395. Microb. Genom. 2017, 3, e000129.
- Bédard, E.; Trigui, H.; Liang, J.; Doberva, M.; Paranjape, K.; Lalancette, C.; Faucher, S.P.; Prévost, M. Local adaptation of Legionella pneumophila within a hospital hot water system increases tolerance to copper. bioRxiv 2020.
- Williams, C.L.; Neu, H.M.; Gilbreath, J.J.; Michel, S.L.J.; Zurawski, D.V.; Merrell, D.S. Copper Resistance of the Emerging Pathogen Acinetobacter baumannii. Appl. Environ. Microbiol. 2016, 82, 6174–6188.
- Lin, Y.-S.E.; Vidic, R.D.; Stout, J.E.; Yu, V.L. Individual and combined effects of copper and silver ions on inactivation of Legionella pneumophila. Water Res. 1996, 30, 1905–1913.
- Mathys, W.; Stanke, J.; Harmuth, M.; Junge-Mathys, E. Occurrence of Legionella in hot water systems of single-family residences in suburbs of two German cities with special reference to solar and district heating. Int. J. Hyg. Environ. Health 2008, 211, 179–185.
- Lin, Y.E.; Vidic, R.D.; Stout, J.E.; Victor, L.Y. Negative effect of high pH on biocidal efficacy of copper and silver ions in controlling Legionella pneumophila. Appl. Environ. Microbiol. 2002, 68, 2711–2715.
- Huang, H.-I.; Shih, H.-Y.; Lee, C.-M.; Yang, T.C.; Lay, J.-J.; Lin, Y.E. In vitro efficacy of copper and silver ions in eradicating Pseudomonas aeruginosa, Stenotrophomonas maltophilia and Acinetobacter baumannii: Implications for on-site disinfection for hospital infection control. Water Res. 2008, 42, 73–80.
- Shih, H.-Y.Y.; Lin, Y.E. Efficacy of Copper-Silver Ionization in Controlling Biofilm- and Plankton-Associated Waterborne Pathogens. Appl. Environ. Microbiol. 2010, 76, 2032–2035.
- Landeen, L.K.; Yahya, M.T.; Kutz, S.M.; Gerba, C.P. Microbiological Evaluation of Copper: Silver Disinfection Units for Use in Swimming Pools. Water Sci. Technol. 1989, 21, 267–270.
- Kusnetsov, J.; Iivanainen, E.; Elomaa, N.; Zacheus, O.; Martikainen, P.J. Copper and silver ions more effective against Legionellae than against mycobacteria in a hospital warm water system. Water Res. 2001, 35, 4217–4225.
- Lin, Y.E.; Vidic, R.D.; Stout, J.E.; McCartney, C.A.; Yu, V.L. Inactivation of Mycobacterium avium by copper and silver ions. Water Res. 1998, 32, 1997–2000.
- Miuetzner, S.; Schwille, R.C.; Farley, A.; Wald, E.R.; Ge, J.H.; States, S.J.; Libert, T.; Wadowsky, R.M. Efficacy of thermal treatment and copper-silver ionization for controlling Legionella pneumophila in high-volume hot water plumbing systems in hospitals. Am. J. Infect. Control 1997, 25, 452–457.
- Cachafeiro, S.P.; Naveira, I.M.; García, I.G. Is copper-silver ionisation safe and effective in controlling legionella? J. Hosp. Infect. 2007, 67, 209–216.
- Mathys, W.; Hohmann, C.P.; Junge-Mathys, E. Efficacy of Copper-Silver Ionization in Controlling Legionella in a Hospital Hot Water Distribution System: A German Experience. In Legionella; Marre, R., Kwaik, Y.A., Bartlett, C., Cianciotto, N.P., Fields, B.S., Frosch, M., Hacker, J., Luck, P.C., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 419–424. ISBN 1-55581-230-9.
- Ramamurthy, T.; Ghosh, A.; Pazhani, G.P.; Shinoda, S. Current Perspectives on Viable but Non-Culturable (VBNC) Pathogenic Bacteria. Front. Public Health 2014, 2, 103.
- Dietersdorfer, E.; Kirschner, A.; Schrammel, B.; Ohradanova-Repic, A.; Stockinger, H.; Sommer, R.; Walochnik, J.; Cervero-Aragó, S. Starved viable but non-culturable (VBNC) Legionella strains can infect and replicate in amoebae and human macrophages. Water Res. 2018, 141, 428–438.
- Williams, K.; Pruden, A.; Falkinham, J.O.; Edwards, M. Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters. Pathogens 2015, 4, 355–372.
- Dusserre, E.; Ginevra, C.; Hallier-Soulier, S.; Vandenesch, F.; Festoc, G.; Etienne, J.; Jarraud, S.; Molmeret, M. A PCR-based method for monitoring Legionella pneumophila in water samples detects viable but noncultivable legionellae that can recover their cultivability. Appl. Environ. Microbiol. 2008, 74, 4817–4824.
- Wang, H.; Bédard, E.; Prévost, M.; Camper, A.K.; Hill, V.R.; Pruden, A. Methodological approaches for monitoring opportunistic pathogens in premise plumbing: A review. Water Res. 2017, 117, 68–86.
- Rowland, J.L.; Niederweis, M. Resistance mechanisms of Mycobacterium tuberculosis against phagosomal copper overload. Tuberculosis 2012, 92, 202–210.
- Gião, M.S.; Wilks, S.A.; Keevil, C.W. Influence of copper surfaces on biofilm formation by Legionella pneumophila in potable water. Biometals 2015, 28, 329.
- Gomes, I.B.; Simões, L.C.; Simões, M. Influence of surface copper content on Stenotrophomonas maltophilia biofilm control using chlorine and mechanical stress. Biofouling 2020, 36, 1–13.
- Bédard, E.; Charron, D.; Lalancette, C.; Déziel, E.; Prévost, M. Recovery of Pseudomonas aeruginosa culturability following copper- and chlorine-induced stress. FEMS Microbiol. Lett. 2014, 356, 226–234.
- Dwidjosiswojo, Z.; Richard, J.; Moritz, M.M.; Dopp, E.; Flemming, H.-C.; Wingender, J. Influence of copper ions on the viability and cytotoxicity of Pseudomonas aeruginosa under conditions relevant to drinking water environments. Int. J. Hyg. Environ. Health 2011, 214, 485–492.
- Dopp, E.; Richard, J.; Dwidjosiswojo, Z.; Simon, A.; Wingender, J. Influence of the copper-induced viable but non-culturable state on the toxicity of Pseudomonas aeruginosa towards human bronchial epithelial cells in vitro. Int. J. Hyg. Environ. Health 2017, 220, 1363–1369.
- Lehtola, M.J.; Miettinen, I.T.; Keinänen, M.M.; Kekki, T.K.; Laine, O.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Microbiology, chemistry and biofilm development in a pilot drinking water distribution system with copper and plastic pipes. Water Res. 2004, 38, 3769–3779.
- Schwan, W.R.; Warrener, P.; Keunz, E.; Kendall Stover, C.; Folger, K.R. Mutations in the cueA gene encoding a copper homeostasis P-type ATPase reduce the pathogenicity of Pseudomonas aeruginosa in mice. Int. J. Med. Microbiol. 2005, 295, 237–242.
- Berg, J.; Tom-Petersen, A.; Nybroe, O. Copper amendment of agricultural soil selects for bacterial antibiotic resistance in the field. Lett. Appl. Microbiol. 2005, 40, 146–151.
- Hu, H.-W.; Wang, J.-T.; Li, J.; Li, J.-J.; Ma, Y.-B.; Chen, D.; He, J.-Z. Field-based evidence for copper contamination induced changes of antibiotic resistance in agricultural soils. Environ. Microbiol. 2016, 18, 3896–3909.
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic Resistance Gene Abundances Correlate with Metal and Geochemical Conditions in Archived Scottish Soils. PLoS ONE 2011, 6, e27300.
- Berg, J.; Thorsen, M.K.; Holm, P.E.; Jensen, J.; Nybroe, O.; Brandt, K.K. Cu Exposure under Field Conditions Coselects for Antibiotic Resistance as Determined by a Novel Cultivation-Independent Bacterial Community Tolerance Assay. Environ. Sci. Technol. 2010, 44, 8724–8728.
- Graham, D.W.; Olivares-Rieumont, S.; Knapp, C.W.; Lima, L.; Werner, D.; Bowen, E. Antibiotic Resistance Gene Abundances Associated with Waste Discharges to the Almendares River near Havana, Cuba. Environ. Sci. Technol. 2011, 45, 418–424.
- Zhang, M.; Chen, L.; Ye, C.; Yu, X. Co-selection of antibiotic resistance via copper shock loading on bacteria from a drinking water bio-filter. Environ. Pollut. 2018, 233, 132–141.
- Becerra-Castro, C.; Machado, R.; Vaz-Moreira, I.; Manaia, C. Assessment of copper and zinc salts as selectors of antibiotic resistance in Gram-negative bacteria. Sci. Total Environ. 2015, 530.
- Barker, J.; Scaife, H.; Brown, M.R. Intraphagocytic growth induces an antibiotic-resistant phenotype of Legionella pneumophila. Antimicrob. Agents Chemother. 1995, 39, 2684.
- Hwang, M.G.; Katayama, H.; Ohgaki, S. Effect of Intracellular Resuscitation of Legionella pneumophila in Acanthamoeba polyphage Cells on the Antimicrobial Properties of Silver and Copper. Environ. Sci. Technol. 2006, 40, 7434–7439.
- World Health Organization. Copper in Drinking-Water; World Health Organization: Geneva, Switzerland, 2016.
- Cervantes, H.I.; Álvarez, J.A.; Muñoz, J.M.; Arreguín, V.; Mosqueda, J.L.; Macías, A.E. Antimicrobial activity of copper against organisms in aqueous solution: A case for copper-based water pipelines in hospitals? Am. J. Infect. Control 2013, 41, e115–e118.
- Oglesby-Sherrouse, A.G.; Djapgne, L.; Nguyen, A.T.; Vasil, A.I.; Vasil, M.L. The complex interplay of iron, biofilm formation, and mucoidy affecting antimicrobial resistance of Pseudomonas aeruginosa. Pathog. Dis. 2014, 70, 307–320.


