 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tanaya Walimbe | + 5096 word(s) | 5096 | 2020-12-08 07:49:07 | | | |
| 2 | Bruce Ren | Meta information modification | 5096 | 2020-12-16 05:17:39 | | |
Video Upload Options
Collagen, the most abundant protein in mammals, has garnered the interest of scientists for over 50 years. Its ubiquitous presence in all body tissues combined with its excellent biocompatibility has led scientists to study its potential as a biomaterial for a wide variety of biomedical applications with a high degree of success and widespread clinical approval. More recently, in order to increase their tunability and applicability, collagen hydrogels have frequently been co-polymerized with other natural and synthetic polymers. Of special significance is the use of bioactive glycosaminoglycans—the carbohydrate-rich polymers of the ECM responsible for regulating tissue homeostasis and cell signaling. This review covers the recent advances in the development of collagen-glycosaminoglycan blend hydrogels for translational biomedical research.
1. Introduction
Universal and ubiquitous in all multicellular organisms—collagen is essential to life as we know it. The triple helical protein not only amounts to the most abundant protein in the human body, but also plays countless roles in tissue structure, cell signaling, and modulation of cell behavior. There are over 29 forms of collagen found in mammalian tissues [1], of which type I, type II, and type III collagen are considered fibrillar collagens. As the main component of all connective tissues, fibrillar collagen accounts for about 55% of the dry mass of skin [2], 25–40% of the dry mass of articular cartilage, 70% of the dry mass of skeletal muscle [3], and 80–90% of the dry mass of bone [4]. It is also abundant in the cornea [5], blood vessels [6], the gut [7], and intervertebral discs [8]. Collagen type 1 is the most abundant fibrillar protein accounting for 90% of the collagen in the body and is composed of monomers containing three polypeptide chains that form a single right-handed triple helical structure [9][10][11]. It is these triple-helical monomers that assemble into collagen fibrils. In the body, collagen fibrils are crosslinked by the enzyme, lysyl oxidase, that acts on the ε-amine groups of lysine residues and supports crosslinks between modified lysine residues and also between modified and other lysine residues within collagen through aldol condensation reactions [12][13]. Crosslinking adds stability to the collagen fibrils and increases collagen tensile strength and resistance to enzymatic degradation [14]. The crosslinked fibers, composed of monomers assembled in a quarter stagger, show a characteristic visual banding pattern with a D-period of 67 nm [15]. Figure 1 shows the structure of collagen and its fibrils. The collagen structure aides in limiting tissue compliance, and also supports interactions with cells and other proteins and glycosaminoglycans. Thus it serves as both a key structural component and signaling component of the extracellular matrix.
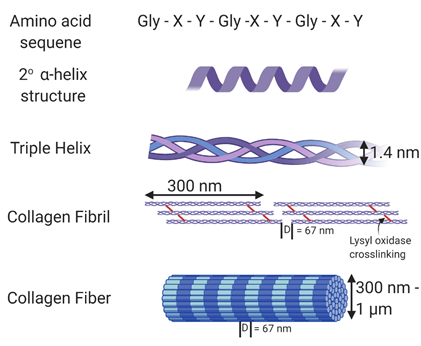
Figure 1. Structure of collagen. The amino acid sequence of collagen consists of Gly-Xaa-Yaa repeats, with Xaa and Yaa commonly occupied by proline and hydroxyproline. This unique sequence allows collagen to form an α helix secondary structure. Fibrillar collagen is a triple helix containing crosslinks formed through the action of lysyl oxidase. In vivo, these collagen fibrils form fibers with varying thickness and a D-banding pattern of 67 nm. Made using Biorender.
The biophysical properties of collagen coupled with the relative abundance and ease of isolation from tissues including skin and tendon make it an interesting and popular biomaterial for tissue engineering, regenerative medicine, and drug delivery. The amino acid sequence of type I collagen consists of Gly-Xaa-Yaa repeats [16], with a substantial amount of Xaa and Yaa repeats consisting of proline (28%) and 4-hydroxyproline (38%) resulting from a post-translational modification of peptide-bound prolyl residues [17][18]. This atypical hydroxyproline content acts as a distinctive marker of collagen and is used in bioassays to identify collagen. In native collagen, the crosslinks generated through the action of lysyl oxidase are primarily found in the telopeptide regions of collagen, and these regions are mostly enzymatically removed from collagen during the process typically used to isolate collagen. As such, polymerized collagen in vitro is generally weak and low-density gels do not sustain their shape in the absence of an external crosslinker. Thus, for many applications researchers either crosslink collagen via one of a host of varying methods or add additional macromolecules to alter the biophysical properties of the hydrogels.
In addition to interacting with biomacromolecules, collagen acts as a ligand for cell receptors. The β1 integrin subfamily is responsible for facilitating binding to collagen. Specifically, α1β1, α2β1, α10β1, and α11β1 receptors bind to the GFOGER sequence in nondenatured collagen type 1 and binding is highly dependent on the presence of the glutamic acid within the ligand sequence[19]. Activation of integrins through collagen binding in turn causes a series of cellular outside-in downstream signaling events. Integrin activation leads to rapid activation of lipid kinases which promote the tyrosine phosphorylation of proteins such as focal adhesion kinase (FAK), p130Cas, and Src [20][21][22]. These then lead to the activation of signaling pathways such as Rho, Rac-1 GTPase as well as cytoskeletal proteins, which drive reorganization of the actin cytoskeleton [23]. These pathway activation responses manifest long term as changes in proliferation, differentiation, migration, and metabolism of cells [23][24]. The integrin-ligand interactions are important in tissue development, healing, and homeostasis as they represent a biophysical connection between the scaffold and the cell.
When collagen is denatured, such as in the form of gelatin, or in areas of damage including wounded or fibrotic tissue, collagen is partially denatured and can support αvβ3 and α5β3 binding via normally inaccessible RGD sequences [25]. Given the importance of integrin activation in cellular processes, variation in activation of integrins can lead to unwanted consequences. In a careful study comparing cell binding and spreading in collagen, collagen and gelatin blends, and gelatin in monolayer form, in 2-dimensions on crosslinked gels, and in 3-dimensional culture, Davidenko, et al. demonstrated the importance of specific integrin binding in cell-collagen interactions. They also demonstrated the altering effect 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) crosslinking can have on these same interactions. As expected, they found that α2β1 and α1β1 integrin–collagen interactions dominated on fibrillar collagen while αvβ3 and α5β3 dominated on gelatin. When cells are cultured in 3D, crosslinking with EDC reduces integrin binding to collagen and gelatin presumably though depletion of critical glutamic and aspartic acid residues within the binding regions. The decreased specific binding also came with increased non-specific interactions that may be due to changes in scaffold mechanics or chemistry [26]. Careful consideration thus needs to be given to integrin–ligand interactions for designing successful tissue-engineered scaffolds.
As researchers uncovered the innumerable roles played by collagen either as a building block or as a fundamental signaling protein, its popularity as a material for tissue engineering for restoring tissue function also surged. The versatility of fibrillar collagen—which could form the backbone of rigid tissues with a high Young’s modulus such as the bone, or provide support to compliant soft tissues with optical transparency such as the corneal stroma, or act as an elastomer with high shock absorbing properties in tissues such as the articular cartilage—truly made it a polymer with endless potential. Capturing these higher order supramolecular structures of collagen in vitro in hydrogels, however, has been challenging. Besides mechanical cues, cells are known to be sensitive to pore size and microstructure and alignment of the collagen fibrils [27][28][29][30]. Alongside, the environment in which collagen is polymerized such as ionic strength, temperature, pH, etc., imparts significant variability to hydrogel formation, structure, and properties [31]. The fact that type I collagen is a hallmark of scar tissue also warrants caution when using collagen to regenerate healthy tissue. This review aims to summarize the current strides made toward optimizing collagen hydrogels for increasing their functionality, tunability, and applicability. We have attempted to highlight the benefits of including additional biopolymers, more specifically, bioactive glycosaminoglycans, within the collagen hydrogels for tissue engineering applications.
2. Collagen–GAG Hydrogels
Early studies with collagen hydrogels including GAGs set the foundation for the importance of GAG-collagen blends in affecting biophysical properties of gels and influencing cell behavior. For example, in 1989, Docherty, et al. reported that inclusion of GAGs in collagen gels supported enhanced fibroblast motility [32]. Further, they shed light on the importance of GAG identity by demonstrating heparin-inhibited collagen polymerization, while low concentrations of HA and CS enhanced cell migration rates, with HA doing so more effectively, while low MW HA was more effective than higher MW HA [33]. In 1996 Bitner et al. demonstrated that decorin altered the ability of fibroblasts to contract collagen gels [34]. These and other studies peaked researchers interest in combined collagen-GAG formulations.
There are six different types of GAGs which are classified based on their disaccharide monomer repeats—hyaluronic acid (HA), heparin, heparan sulfate (HS), chondroitin sulfate (CS), dermatan sulfate (DS), and keratan sulfate (KS) [35]. GAGs are polymer chains composed from disaccharide monomer repeats of glucuronic acid (GlcA) or its epimer iduronic acid (IdoA), and N-acetylgalactosamine (GalNAc) or N-acetylglucosamine (GlcNAc) (Figure 2) [36]. They are post-translationally modified to have variable sulfation patterns that lead to their added complexity and highly negative charge [37]. As with collagen, GAGs are not merely a structural component of the ECM, they also perform a diverse range of functions to modulate tissue homeostasis. GAGs are known for the anti-inflammatory and protective role they play because of their ability to mask cells and other proteins to prevent activation of immune cells—for example, GAGs are a major component of the endothelial glycocalyx, which when stripped, leads to the exposure of ligands that bind and activate neutrophils, monocytes, and platelets [38][39][40]. Additionally, since GAGs are negatively charged, they interact with and attract water to load bearing tissues such as the cartilage and vocal folds, thus increasing their compressive strength and providing shock-absorbing capacity to these tissues [41][42]. This makes them all the more attractive as components of blended hydrogels. Alongside, since they do not contain a protein component, they exhibit minimal antigenicity and demonstrate excellent biocompatibility for tissue engineering purposes. Moreover, modifications can easily be introduced into GAGs through the carboxylic acid group present on the backbone, and more importantly simple modifications can be used to tailor GAGs for specific applications based on their functionality.
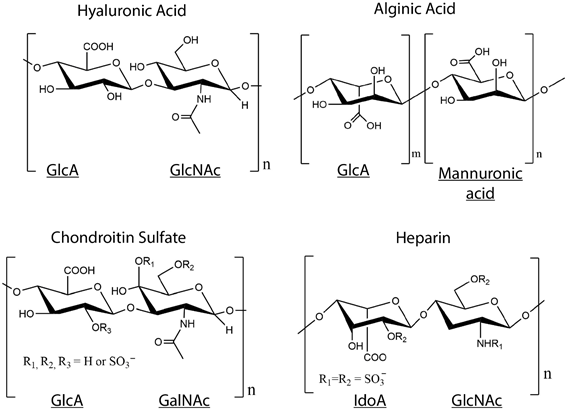
Figure 2. Structures of glycosaminoglycans (GAGs). Disaccharide monomer repeats of glucuronic acid (GlcA) or its epimer iduronic acid (IdoA), and N-acetylgalactosamine (GalNAc) or N-acetylglucosamine (GlcNAc) form the backbone of GAGs, except alginate, which is a GAG analog with GlcA and mannuronic acid repeats. Chondroitin sulfate and heparin are post-translationally sulfated and contain SO3− groups on some disaccharide units.
As with crosslinked collagen hydrogels, parameters such as concentration, crosslinking density, and retention of bioactivity are important fabrication parameters for the design of collagen-GAG hydrogels. High concentrations of GAGs inhibit fibril formation [43], limiting the amount of GAGs that can be added to the hydrogel without negatively affecting the fibril formation. Degree of modification and crosslinking density directly correlate with increased stiffness and stability of the hydrogels, which can be used to design hydrogels to match tissue microenvironments. However, care needs to be taken to not over-substitute GAGs, since higher degrees of modifications result in the loss of bioactivity [44]. Therefore, a balance between concentration, modification degree, and bioactivity needs to be found for the design of GAG-collagen hydrogels. Designing successful hydrogel candidates consequently requires careful understanding of the required tissue outcome and parameters important for regeneration. The next section includes examples of collagen-GAG hydrogels organized by the type of GAG used.
2.1. Collagen–HA Hydrogels
HA is the only non-sulfated GAG and is the most widely used GAG for tissue engineering because of its ease of handling and modification, excellent biocompatibility, diverse molecular weights that can be used to tune its biological and mechanical properties, and high swelling capacity [45]. Hydrogels with HA and collagen do not bind and modulate growth factors to the same extent as those containing sulfated GAG, and in turn, do not affect cell behavior to the same degree. In order to increase the ability of HA-collagen hydrogels to bind growth factors to sustain regeneration, researchers have chemically modified HA to contain sulfate groups and increase its negative charge [46][47][48]. Hydrogels containing sulfated HA could have especially promising applicability for treating chronic wounds and inflammation due to their ability to sequester and release growth factors while modulating cell behavior. Another area of interest is to use these growth factor laden hydrogels to guide stem cell behavior and drive differentiation based on the presentation of growth factors in the hydrogels [49][50][51][52]. Studies have used this approach to guide osteogenic differentiation of MSCs without the use of steroids like dexamethasone . Collagen hydrogels contain modified and unmodified HA have shown great promise as tissue engineering and drug delivery scaffolds.
Thiolated HA is a popular material for hydrogel formation since the thiol groups can be used for both crosslinking and for conjugation of additional functional molecules. Thiolated HA has been extensively used with crosslinked gelatin and poly (ethylene glycol) diacrylate (PEGDA) for close to two decades [53][54]. Other researchers have taken the route of modifying the collagen with methacrylate groups to crosslink with the thiolated HA [55]. However, to avoid the drawbacks inherent to non-fibrillar gelatin and crosslinked collagen, namely the exposure of normally cryptic integrin-binding sequences, the loss of integrin-binding sequences exposed on fibrillar collagen, as well as potential loss of key integrin ligands when crosslinking collagen, our lab designed interpenetrating (IPN) hydrogels with fibrillar collagen type I and type III entrapped within crosslinked thiolated HA. HA crosslinking was achieved using PEGDA, and development of an IPN was achieved by modulating the pH of the hydrogels during polymerization to drive collagen polymerization faster than the Michael type addition between thiols and acrylates [56]. Interpenetrating collagen fibrils could be visualized in the resulting hydrogel, and the gels showed good biocompatibility in culture over 21 days. In this way, tunable hydrogels can be formed without altering the structure of the collagen.
IPNs can also be formed using free radical polymerization. Ultraviolet light (UV)-induced free radical generation offers the advantage of crosslinking acrylated or methacrylated HA after collagen fibrillogenesis is complete to form IPNs without the need to modify the collagen [57][58][59]. Hydrogels synthesized using glycidyl methacrylate HA or methacrylic anhydride modified HA and collagen formed IPNs that slowed degradation of the scaffolds due to UV crosslinks, improved mechanical properties of the scaffolds, and allowed for encapsulation of cells without being cytotoxic. These hydrogels are easy to pattern since the precursor can be patterned and polymerization commenced with exposure to UV light. These gels also provided attachment to various cell types such as Schwann cells and fibroblasts, demonstrating their versatility for biomedical applications. However, the use of a photointiator for UV crosslinking has limited clinical application because of the risk of free radical-induced cell damage and toxicity from the initiators used.
EDC crosslinked HA-collagen hydrogels have been explored for many applications including dermal, corneal, neural, and cartilage tissue engineering, as well as wound healing, etc. Unlike the Collagen HA IPN described above, by using EDC the collagen is crosslinked directly to the carboxylate groups on HA. One advantage of forming collagen-GAG blend hydrogels is the ease with which crosslinker concentration can be used to control hydrogel stiffness while maintaining collagen and GAG biological activity. In an interesting study evaluating the effect of stiffness on stem cell lineage, Her et al. designed HA-collagen scaffolds with stiffness varying from 1 kPa to 10 kPa by varying the amount of EDC-supported crosslinking, and found that they could drive hMSC differentiation toward a neuronal lineage on softer substrates and a glial lineage on the stiff substrates [60]. Murphy et al. found a similar effect while attempting to differentiate MSCs into chondrogenic vs. osteogenic phenotypes, with the amount and crosslinking of GAGs to the collagen influencing outcomes [61]. Other studies have also shown that the addition of HA to collagen scaffolds improves chondrogenic differentiation of MSCs [62][63][64], which is not surprising, given that the articular cartilage consists of ECM rich in GAGs and collagen for shock absorption and mechanical loading properties.
Because all GAGs contain carboxylate groups that can be activated by EDC and other agents for crosslinking to free amines on collagen fibrils, it is possible to form collagen hydrogels with blends of GAGs. For example, in studies aimed at designing scaffolds for dermal repair, Wang et al. crosslinked HA and CS with collagen using EDC to form nine different blends with varying ratios of collagen:HA:CS [65]. Scaffolds seeded with allogenic fibroblasts in a 9:1:1 blend of crosslinked Col:HA:CS showed almost normal skin 6 weeks after implantation in Sprague-Drawley rats in comparison to controls, indicating their potential for regenerating skin tissue. GAGs can also be used to control the optical properties of collagen gels, which when polymerized alone are not optically transparent. In another study, EDC/NHS crosslinking resulted in collagen-gelatin-HA films with optical performance, hydrophilicity, and mechanical properties suitable for corneal tissue engineering [66]. Further, these same gels were used to design a wound-healing scaffold for chronic wounds, since EDC crosslinking increases biological stability by decreasing enzymatic degradation in the highly proteolytic chronic wound environment [67]. Collectively, these studies demonstrate the importance of modulating the degree of crosslinking and biological activity to achieve intended biological outcomes.
One limitation to the EDC crosslinked materials described above is the development of largely elastic hydrogels. To circumvent problems associated with these purely elastic materials, the Chaudhuri group designed IPNs of HA and collagen by mixing HA-hydrazine, collagen, and HA-aldehyde (or benzaldehyde) solutions to form stress relaxation hydrogels that contain dynamic covalent bonds [68]. They showed that stress relaxation promoted cell spreading, fiber remodeling, and focal adhesion formation in 3D, and could guide mechanotransduction and recapitulate the fibrillar collagen architecture of many cellular microenvironments.
While EDC is a popular choice to induce crosslinking, other agents have also been studied to crosslink GAG-collagen gels. GAGs, including HA can be readily modified to contain aldehyde groups, however reactions between aldehydes and amines results in unstable crosslinks. Oxime reactions have the advantage of being more stable than Schiff bases formed between aldehydes or ketones and amines, while providing a similar level of tunability. Hardy and Schmidt used oxime-bond click chemistry to couple aldehyde group containing HA with aminooxy-terminated poly (ethylene glycol) (PEG), and further incorporated collagen type I in these gels to provide adhesion sites for MSCs to aid neural tissue regeneration [69]. The chemistry was not toxic to primary Schwann cells, and the hydrogels could be tuned to have mechanical properties analogous to those found in soft tissues such as the central and peripheral nervous system, demonstrating the wide breadth of tissues HA-collagen hydrogels can be tuned towards. Some studies have also attempted to use adipic acid dihydrazide (AAD)-modified HA to crosslink with collagen hydrogels. Genepin crosslinking of AAD modified HA and collagen have shown increased stability and retention of HA in these composite scaffolds [70], possibly because of slowing of degradation due to the crosslinked nature of the hydrogels.
Overall, this vast array of studies shows that the addition of crosslinked HA improved handling and biological outcome of the hydrogels over those seen with collagen alone. Collagen-HA hydrogels have wide-ranging applicability for tissue engineering due to their superior ability to be modulated based on the target tissue biophysical properties. As with any crosslinked gel, the inverse correlation between bioactivity and tunability will dictate the efficacy of the scaffolds for tissue engineering applications. The added complication of balancing hydrophilicity and hydrophobicity to ensure polymerization of collagen in the water-loving HA provides some design challenges that need careful consideration, since a high concentration of GAGs, including HA, will inhibit polymerization of collagen. Therefore, along with degree of modification, concentrations of the collagen and GAGs along with the pH of the solution will be crucial in designing successful constructs with the desired biophysical properties. The next generation of treatments will potentially involve the use of these engineered matrices in conjugation with growth factors and cells for sustained regeneration of tissues.
2.2. Collagen–CS Hydrogels
As the most abundant GAG in the body, CS is found in the vitreous fluid, the glycocalyx and other connective tissues, but is found in highest concentrations in the cartilage. It is a sulfated GAG that is known for its shock absorbing properties, maintenance of cartilage structure and function, and prevention of inflammation, due to which it has been extensively used for the treatment of osteoarthritis [71][72][73].
Like with HA, the use of EDC/NHS chemistry has been routinely performed to crosslink CS to collagen for hydrogel formation due to its low cytotoxicity and ease of handling. However, even with its extensive use, crosslinking Collagen-GAG hydrogels with EDC is not trivial, especially when the goal is to form interpenetrating networks. The van Kuppevelt group conducted extensive studies to optimize crosslinked CS-collagen based hydrogels. In an initial study, two different types of crosslinking techniques, namely, DHT and EDC crosslinking were investigated as a means to incorporate CS in type I collagen hydrogels [74]. EDC crosslinking of CS with collagen pre-crosslinked using DHT treatment resulted in collapsed matrices with CS decorating only the exterior of the collagen, possibly due to the already crosslinked and most probably denatured nature of the collagen. EDC crosslinking to non-crosslinked collagen under aqueous conditions also led to partial collapse of the matrix, which could be circumvented in the presence of ethanol. The addition of an organic solvent such as ethanol likely suppresses urea forming side reactions that commonly occur in water and promote amide bond formation between collagen and GAGs, resulting in evenly distributed crosslinked hydrogels. However, the addition of ethanol precludes the polymerization of these scaffolds in the presence of cells in vitro and limits that potential to implant cell-embedded scaffolds in vivo. They were however able to test the bovine type I collagen with CS or HS crosslinked using EDC-NHS in the presence of ethanol in rats (post ethanol removal) to evaluate tissue response to these hydrogels. The addition of GAGs reduced foreign body reactions, which in turn reduced degradation of the scaffolds in comparison to purely collagen implants. The highly negative charged scaffolds containing GAGs also led to enhanced angiogenesis, possibly through sequestration of growth factors such as FGF, vascular endothelial growth factor (VEGF), platelet derived growth factor (PDGF), etc. [75].
When the goal is not to form an IPN, but a CS-collagen gel, EDC crosslinking can be beneficial. For example, Gao et al., used CS activated with EDC NHS (CS-sNHS) to form crosslinked hydrogels with type II collagen to use as a cell delivery system to treat defects in the articular cartilage. Crosslinking type II collagen with the CS is an innovative way to get type II collagen to stay in the hydrogel, since it is not capable of forming strong fibrils spontaneously in vitro. The scaffolds showed good biocompatibility and were tunable depending on the amount of CS-sNHS added. Several additional studies have demonstrated improved outcomes with the use of collagen-CS scaffolds for cartilage tissue engineering. While these studies approached the incorporation of CS in collagen hydrogels using different crosslinking methods such as via methacrylated CS or using genipin, the end goal was similar-to promote chondrogenesis and tune the mechanical properties of these hydrogels to suit articular cartilage tissue engineering . The next generation of cartilage therapy will likely include hydrogels with encapsulated cells for superior and sustained regeneration of the cartilage.
EDC/NHS crosslinked CS-Collagen 3D sponges have also been used for applications including neural tissue engineering applications and embryonic neural cell culture, and showed superior thermal resistance and lower enzyme sensitivity than collagen alone [76][77][78]. This shows the flexibility and promise of CS-Collagen systems for tissue regeneration.
It is also possible to incorporate GAGs into collagen in the absence of additional chemical crosslinking. Stuart and Panitch demonstrated that the addition of CS to collagen type I led to network organization with an increase in void space and decreased stiffness of the gels [78]. Since type II and type III collagen are also a major ECM component in some tissues such as the articular cartilage and vocal folds, researchers have sought to incorporate these fibrillar collagens in an attempt to make more biologically relevant hydrogels. Studies in our lab have shown that polymerizing type I and type III collagen together with CS leads to a more open and compliant hydrogel network [79].
CS–collagen hydrogels thus provide an exciting class of hydrogels which can be used to reduce collagen compaction, modulate cell response, and drive differentiation of stem cells while allowing the same flexible level of tunability offered by collagen–HA hydrogels. Taken together, the benefits of adding CS to collagen with respect to amplifying the bioactivity and cell instructive nature of the hydrogels further suggests that the addition of GAGs to collagen can improve tissue outcomes.
2.3. Collagen-Heparin Gels
Unfractionated heparin is best known for its anticoagulant properties and commonly used as a blood thinner [80]. It is the most negatively charged GAG, and thus demonstrates binding and sequestering of growth factors along with being anti-inflammatory. This ability can be exploited to design bioinstructive hydrogels that are capable of providing sustained release of growth factors along with the benefits of the biocompatible materials themselves. Copes et al. exploited this ability of heparin to engineer collagen–heparin hydrogels for the controlled release of pleitropin for vascular applications [79]. Though studies have shown that high concentrations of heparin can inhibit collagen fibrillogenensis [81][82], no effect of heparin on collagen fibril formation was seen in this study. This strategy of providing cell instructive cues and entrapping growth factors can be useful for sustained release of many growth factors to provide persistent regeneration [83]. Another attractive area of application is to guide controlled differentiation of stem cells by sequestering relevant growth factors for augmenting tissues[84].
In other uses, maleimide functionalized heparin linking to star-PEG, has been used with collagen to form hydrogels containing cell instructive peptides as an innovative way to combine semi-synthetic and native ECM molecules to closely mimic the ECM [85]. Modifying the heparin to add functionalities can similarly be applied in the future to include cues for regeneration and repair. While unfractionated heparin is anticoagulating, low molecular weight heparins can be substituted to pivot to applications where anticoagulation is not a desirable outcome. Combined with the growth factor sequestering ability of heparin, this creates additional exciting venues for regenerative medicine applications.
2.4. Collagen-Alginate Gels
Alginate is a naturally occurring anionic carbohydrate polymer obtained from seaweed. While not bioactive like other GAGs and not a traditional component of the ECM, alginate has been widely used for tissue engineering because of its biocompatibility, low toxicity, relatively low cost, and tunability. Sodium alginate forms a hydrogel in aqueous solutions by divalent cation binding [86]. This makes it a highly versatile and tunable GAG analog that can be used as a building block in hydrogels. Another major advantage is that the degree of alginate crosslinking will not affect collagen adhesion sites, providing a way to engineer tunable scaffolds without significantly affecting the bioactivity of collagen fibrils.
To model in vitro tumor environments that offer flexible control of mechanical and biophysical features, Liu et al. engineered composite alginate-collagen hydrogels. Hybrid alginate-collagen hydrogels can be tuned to demonstrate a wide range of elastic moduli by changing the concentration of CaCl2 crosslinker, and used to model tumor invasion of breast cancer cells [87]. Moxon et al. blended collagen with alginate to mimic the hyaluronic acid and collagen-rich environment of the brain ECM [88]. The substrate was conducive to the growth of human iPSC-derived neurons, and tunable based on the amount of ionic crosslinker added for gelation of alginate. Comparing different GAG blends with collagen for vocal fold tissue engineering, collagen-alginate composite hydrogels were shown to resist scaffold compaction and mass loss for at least 42 days in culture while allowing for ECM synthesis in comparison to collagen-HA composite hydrogels, thus showing that the type of GAG used can influence not only gel properties, but also cellular outcome [89].
Several studies have used collagen-alginate hydrogels as biocompatible substrates for chondrocytes geared toward tissue engineering of the articular cartilage [90][91]. Yang et al. employed 3D bioprinting to precisely print collagen-alginate gels for articular cartilage, since bioprinting can provide improved control over spatial and architectural orientation [92]. The high water retention capacity and reduction of gel contraction in composite collagen-alginate hydrogels makes it an attractive hydrogel for articular cartilage tissue engineering in comparison to collagen alone.
In the space of drug delivery, alginate hydrogels have been used as carriers for combinatorial photothermal and immuno tumor therapy by simultaneous encapsulation of the photothermal drug methylene blue (MB) and immunological agent imiquimod (R837) for prolonged and sustained delivery [93]. In a separate study, Lee et al., encapsulated glial cell line-derived neurotrophic factor (GDNF) secreting HEK293 cells in collagen-alginate microspheres for the controlled release of GDNF. They further went on to test these cell encapsulated microspheres for the treatment of neurodegenerative posterior eye diseases in a rat model and showed promising results with improved photoreceptor survival in dystrophic rat eyes [94].
While not a native GAG, alginate continues to grow in popularity because of its mild gelation conditions, biocompatibility, and tunability. Unfortunately, release of the divalent ions responsible for crosslinking the gel causes alginate gels to dissolve, limiting the long-term stability of alginate-collagen gels in physiological conditions. This can be beneficial or negative depending on the application, but needs to taken into consideration before designing the hydrogels. Alongside, alginate plays a passive role in the hydrogels by not modulating signaling directly. Collagen-alginate hydrogels thus make for attractive carriers for drug delivery. Future studies will likely look at incorporating bioactive factors and targeting peptides to increase alginate functionality for drug delivery applications.
3. Conclusions and Future Perspectives
The addition of GAGs to collagen hydrogels has expanded the possibilities for the application of collagen hydrogels from tissue engineering to drug delivery. GAGs not only act as a tunable component of the gels, but also represent a more physiologically relevant ECM biomimic that is capable of modulating collagen fibrillogenesis, hydrogel properties and swelling, as well as guide cell behavior. Strides have been made in incorporating GAGs and collagen in spatially controlled hydrogels and innovative crosslinking strategies have allowed the preservation of fibrillar collagen in hydrogels that are still tunable without modification of the collagen. Based on the type of GAG used, different functionalities can be exploited to engineer biomimetics that closely resemble the tissue microenvironment in vivo, as well as allow for sustained release of growth factors and provide molecular cues that guide regeneration. Combinatorial approaches to biomaterials can thus provide higher control of cell function as well as open an array of new treatments for tackling complicated diseases. This review attempts to show examples of how far blended hydrogels of collagen and have come, but does not touch upon other blends of collagen hydrogels such as those blended with synthetic polymers or other natural polymers such as fibrin due to space constraints. One thing that does stand out, however, is that researchers may want to consider using a combination of not only biomaterials such as collagen and GAGs, but also cells and growth factors in order to modulate biophysical cell–gel interactions and develop successful efficacious treatments for regenerative medicine and drug delivery.
References
- Gorgieva, S.; Kokol, V. Collagen- vs. Gelatine-Based Biomaterials and Their Biocompatibility: Review and Perspectives. Biomater. Appl. Nanomed. 2011, doi:10.5772/24118.
- Lovell, C.R.; Smolenski, K.A.; Duance, V.C.; Light, N.D.; Young, S.; Dyson, M. Type I and III collagen content and fibre distribution in normal human skin during ageing. Br. J. Dermatol. 1987, 117, 419–428, doi:10.1111/j.1365-2133.1987.tb04921.x.
- McKee, T.J.; Perlman, G.; Morris, M.; Komarova, S.V. Extracellular matrix composition of connective tissues: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 10542, doi:10.1038/s41598-019-46896-0.
- Boskey, A.L.; Robey, P.G. The Composition of Bone. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism; American Society for Bone and Mineral Research: Washington, DC, USA, 2018; pp. 84–92.
- Meek, K.M. Corneal collagen-its role in maintaining corneal shape and transparency. Biophys. Rev. 2009, 1, 83–93, doi:10.1007/s12551-009-0011-x.
- Rhodes, J.M.; Simons, M. The extracellular matrix and blood vessel formation: Not just a scaffold. J. Cell Mol. Med. 2007, 11, 176–205, doi:10.1111/j.1582-4934.2007.00031.x.
- Komuro, T. The lattice arrangement of the collagen fibres in the submucosa of the rat small intestine: Scanning electron microscopy. Cell Tissue Res. 1988, 251, 117–121, doi:10.1007/BF00215455.
- Sharabi, M.; Wade, K.; Haj-Ali, R. The Mechanical Role of Collagen Fibers in the Intervertebral Disc. In Biomechanics of the Spine; Galbusera, F., Wilke, H.-J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 105–123.
- Ramachandran, G.N.; Kartha, G. Structure of collagen. Nature 1954, 174, 269–270, doi:10.1038/174269c0.
- Rich, A.; Crick, F.H. The structure of collagen. Nature 1955, 176, 915–916, doi:10.1038/176915a0.
- Cowan, P.M.; McGavin, S.; North, A.C. The polypeptide chain configuration of collagen. Nature 1955, 176, 1062–1064, doi:10.1038/1761062a0.
- Pinnell, S.R.; Martin, G.R. The cross-linking of collagen and elastin: Enzymatic conversion of lysine in peptide linkage to alpha-aminoadipic-delta-semialdehyde (allysine) by an extract from bone. Proc. Natl. Acad. Sci. USA 1968, 61, 708–716, doi:10.1073/pnas.61.2.708.
- Eyre, D.R.; Paz, M.A.; Gallop, P.M. Cross-linking in collagen and elastin. Annu. Rev. Biochem. 1984, 53, 717–748, doi:10.1146/annurev.bi.53.070184.003441.
- Davidenko, N.; Schuster, C.F.; Bax, D.V.; Raynal, N.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Control of crosslinking for tailoring collagen-based scaffolds stability and mechanics. Acta Biomater. 2015, 25, 131–142, doi:10.1016/j.actbio.2015.07.034.
- Meek, K.M.; Chapman, J.A.; Hardcastle, R.A. The staining pattern of collagen fibrils. Improved correlation with sequence data. J. Biol. Chem. 1979, 254, 10710–10714.
- Fietzek, P.P.; Kuhn, K. Information contained in the amino acid sequence of the alpha1(I)-chain of collagen and its consequences upon the formation of the triple helix, of fibrils and crosslinks. Mol. Cell Biochem. 1975, 8, 141–157, doi:10.1007/BF01792765.
- Berg, R.A.; Prockop, D.J. The thermal transition of a non-hydroxylated form of collagen. Evidence for a role for hydroxyproline in stabilizing the triple-helix of collagen. Biochem. Biophys. Res. Commun. 1973, 52, 115–120, doi:10.1016/0006-291x(73)90961-3.
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958, doi:10.1146/annurev.biochem.77.032207.120833.
- Knight, C.G.; Morton, L.F.; Peachey, A.R.; Tuckwell, D.S.; Farndale, R.W.; Barnes, M.J. The collagen-binding A-domains of integrins alpha(1)beta(1) and alpha(2)beta(1) recognize the same specific amino acid sequence, GFOGER, in native (triple-helical) collagens. J. Biol. Chem. 2000, 275, 35–40, doi:10.1074/jbc.275.1.35.
- Legate, K.R.; Wickstrom, S.A.; Fassler, R. Genetic and cell biological analysis of integrin outside-in signaling. Genes Dev. 2009, 23, 397–418, doi:10.1101/gad.1758709.
- Guan, J.L. Role of focal adhesion kinase in integrin signaling. Int. J. Biochem. Cell Biol. 1997, 29, 1085–1096, doi:10.1016/s1357-2725(97)00051-4.
- Moraes, J.A.; Frony, A.C.; Dias, A.M.; Renovato-Martins, M.; Rodrigues, G.; Marcinkiewicz, C.; Assreuy, J.; Barja-Fidalgo, C. Alpha1beta1 and integrin-linked kinase interact and modulate angiotensin II effects in vascular smooth muscle cells. Atherosclerosis 2015, 243, 477–485, doi:10.1016/j.atherosclerosis.2015.09.026.
- Heino, J. Cellular Signaling by Collagen-Binding Integrins. In I Domain Integrins; Gullberg, D., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 143–155, doi:10.1007/978-94-017-9153-3_10.
- Wu, D.; Witt, R.L.; Harrington, D.A.; Farach-Carson, M.C. Dynamic Assembly of Human Salivary Stem/Progenitor Microstructures Requires Coordinated alpha1beta1 Integrin-Mediated Motility. Front. Cell Dev. Biol. 2019, 7, 224, doi:10.3389/fcell.2019.00224.
- Davis, G.E. Affinity of integrins for damaged extracellular matrix: Alpha v beta 3 binds to denatured collagen type I through RGD sites. Biochem. Biophys. Res. Commun. 1992, 182, 1025–1031, doi:10.1016/0006-291x(92)91834-d.
- Davidenko, N.; Schuster, C.F.; Bax, D.V.; Farndale, R.W.; Hamaia, S.; Best, S.M.; Cameron, R.E. Evaluation of cell binding to collagen and gelatin: A study of the effect of 2D and 3D architecture and surface chemistry. J. Mater. Sci. Mater. Med. 2016, 27, 148, doi:10.1007/s10856-016-5763-9.
- Sapudom, J.; Rubner, S.; Martin, S.; Kurth, T.; Riedel, S.; Mierke, C.T.; Pompe, T. The phenotype of cancer cell invasion controlled by fibril diameter and pore size of 3D collagen networks. Biomaterials 2015, 52, 367–375, doi:10.1016/j.biomaterials.2015.02.022.
- Roeder, B.A.; Kokini, K.; Voytik-Harbin, S.L. Fibril microstructure affects strain transmission within collagen extracellular matrices. J. Biomech. Eng. 2009, 131, 031004, doi:10.1115/1.3005331.
- Corin, K.A.; Gibson, L.J. Cell contraction forces in scaffolds with varying pore size and cell density. Biomaterials 2010, 31, 4835–4845, doi:10.1016/j.biomaterials.2010.01.149.
- Ray, A.; Morford, R.K.; Ghaderi, N.; Odde, D.J.; Provenzano, P.P. Dynamics of 3D carcinoma cell invasion into aligned collagen. Integr. Biol. 2018, 10, 100–112, doi:10.1039/c7ib00152e.
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Review of collagen I hydrogels for bioengineered tissue microenvironments: Characterization of mechanics, structure, and transport. Tissue Eng. Part B Rev. 2014, 20, 683–696, doi:10.1089/ten.TEB.2014.0086.
- Docherty, R.; Forrester, J.V.; Lackie, J.M.; Gregory, D.W. Glycosaminoglycans facilitate the movement of fibroblasts through three-dimensional collagen matrices. J. Cell Sci. 1989, 92, 263–270.
- Bittner, K.; Liszio, C.; Blumberg, P.; Schonherr, E.; Kresse, H. Modulation of collagen gel contraction by decorin. Biochem. J. 1996, 314, 159–166, doi:10.1042/bj3140159.
- Casale, J.; Crane, J.S. Biochemistry, Glycosaminoglycans. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020.
- Gandhi, N.S.; Mancera, R.L. The structure of glycosaminoglycans and their interactions with proteins. Chem. Biol. Drug Des. 2008, 72, 455–482, doi:10.1111/j.1747-0285.2008.00741.x.
- Sasisekharan, R.; Venkataraman, G. Heparin and heparan sulfate: Biosynthesis, structure and function. Curr. Opin. Chem. Biol. 2000, 4, 626–631, doi:10.1016/s1367-5931(00)00145-9.
- Alphonsus, C.S.; Rodseth, R.N. The endothelial glycocalyx: A review of the vascular barrier. Anaesthesia 2014, 69, 777–784, doi:10.1111/anae.12661.
- Masola, V.; Zaza, G.; Onisto, M.; Lupo, A.; Gambaro, G. Glycosaminoglycans, proteoglycans and sulodexide and the endothelium: Biological roles and pharmacological effects. Int. Angiol. 2014, 33, 243–254.
- Chappell, D.; Brettner, F.; Doerfler, N.; Jacob, M.; Rehm, M.; Bruegger, D.; Conzen, P.; Jacob, B.; Becker, B.F. Protection of glycocalyx decreases platelet adhesion after ischaemia/reperfusion: An animal study. Eur. J. Anaesthesiol. 2014, 31, 474–481, doi:10.1097/EJA.0000000000000085.
- Glant, T.T.; Buzas, E.I.; Finnegan, A.; Negroiu, G.; Cs-Szabo, G.; Mikecz, K. Critical roles of glycosaminoglycan side chains of cartilage proteoglycan (aggrecan) in antigen recognition and presentation. J. Immunol. 1998, 160, 3812–3819.
- Walimbe, T.; Panitch, A.; Sivasankar, P.M. A Review of Hyaluronic Acid and Hyaluronic Acid-based Hydrogels for Vocal Fold Tissue Engineering. J. Voice 2017, 31, 416–423, doi:10.1016/j.jvoice.2016.11.014.
- Parry, D.A.D.; Flint, M.H.; Gillard, G.C.; Craig, A.S. A role for glycosaminoglycans in the development of collagen fibrils. FEBS Lett. 1982, 149, 1–7, doi:10.1016/0014-5793(82)81060-0.
- Eng, D.; Caplan, M.; Preul, M.; Panitch, A. Hyaluronan scaffolds: A balance between backbone functionalization and bioactivity. Acta Biomater. 2010, 6, 2407–2414, doi:10.1016/j.actbio.2009.12.049.
- Burdick, J.A.; Prestwich, G.D. Hyaluronic acid hydrogels for biomedical applications. Adv. Mater. 2011, 23, H41–H56, doi:10.1002/adma.201003963.
- Thones, S.; Rother, S.; Wippold, T.; Blaszkiewicz, J.; Balamurugan, K.; Moeller, S.; Ruiz-Gomez, G.; Schnabelrauch, M.; Scharnweber, D.; Saalbach, A.; et al. Hyaluronan/collagen hydrogels containing sulfated hyaluronan improve wound healing by sustained release of heparin-binding EGF-like growth factor. Acta Biomater. 2019, 86, 135–147, doi:10.1016/j.actbio.2019.01.029.
- Rother, S.; Galiazzo, V.D.; Kilian, D.; Fiebig, K.M.; Becher, J.; Moeller, S.; Hempel, U.; Schnabelrauch, M.; Waltenberger, J.; Scharnweber, D.; et al. Hyaluronan/Collagen Hydrogels with Sulfated Hyaluronan for Improved Repair of Vascularized Tissue Tune the Binding of Proteins and Promote Endothelial Cell Growth. Macromol. Biosci. 2017, 17, doi:10.1002/mabi.201700154.
- Rother, S.; Kronert, V.; Hauck, N.; Berg, A.; Moeller, S.; Schnabelrauch, M.; Thiele, J.; Scharnweber, D.; Hintze, V. Hyaluronan/collagen hydrogel matrices containing high-sulfated hyaluronan microgels for regulating transforming growth factor-beta1. J. Mater. Sci. Mater. Med. 2019, 30, 65, doi:10.1007/s10856-019-6267-1.
- Hempel, U.; Hintze, V.; Moller, S.; Schnabelrauch, M.; Scharnweber, D.; Dieter, P. Artificial extracellular matrices composed of collagen I and sulfated hyaluronan with adsorbed transforming growth factor beta1 promote collagen synthesis of human mesenchymal stromal cells. Acta Biomater. 2012, 8, 659–666, doi:10.1016/j.actbio.2011.10.026.
- Hempel, U.; Moller, S.; Noack, C.; Hintze, V.; Scharnweber, D.; Schnabelrauch, M.; Dieter, P. Sulfated hyaluronan/collagen I matrices enhance the osteogenic differentiation of human mesenchymal stromal cells in vitro even in the absence of dexamethasone. Acta Biomater. 2012, 8, 4064–4072, doi:10.1016/j.actbio.2012.06.039.
- Kliemt, S.; Lange, C.; Otto, W.; Hintze, V.; Moller, S.; von Bergen, M.; Hempel, U.; Kalkhof, S. Sulfated hyaluronan containing collagen matrices enhance cell-matrix-interaction, endocytosis, and osteogenic differentiation of human mesenchymal stromal cells. J. Proteome Res. 2013, 12, 378–389, doi:10.1021/pr300640h.
- Hempel, U.; Matthaus, C.; Preissler, C.; Moller, S.; Hintze, V.; Dieter, P. Artificial matrices with high-sulfated glycosaminoglycans and collagen are anti-inflammatory and pro-osteogenic for human mesenchymal stromal cells. J. Cell Biochem. 2014, 115, 1561–1571, doi:10.1002/jcb.24814.
- Shu, X.Z.; Liu, Y.; Palumbo, F.; Prestwich, G.D. Disulfide-crosslinked hyaluronan-gelatin hydrogel films: A covalent mimic of the extracellular matrix for in vitro cell growth. Biomaterials 2003, 24, 3825–3834, doi:10.1016/s0142-9612(03)00267-9.
- Wirostko, B.; Mann, B.K.; Williams, D.L.; Prestwich, G.D. Ophthalmic Uses of a Thiol-Modified Hyaluronan-Based Hydrogel. Adv. Wound Care 2014, 3, 708–716, doi:10.1089/wound.2014.0572.
- Mazzocchi, A.; Devarasetty, M.; Huntwork, R.; Soker, S.; Skardal, A. Optimization of collagen type I-hyaluronan hybrid bioink for 3D bioprinted liver microenvironments. Biofabrication 2018, 11, 015003, doi:10.1088/1758-5090/aae543.
- Walimbe, T.; Calve, S.; Panitch, A.; Sivasankar, M.P. Incorporation of types I and III collagen in tunable hyaluronan hydrogels for vocal fold tissue engineering. Acta Biomater. 2019, 87, 97–107, doi:10.1016/j.actbio.2019.01.058.
- Suri, S.; Schmidt, C.E. Photopatterned collagen-hyaluronic acid interpenetrating polymer network hydrogels. Acta Biomater. 2009, 5, 2385–2397, doi:10.1016/j.actbio.2009.05.004.
- Brigham, M.D.; Bick, A.; Lo, E.; Bendali, A.; Burdick, J.A.; Khademhosseini, A. Mechanically robust and bioadhesive collagen and photocrosslinkable hyaluronic acid semi-interpenetrating networks. Tissue Eng. Part A 2009, 15, 1645–1653, doi:10.1089/ten.tea.2008.0441.
- Suri, S.; Schmidt, C.E. Cell-laden hydrogel constructs of hyaluronic acid, collagen, and laminin for neural tissue engineering. Tissue Eng. Part A 2010, 16, 1703–1716, doi:10.1089/ten.tea.2009.0381.
- Her, G.J.; Wu, H.C.; Chen, M.H.; Chen, M.Y.; Chang, S.C.; Wang, T.W. Control of three-dimensional substrate stiffness to manipulate mesenchymal stem cell fate toward neuronal or glial lineages. Acta Biomater. 2013, 9, 5170–5180, doi:10.1016/j.actbio.2012.10.012.
- Murphy, C.M.; Matsiko, A.; Haugh, M.G.; Gleeson, J.P.; O’Brien, F.J. Mesenchymal stem cell fate is regulated by the composition and mechanical properties of collagen-glycosaminoglycan scaffolds. J. Mech. Behav. Biomed. Mater. 2012, 11, 53–62, doi:10.1016/j.jmbbm.2011.11.009.
- Matsiko, A.; Levingstone, T.J.; O’Brien, F.J.; Gleeson, J.P. Addition of hyaluronic acid improves cellular infiltration and promotes early-stage chondrogenesis in a collagen-based scaffold for cartilage tissue engineering. J. Mech. Behav. Biomed. Mater. 2012, 11, 41–52, doi:10.1016/j.jmbbm.2011.11.012.
- Koh, R.H.; Jin, Y.; Kang, B.J.; Hwang, N.S. Chondrogenically primed tonsil-derived mesenchymal stem cells encapsulated in riboflavin-induced photocrosslinking collagen-hyaluronic acid hydrogel for meniscus tissue repairs. Acta Biomater. 2017, 53, 318–328, doi:10.1016/j.actbio.2017.01.081.
- Heo, J.; Koh, R.H.; Shim, W.; Kim, H.D.; Yim, H.G.; Hwang, N.S. Riboflavin-induced photo-crosslinking of collagen hydrogel and its application in meniscus tissue engineering. Drug Deliv. Transl. Res. 2016, 6, 148–158, doi:10.1007/s13346-015-0224-4.
- Wang, W.; Zhang, M.; Lu, W.; Zhang, X.; Ma, D.; Rong, X.; Yu, C.; Jin, Y. Cross-linked collagen-chondroitin sulfate-hyaluronic acid imitating extracellular matrix as scaffold for dermal tissue engineering. Tissue Eng. Part C Methods 2010, 16, 269–279, doi:10.1089/ten.TEC.2009.0161.
- Liu, Y.; Ren, L.; Wang, Y. Crosslinked collagen-gelatin-hyaluronic acid biomimetic film for cornea tissue engineering applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2013, 33, 196–201, doi:10.1016/j.msec.2012.08.030.
- Kirk, J.F.; Ritter, G.; Finger, I.; Sankar, D.; Reddy, J.D.; Talton, J.D.; Nataraj, C.; Narisawa, S.; Millan, J.L.; Cobb, R.R. Mechanical and biocompatible characterization of a cross-linked collagen-hyaluronic acid wound dressing. Biomatter 2013, 3, doi:10.4161/biom.25633.
- Lou, J.; Stowers, R.; Nam, S.; Xia, Y.; Chaudhuri, O. Stress relaxing hyaluronic acid-collagen hydrogels promote cell spreading, fiber remodeling, and focal adhesion formation in 3D cell culture. Biomaterials 2018, 154, 213–222, doi:10.1016/j.biomaterials.2017.11.004.
- Hardy, J.G.; Lin, P.; Schmidt, C.E. Biodegradable hydrogels composed of oxime crosslinked poly(ethylene glycol), hyaluronic acid and collagen: A tunable platform for soft tissue engineering. J. Biomater. Sci. Polym. Ed. 2015, 26, 143–161, doi:10.1080/09205063.2014.975393.
- Zhang, L.; Xiao, Y.; Jiang, B.; Fan, H.; Zhang, X. Effect of adipic dihydrazide modification on the performance of collagen/hyaluronic acid scaffold. J. Biomed. Mater. Res. B Appl. Biomater. 2010, 92, 307–316, doi:10.1002/jbm.b.31516.
- Guo, Y.; Yuan, T.; Xiao, Z.; Tang, P.; Xiao, Y.; Fan, Y.; Zhang, X. Hydrogels of collagen/chondroitin sulfate/hyaluronan interpenetrating polymer network for cartilage tissue engineering. J. Mater. Sci. Mater. Med. 2012, 23, 2267–2279, doi:10.1007/s10856-012-4684-5.
- Liang, W.H.; Kienitz, B.L.; Penick, K.J.; Welter, J.F.; Zawodzinski, T.A.; Baskaran, H. Concentrated collagen-chondroitin sulfate scaffolds for tissue engineering applications. J. Biomed. Mater. Res. A 2010, 94, 1050–1060, doi:10.1002/jbm.a.32774.
- Zhang, L.; Li, K.F.; Xiao, W.Q.; Zheng, L.; Xiao, Y.M.; Fan, H.S.; Zhang, X.D. Preparation of collagen-chondroitin sulfate-hyaluronic acid hybrid hydrogel scaffolds and cell compatibility in vitro. Carbohydr. Polym. 2011, 84, 118–125, doi:10.1016/j.carbpol.2010.11.009.
- Pieper, J.S.; Oosterhof, A.; Dijkstra, P.J.; Veerkamp, J.H.; van Kuppevelt, T.H. Preparation and characterization of porous crosslinked collagenous matrices containing bioavailable chondroitin sulphate. Biomaterials 1999, 20, 847–858, doi:10.1016/s0142-9612(98)00240-3.
- Pieper, J.S.; van Wachem, P.B.; van Luyn, M.J.A.; Brouwer, L.A.; Hafmans, T.; Veerkamp, J.H.; van Kuppevelt, T.H. Attachment of glycosaminoglycans to collagenous matrices modulates the tissue response in rats. Biomaterials 2000, 21, 1689–1699, doi:Doi 10.1016/S0142-9612(00)00052-1.
- Pietrucha, K. Physicochemical properties of 3D collagen-CS scaffolds for potential use in neural tissue engineering. Int. J. Biol. Macromol. 2015, 80, 732–739, doi:10.1016/j.ijbiomac.2015.07.005.
- Pietrucha, K.; Szymański, J.; Drobnik, J. The Behavior of Embryonic Neural Cells within the 3D Micro-structured Collagen-Based Scaffolds. In Proceedings of the 6th European Conference of the International Federation for Medical and Biological Engineering, Dubrovnik, Croatia, 7–11 September 2014; pp. 549–552.
- Stuart, K.; Panitch, A. Influence of chondroitin sulfate on collagen gel structure and mechanical properties at physiologically relevant levels. Biopolymers 2008, 89, 841–851, doi:10.1002/bip.21024.
- Stuart, K.; Panitch, A. Characterization of gels composed of blends of collagen I, collagen III, and chondroitin sulfate. Biomacromolecules 2009, 10, 25–31, doi:10.1021/bm800888u.
- Riley, J.F.; Shepherd, D.M.; West, G.B.; Stroud, S.W. Function of heparin. Nature 1955, 176, 1123, doi:10.1038/1761123a0.
- Copes, F.; Chevallier, P.; Loy, C.; Pezzoli, D.; Boccafoschi, F.; Mantovani, D. Heparin-Modified Collagen Gels for Controlled Release of Pleiotrophin: Potential for Vascular Applications. Front. Bioeng. Biotechnol. 2019, 7, 74, doi:10.3389/fbioe.2019.00074.
- Guidry, C.; Grinnell, F. Heparin modulates the organization of hydrated collagen gels and inhibits gel contraction by fibroblasts. J. Cell Biol. 1987, 104, 1097–1103, doi:10.1083/jcb.104.4.1097.
- Salchert, K.; Streller, U.; Pompe, T.; Herold, N.; Grimmer, M.; Werner, C. In vitro reconstitution of fibrillar collagen type I assemblies at reactive polymer surfaces. Biomacromolecules 2004, 5, 1340–1350, doi:10.1021/bm0499031.
- Watarai, A.; Schirmer, L.; Thones, S.; Freudenberg, U.; Werner, C.; Simon, J.C.; Anderegg, U. TGFbeta functionalized starPEG-heparin hydrogels modulate human dermal fibroblast growth and differentiation. Acta Biomater. 2015, 25, 65–75, doi:10.1016/j.actbio.2015.07.036.
- Fahimipour, F.; Dashtimoghadam, E.; Mahdi Hasani-Sadrabadi, M.; Vargas, J.; Vashaee, D.; Lobner, D.C.; Jafarzadeh Kashi, T.S.; Ghasemzadeh, B.; Tayebi, L. Enhancing cell seeding and osteogenesis of MSCs on 3D printed scaffolds through injectable BMP2 immobilized ECM-Mimetic gel. Dent. Mater. 2019, 35, 990–1006, doi:10.1016/j.dental.2019.04.004.
- Binner, M.; Bray, L.J.; Friedrichs, J.; Freudenberg, U.; Tsurkan, M.V.; Werner, C. Cell-instructive starPEG-heparin-collagen composite matrices. Acta Biomater. 2017, 53, 70–80, doi:10.1016/j.actbio.2017.01.086.
- Smidsrod, O.; Skjakbrk, G. Alginate as immobilization matrix for cells. Trends Biotechnol. 1990, 8, 71–78, doi:10.1016/0167-7799(90)90139-o.
- Liu, C.; Lewin Mejia, D.; Chiang, B.; Luker, K.E.; Luker, G.D. Hybrid collagen alginate hydrogel as a platform for 3D tumor spheroid invasion. Acta Biomater. 2018, 75, 213–225, doi:10.1016/j.actbio.2018.06.003.
- Moxon, S.R.; Corbett, N.J.; Fisher, K.; Potjewyd, G.; Domingos, M.; Hooper, N.M. Blended alginate/collagen hydrogels promote neurogenesis and neuronal maturation. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109904, doi:10.1016/j.msec.2019.109904.
- Hahn, M.S.; Teply, B.A.; Stevens, M.M.; Zeitels, S.M.; Langer, R. Collagen composite hydrogels for vocal fold lamina propria restoration. Biomaterials 2006, 27, 1104–1109, doi:10.1016/j.biomaterials.2005.07.022.
- Jin, G.Z.; Kim, H.W. Efficacy of collagen and alginate hydrogels for the prevention of rat chondrocyte dedifferentiation. J. Tissue Eng. 2018, 9, 2041731418802438, doi:10.1177/2041731418802438.
- Mahapatra, C.; Jin, G.Z.; Kim, H.W. Alginate-hyaluronic acid-collagen composite hydrogel favorable for the culture of chondrocytes and their phenotype maintenance. Tissue Eng. Regen. Med. 2016, 13, 538–546, doi:10.1007/s13770-016-0059-1.
- Yang, X.; Lu, Z.; Wu, H.; Li, W.; Zheng, L.; Zhao, J. Collagen-alginate as bioink for three-dimensional (3D) cell printing based cartilage tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 83, 195–201, doi:10.1016/j.msec.2017.09.002.
- Mei, E.C.; Li, S.K.; Song, J.W.; Xing, R.R.; Li, Z.M.; Yan, X.H. Self-assembling Collagen/Alginate hybrid hydrogels for combinatorial photothermal and immuno tumor therapy. Colloids Surfaces A Physicochem. Eng. Asp. 2019, 577, 570–575, doi:10.1016/j.colsurfa.2019.06.023.
- Wong, F.S.; Wong, C.C.; Chan, B.P.; Lo, A.C. Sustained Delivery of Bioactive GDNF from Collagen and Alginate-Based Cell-Encapsulating Gel Promoted Photoreceptor Survival in an Inherited Retinal Degeneration Model. PLoS ONE 2016, 11, e0159342.


