 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | rotem leshem | + 3209 word(s) | 3209 | 2020-10-21 06:04:54 | | | |
| 2 | Rita Xu | -854 word(s) | 2355 | 2020-10-22 10:42:22 | | |
Video Upload Options
Human functions and traits are linked to cerebral networks serving different emotional and cognitive control systems, some of which rely on hemispheric specialization and integration to promote adaptive goal-directed behavior. Among the neural systems discussed in this context are those underlying pro- and antisocial behaviors. The diverse functions and traits governing our social behavior have been associated with lateralized neural activity. However, as with other complex behaviors, specific hemispheric roles are difficult to elucidate. This is due largely to environmental and contextual influences, which interact with neural substrates in the development and expression of pro and antisocial functions. This paper will discuss the reciprocal ties between environmental factors and hemispheric functioning in the context of antisocial behavior. The paper will attempt to familiarize readers with the prominent literature and primary questions to encourage further research and in-depth discussion in this field.
1. Introduction
The search for neurobiological correlates of adverse social behaviors, such as violence, addiction, and crime, encompasses an ongoing investigation into the role of functional cerebral asymmetry and interhemispheric transfer [1][2][3]. Adult antisocial behaviors are defined as disruptive behaviors that violate social rules and involve defiance of authority and disregard for the rights of others [4]. They often include rule-breaking, criminal, and violent acts, as well as a failure to follow moral guidelines [5], and have been associated with the pursuit of power and the manipulation and exploitation of others to achieve personal goals [1]. While antisocial behavior is a component in psychiatric disorders, including antisocial personality disorder (APD) and psychopathy, it can also occur on its own [4].
The diverse functions and traits governing both pro and antisocial behavior, from inhibitory control to attention to empathy, have been associated with lateralized neural activity [1][6]. However, as with other complex behaviors, specific hemispheric roles are difficult to elucidate [7][8][9]. This is presumably due in large part to environmental and contextual influences, which interact with neural substrates in the development and expression of pro and antisocial functions [10][11].
2. Cerebral Asymmetry
The idea that the two hemispheres operate in different, albeit integrated, ways is the basis for discussing neural lateralization and asymmetry with respect to any particular behavior or ability [12][13]. Lateralization is believed to enable efficient use of brain tissue and increase neural capacity by preventing unnecessary duplication, contributing significantly to the brain’s plasticity and adaptability in the face of social, environmental, and pathological changes [14][15][16].
At the most basic level, the left hemisphere (LH) is generally associated with language functions including speech, comprehension, and writing, as well as arithmetic. Meanwhile, the right hemisphere (RH) is said to underlie creativity, visuo-spatial skills, and facial recognition [12][14][17]. Beyond this relatively simplistic division, several researchers have attempted to characterize hemispheric and inter-hemispheric activity in greater detail [8, 18]. The two hemispheres are believed to interpret the same data and organize contextual connections between elements of information in different ways [18][19][20]. In this view of hemispheric lateralization, the LH is associated with sequential information processing and logical thinking, and it facilitates analysis by organizing symbolic or iconic material to create a strictly ordered and unambiguously understood context [14][21]. The RH, meanwhile, is associated with image thinking and the single-stage parallel processing of many elements of information, creating an integral but ambiguous context [21].
While this and other conceptualizations of cerebral asymmetry highlight different modes of experiencing and responding to the world, it is clear that the two hemispheres are interdependent [22][23][24]. Under normal circumstances, the brain works as an integrated whole, with the two hemispheres transferring information between them via the corpus callosum to create a unified and coherent experience [1][14]. When the balance between them is disrupted by damage to a particular hemisphere or to the quality of interhemispheric transfer, specific deficits in cognitive and emotional processes can arise [2][17][25][26]. In the following section, I present research on lateralized dysfunction in relation to antisocial behaviors, including antisocial personality disorder and psychopathic traits. Later in the paper, I discuss modulation of these brain–behavior relationships by environmental and contextual factors.
3. Neural and Cognitive Substrates of Antisocial Behavior
Successful, adaptive social and interpersonal relationships generally involve solidarity, compassion, and consideration for the feelings and needs of another, all of which rely heavily on emotional comprehension and processing abilities [1][27]. Adaptive social behavior also relies on cognitive control and executive functions, including goal-directed behavior, decision-making, and planning [28][29], which support self-regulation and the inhibition of inappropriate behavior [29][30]. When any of the emotional and cognitive functions underlying adaptive social functioning are damaged, antisocial behavior can occur [31][32][33].
In accordance with the reliance of social functioning on both emotional and cognitive components, antisocial behavior has been associated with damage to brain regions involved in decision-making, inhibitory control, impulsivity, reactive aggression, and dysregulation of emotions [34][35][36]. These areas converge into two main neural networks (see Figure 1): the cognitive control network, including the lateral prefrontal and parietal cortices and parts of the anterior cingulate cortex (ACC) associated with the cognitive control processes, and the socio-emotional network, which includes the amygdala, ventral striatum, ventromedial prefrontal cortex (VMPFC), orbitofrontal cortex (OFC), and superior temporal sulcus [37][38]. Each of these networks can further be divided into left and right hemisphere components [39] which are discussed in the context of antisocial behavior below.
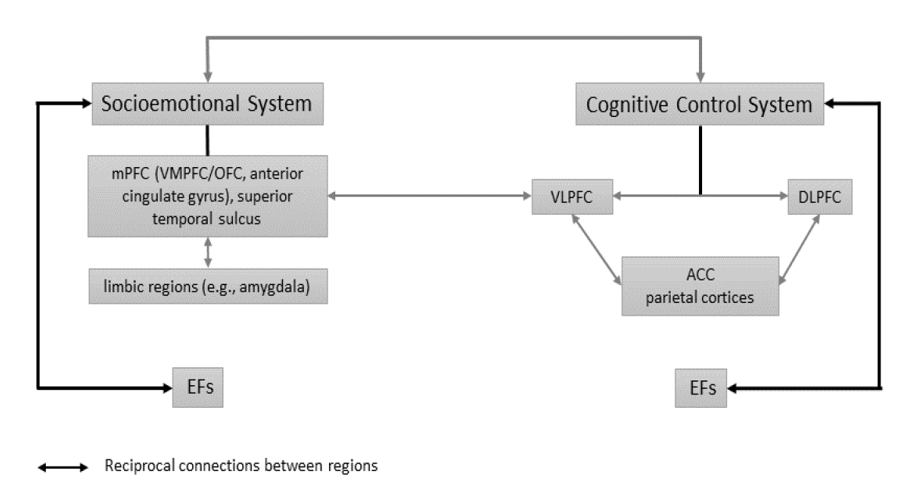
Figure 1. (Adapted from 40). Representation of the socioemotional and cognitive control systems, which are associated with executive mechanisms related to antisocial behavior. mPFC = medical prefrontal cortex; OFC = orbitofrontal cortex; VLPFC = ventrolateral prefrontal cortex; DLPFC = dorsolateral prefrontal cortex; ACC = anterior cingulate cortex; EFs = executive functions.
4. Lateralized Neural and Cognitive Substrates in Antisocial Behavior
There is evidence that anatomical and functional abnormalities in regions including the OFC, dorsolateral prefrontal cortex (DLPFC), ACC, and temporal cortex, as well as subcortical regions result in different antisocial styles when the two hemispheres are differentially affected [11][26][35][36][40]. Generally speaking, reduced RH activity appears to be associated with impaired socioemotional and behavioral functioning, such as the inability to withdraw from aversive and dangerous situations, whereas disinhibition and impaired approach behaviors, such as impulsivity, stimulation seeking, and aggression, have been linked primarily to an overactive LH [41][42][43][44]. In accordance, there are studies describing antisocial behavior characterized by affective and interpersonal deficits, believed to be mediated by an underactive RH, and antisocial behavior characterized by an impulsive, aggressive, and reward-seeking style, associated with an overactive LH [1][5][6][45]. Other studies report different, and sometimes contradicting patterns associated with LH versus RH dysfunction [14][36][46]. The large variation in the findings on lateralized neural dysfunction likely reflects heterogeneity among the samples of antisocial individuals with different patterns of cognitive and emotional deficits, as well as diverse behavioral and neuroimaging methodologies [47]. Beyond this, it is clear that neural substrates interact with the environment, which also plays a key role in the development and expression of antisocial tendencies.
5. Environmental Influences on Neural Substrates and Behavioral Expressions of Antisocial Tendencies
The roles of external environmental factors on antisocial behavior, within the context of cerebral lateralization, are discussed below with respect to two broadly-defined categories: (1) environmental effects on early and ongoing neural development, contributing to the creation of neural function and dysfunction associated with antisocial behavior, and (2) contextual determinants of antisocial behavior, or aspects of the environment that contribute to the expression of antisocial behaviors.
5.1. Environmental Effects on Early and Ongoing Development of Lateralized Neural Functions Associated
with Antisocial Behavior
While structural and functional hemispheric asymmetries become established in the human fetus, attesting to genetic underpinnings [39][48], postnatal environments also have lifelong effects on brain health [49]. A significant body of work shows that mental states and behavioral outcomes, including antisocial traits and behavior, are a reflection of both environmental experiences and innate factors that impact the brain’s ability to adapt to changing environmental demands [50][51][52][53][54]. Brain asymmetries and lateralized functions are also determined by internal and external factors before and in the years immediately following birth [49][55]. Certain areas of the RH develop earlier than the homologous areas of the LH, beginning in utero and continuing at least through the first few years of human life [56][57][58]. Brain development appears to begin with a growth spurt of the RH during the first two years of life that is shaped by emotional communications with the caregiver [59]. The RH, which is heavily involved in emotional and social processing, is believed to maintain dominance for the first three years after birth until LH development catches up [59]. Atypical brain asymmetries are believed to be the product of disruptions in these early neurodevelopmental processes [60][61]. The resulting alterations in neural circuits have been associated with behavioral phenotypes, including antisocial tendencies, as expressed in delinquency and physical aggression [60][62][63][64][65]. The most studied environmental factors among diverse antisocial populations are parenting variables (e.g., neglect, inconsistent parenting), negative life experiences (e.g., sexual abuse, physical abuse, exposure to violence), antisocial culture (e.g., poor neighborhoods), family wealth or economic status, and personal employment [50][63][66][67][68]. Beyond direct influences on RH and LH development, exposure to stress, neglect, and maltreatment during childhood can result in poor interhemispheric communication due to corpus callosal abnormalities [69][70]. A deprivation or lack of early experience reduces brain stimulation during development, which weakens the growth and development of the corpus callosum and thereby disrupts hemispheric specialization [71][72]. Reduced communication between the cortical and subcortical regions in the two hemispheres can potentially play a causal role in the affective and cognitive deficits that characterize individuals with antisocial behavior [2][26] (see Figure 2).
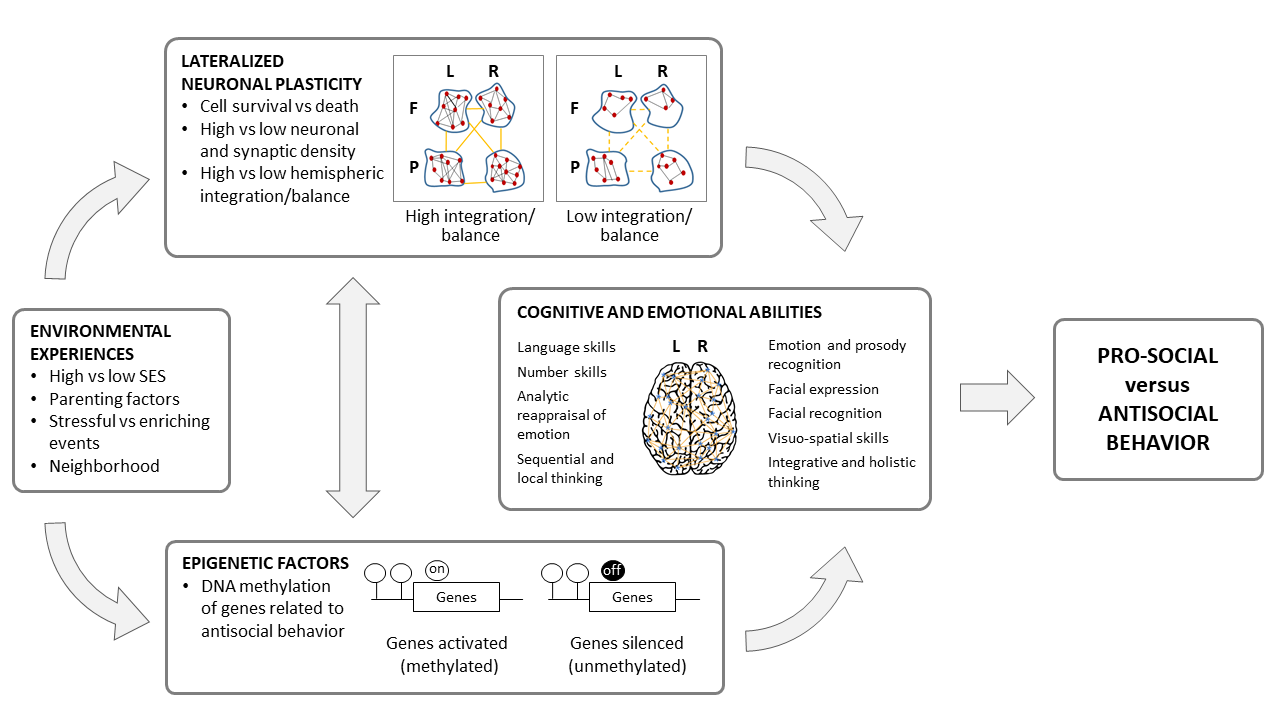
Figure 2. The interplay between environmental, epigenetic, and neural influences on the cognitive and emotional abilities underlying pro- and anti-social behavior. Life experiences and diverse environments affect connections between higher-order association areas in the frontal and parietal lobes in the left (L) and right (R) hemispheres, as well as DNA methylation-dependent gene expression. These interrelated mechanisms, in turn, affect lateralized emotional and cognitive abilities related to antisocial behavior.
The Role of Epigenetic Factors in the Development of Neural Asymmetry and Antisocial Behavior
Due to their role in the development of hemispheric asymmetries, epigenetic mechanisms, which are tied to the environment for extensive review see [39], can also a ect antisocial behavior. A primary epigenetic mechanism is DNA methylation (DNAm), a covalent modification of the DNA molecule that serves as an interface between the dynamic environment and the fixed genome [73][74]. Unlike the DNA sequence, which is stable and robust, epigenetic processes are often tissue specific, developmentally regulated, and highly dynamic [74]. The epigenetic control of biological pathways within the neuroendocrine, serotonergic, and oxytocinergic systems plays a crucial role in the cognitive and emotional processes underlying social behavior [75][76][77][78][79]. DNAm is also linked to the lateralization of nervous system organization, which is among the mechanisms through which it can contribute to antisocial behavior and related psychopathic traits.
Indeed, epigenetic mechanisms might underlie relationships between environmental factors and antisocial tendencies (see Figure 2) [10][80]. Criminology research examining early life adversity, from socioeconomic conditions to family environment to trauma, shows that life experiences can result in stable changes in DNAm [52][53][81][82]. Tobacco exposure, substance use, and psychosocial stress are among the few established environmental sources of methylation change related to antisocial behavior [83][84]. Given these recent findings, further research is needed to better understand the ontogenesis of functional lateralization at the population and individual levels, particularly with respect to antisocial behavior.
5.2. Environmental and Contextual Determinants of Antisocial Behavior
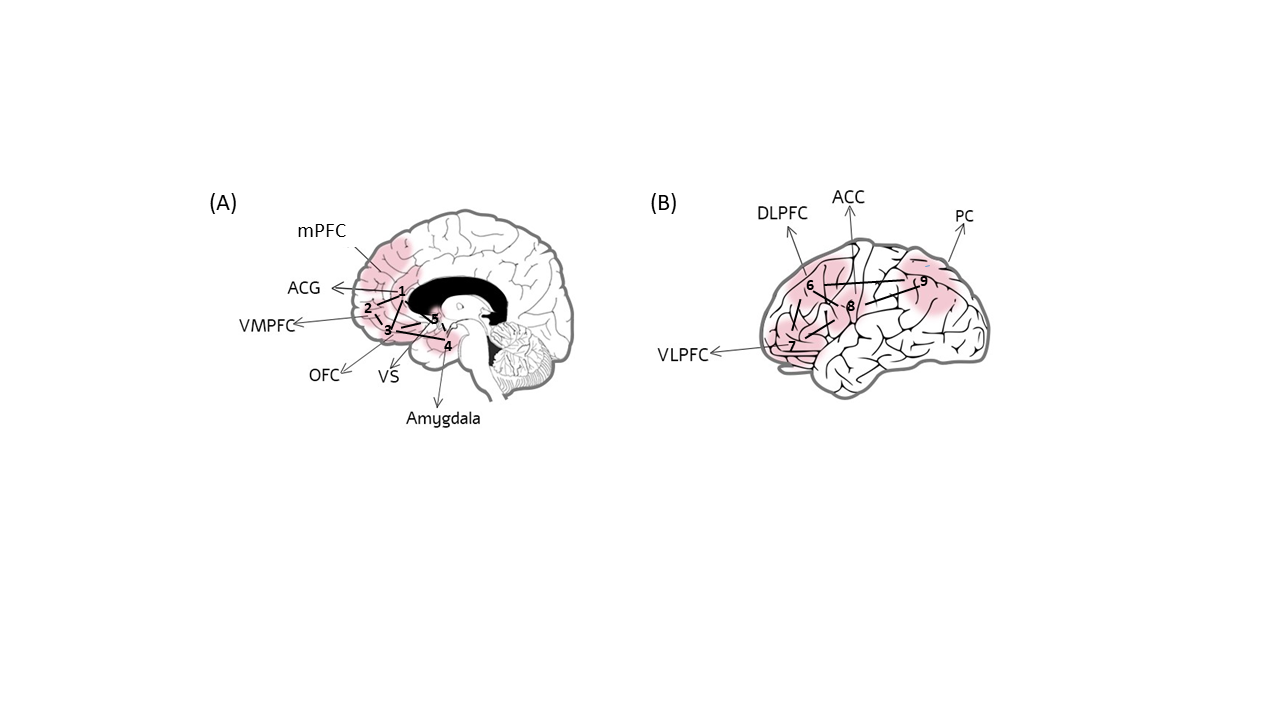
Environmental factors not only influence the development of lateralized dysfunctions underlying antisocial tendencies, but also influence the expression of these tendencies [85][86]. While the neural factors described above appear to predispose individuals to antisocial behavior, the deficits manifested in a given situation also depend on situational demands and stimulus types, which can differentially activate different regions in one or both hemispheres (see Figure 3) [87][88][89].
Figure 3. Brain areas associated with performance of tasks with predominantly emotional (A) versus cognitive (B) stimulus types. Stimulus types as well as situational demands (e.g., high/low cognitive load) can activate different regions in the socioemotional and cognitive control networks in one or both hemispheres. Depending on these and other contextual factors, deficits in these neural circuits can lead to the expression of different antisocial behaviors.
In healthy individuals, different stimulus characteristics are known to be associated with different neural patterns during cognitive and emotional task performance [90][91][92]. Nejati et al. [28], for example, investigated how the left DLPFC (l-DLPFC) and right OFC (r-OFC) interact in healthy participants during affective and cognitive executive functioning, using several behavioral tasks and transcranial direct current stimulation (tDCS). They found that the affective components of executive functioning, which they defined as risk-taking behavior and risky decision-making, required involvement of both the r-OFC and the l-DLPFC, while the cognitive components of inhibitory control and planning relied more exclusively on l-DLPFC activity. Their findings reflect the differential recruitment of the l-DLPFC and r-OFC based on the degree of involvement of cognitive control and emotional–motivational processes in a given task, and they support the possibility that impairments in these regions result in different behavioral outcomes, based on the task at hand.
Accordingly, individuals with deficits in the l-DLPFC might exhibit antisocial behavior in situations involving sudden changes in environmental constraints or internal goals, which require the suppression of irrelevant information and resolution of conflict between competing demands [5][93][94]. In contrast, individuals with deficits in the r-OFC might exhibit antisocial behavior in situations that require resisting temptation or rewards associated with risky and negative outcomes [5][95][96].
Thus, abnormal functioning in these regions will not necessarily result in antisocial behavior, but rather create antisocial tendencies that manifest differently depending on external stimuli and demands. A model that directly addresses dynamic cerebral asymmetry in antisocial behavior is the LH Activation (LHA) hypothesis [97], which largely attributes psychopathy to LH dysfunction. Specifically, the LHA hypothesis predicts that information processing is disrupted among individuals with psychopathic tendencies when LH resources are substantially and differentially activated by processing demands [98]. Cognitive deficits in individuals with psychopathic traits are therefore viewed as state-specific, or evident only under conditions that require the substantial involvement of LH resources [98][99]. This model can explain inconsistent findings among individuals with psychopathic traits in performing tasks that pose different cognitive demands [98][99]. It is also in line with the behavioral variability often exhibited by this population, which can be controlled and restrained at times, and at other times exhibits poor judgment, outbursts of rage, and poor moral judgment [98]. The LHA hypothesis is supported by imaging and behavioral studies showing LH involvement in cognitive strategy use, such as the reappraisal and regulation of emotion [25][100][101].
References
- Hecht, D. Cerebral lateralization of pro-and anti-social tendencies. Exp. Neurobiol. 2014, 23, 1–27.
- Raine, A.; Lencz, T.; Taylor, K.; Hellige, J.B.; Bihrle, S.; Lacasse, L.; Lee, M.; Ishikawa, S.; Colletti, P. Corpus callosum abnormalities in psychopathic antisocial individuals. Arch. Gen. Psychiatry 2003, 60, 1134–1142.
- Ward, T.; Wilshire, C.; Jackson, L. The contribution of neuroscience to forensic explanation. Psychol. Crime Law 2018, 24, 195–209.
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Publishing: Philadelphia, PA, USA, 2013.
- Raine, A.; Yang, Y. Neural foundations to moral reasoning and antisocial behavior. Soc. Cogn. Affect. Neurosci. 2006, 1, 203–213.
- Hecht, D. An inter-hemispheric imbalance in the psychopath’s brain. Personal. Individ. Differ. 2011, 51, 3–10.
- Chen, C.; Omiya, Y. Brain asymmetry in cortical thickness is correlated with cognitive function. Front. Hum. Neurosci. 2014, 8, 877.
- Karolis, V.R.; Corbetta, M.; De Schotten, M.T. The architecture of functional lateralisation and its relationship to callosal connectivity in the human brain. Nat. Commun. 2019, 10, 1–9.
- Kong, X.Z.; Mathias, S.R.; Guadalupe, T.; Glahn, D.C.; Franke, B.; Crivello, F.; Tzourio-Mazoyer, N.; Fisher, S.E.; Thompson, P.M.; Francks, C.; et al. Mapping cortical brain asymmetry in 17,141 healthy individuals
- Frazier, A.; Ferreira, P.A.; Gonzales, J.E. Born this way? A review of neurobiological and environmental evidence for the etiology of psychopathy. Personal. Neurosci. 2019, 2, e8.
- Reddy, K.J.; Menon, K.R.; Hunjan, U.G. Neurobiological Aspects of Violent and Criminal Behaviour: Deficits in Frontal Lobe Function and Neurotransmitters Int. J. Crim. Justice Sci. 2018, 13, 44.
- Corballis, M.C.; Häberling, I.S. The many sides of hemispheric asymmetry: A selective review and outlook. J. Int. Neuropsychol. Soc. 2017, 23, 710.
- Gotts, S.J.; Jo, H.J.; Wallace, G.L.; Saad, Z.S.; Cox, R.W.; Martin, A. Two distinct forms of functional lateralization in the human brain. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3444.
- Gonzalez, C.L.; van Rootselaar, N.A.; Gibb, R.L. Sensorimotor Lateralization Scaffolds Cognitive Specialization. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 238, pp. 405–433.
- Herbet, G.; Duffau, H. Revisiting the functional anatomy of the human brain: Toward a meta-networking theory of cerebral functions. Physiol. Rev. 2020, 100, 1181–1228.
- Vallortigara, G.; Rogers, L. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–633.
- Gazzaniga, M.S. Cerebral specialization and interhemispheric communication: Does the corpus callosum enable the human condition? Brain 2000, 123, 1293–1326.
- O’Regan, L.; Serrien, D.J. Individual differences and hemispheric asymmetries for language and spatial attention. Front. Hum. Neurosci. 2018, 12, 380.
- Badzakova-Trajkov, G.; Häberling, I.S.; Roberts, R.P.; Corballis, M.C. Cerebral asymmetries: Complementary and independent processes. PLoS ONE 2010, 5, e9682.
- Hervé, P.Y.; Zago, L.; Petit, L.; Mazoyer, B.; Tzourio-Mazoyer, N. Revisiting human hemispheric specialization with neuroimaging. Trends Cogn. Sci. 2013, 17, 69–80.
- Rotenberg, V.S.; Weinberg, I. Human memory, cerebral hemispheres, and the limbic system: A new approach. Genet. Soc. Gen. Psychol. Monogr. 1999, 125, 45–70.
- Corballis, M.C. Humanity and the left hemisphere: The story of half a brain. Laterality 2020, 1–15, doi:10.1080/1357650X.2020.1782929.
- Forrester, G.S.; Todd, B.K. A comparative perspective on lateral biases and social behavior. In Progress in Brain Research; Forrester, G.S., Hopkins, W.D., Hudry, K., Lindell, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 238, pp. 377–403.
- Ocklenburg, S.; Gunturkun, O. Hemispheric asymmetries: The comparative view. Front. Psychol. 2012, 3, 5.
- Ochsner, K.N.; Silvers, J.A.; Buhle, J.T. Functional imaging studies of emotion regulation: A synthetic review and evolving model of the cognitive control of emotion. Ann. N. Y. Acad. Sci. 2012, 1251, e1.
- Savopoulos, P.; Lindell, A.K. Born criminal? Differences in structural, functional and behavioural lateralization between criminals and noncriminals. Laterality Asymmetries Body Brain Cogn. 2018, 23, 738–760.
- Kotz, S.A.; Paulmann, S. Emotion, Language, and the Brain: Emotional Speech and Language Comprehension. Lang. Linguist. Compass 2011, 5, 108–125.
- Nejati, V.; Salehinejad, M.A.; Nitsche, M.A. Interaction of the left dorsolateral prefrontal cortex (l-DLPFC) and right orbitofrontal cortex (OFC) in hot and cold executive functions: Evidence from transcranial direct current stimulation (tDCS). Neuroscience 2018, 369, 109–123.
- Nigg, J.T. Annual Research Review: On the relations among self‐regulation, self‐control, executive functioning, effortful control, cognitive control, impulsivity, risk‐taking, and inhibition for developmental psychopathology. J. Child Psychol. Psychiatry 2017, 58, 361–383.
- Barkley, R.A. The executive functions and self-regulation: An evolutionary neuropsychological perspective. Neuropsychol. Rev. 2001, 11, 1–29.
- Brazil, I.A.; van Dongen, J.D.; Maes, J.H.; Mars, R.B.; Baskin-Sommers, A.R. Classification and treatment of antisocial individuals: From behavior to biocognition. Neurosci. Biobehav. Rev. 2018, 91, 259–277.
- Hofhansel, L.; Weidler, C.; Votinov, M.; Clemens, B.; Raine, A.; Habel, U. Morphology of the criminal brain: Gray matter reductions are linked to antisocial behavior in offenders. Brain Struct. Funct. 2020, 1–12, doi:10.1007/s00429-020-02106-6.
- Séguin, J.R. Neurocognitive elements of antisocial behavior: Relevance of an orbitofrontal cortex account. Brain Cogn. 2004, 55, 185–197.
- Bertsch, K.; Grothe, M.; Prehn, K.; Vohs, K.; Berger, C.; Hauenstein, K.; Keiper, P.; Domes, J.; Teipel, S.; Herpertz, S.C. Brain volumes differ between diagnostic groups of violent criminal offenders. Eur. Arch. Psychiatry Clin. Neurosci. 2013, 263, 593–606.
- Scott, E.S.; Steinberg, L. Adolescent development and the regulation of youth crime. Future Child. 2008, 18, 15–33.
- Van der Gronde, T.; Kempes, M.; van El, C.; Rinne, T.; Pieters, T. Neurobiological correlates in forensic assessment: A systematic review. PLoS ONE 2014, 9, e110672.
- Shulman, E.P.; Smith, A.R.; Silva, K.; Icenogle, G.; Duell, N.; Chein, J.; Steinberg, L. The dual systems model: Review, reappraisal, and reaffirmation. Dev. Cogn. Neurosci. 2016, 17, 103–117.
- Steinberg, L. A social neuroscience perspective on adolescent risk-taking. Dev. Rev. 2008, 28, 78–106.
- Schmitz, J.; Güntürkün, O.; Ocklenburg, S. Building an asymmetrical brain: The molecular perspective. Front. Psychol. 2019, 10, 982.
- Pridmore, S.; Chambers, A.; McArthur, M. Neuroimaging in psychopathy. Aust. N. Z. J. Psychiatry 2005, 39, 856–865.
- Alves, N.T.; Fukusima, S.S.; Aznar-Casanova, J.A. Models of brain asymmetry in emotional processing. Psychol. Neurosci. 2008, 1, 63.
- Coan, J.A.; Allen, J.J. Frontal EEG asymmetry and the behavioral activation and inhibition systems. Psychophysiology 2003, 40, 106–114.
- Fetterman, A.K.; Ode, S.; Robinson, M.D. For which side the bell tolls: The laterality of approach-avoidance associative networks. Motiv. Emot. 2013, 37, 33–38.
- Li, C.S.R.; Chao, H.H.A.; Lee, T.W. Neural correlates of speeded as compared with delayed responses in a stop signal task: An indirect analog of risk taking and association with an anxiety trait. Cereb. Cortex 2009, 19, 839–848.
- Raine, A.; Park, S.; Lencz, T.; Bihrle, S.; LaCasse, L.; Widom, C.S.; Al‐Dayeh, L.; Singh, M. Reduced right hemisphere activation in severely abused violent offenders during a working memory task: An fMRI study. Aggress. Behav. Off. J. Int. Soc. Res. Aggress. 2001, 27, 111–129.
- Dutton, D.G. The neurobiology of abandonment homicide. Aggress. Violent Behav. 2002, 7, 407–421.
- Yang, Y.; Raine, A. Prefrontal structural and functional brain imaging findings in antisocial, violent, and psychopathic individuals: A meta-analysis. Psychiatry Res. Neuroimaging 2009, 174, 81–88.
- Kasprian, G.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The prenatal origin of hemispheric asymmetry: An in utero neuroimaging study. Cereb. Cortex 2011, 21, 1076–1083.
- Vasung, L.; Turk, E.A.; Ferradal, S.L.; Sutin, J.; Stout, J.N.; Ahtam, B.; Lin, P.; Grant, P.E. Exploring early human brain development with structural and physiological neuroimaging. Neuroimage 2019, 187, 226–254.
- Glenn, A.L.; Raine, A. Neurocriminology: Implications for the punishment, prediction and prevention of criminal behaviour. Nat. Rev. Neurosci. 2014, 15, 54–63.
- Tremblay, R.E. Developmental origins of disruptive behaviour problems: The ‘original sin’ hypothesis, epigenetics and their consequences for prevention. J. Child Psychol. Psychiatry 2010, 51, 341–367.
- Tremblay, R.E. Developmental origins of chronic physical aggression: An international perspective on using singletons, twins and epigenetics. Eur. J. Criminol. 2015, 12, 551–561.
- Tremblay, R.E.; Vitaro, F.; Côté; S. Developmental origins of chronic physical aggression: A bio-psycho-social model for the next generation of preventive interventions. Annu. Rev. Psychol. 2018, 69, 383–407.
- Tuvblad, C.; Beaver, K.M. Genetic and environmental influences on antisocial behavior. J. Crim. Justice 2013, 41, 273–276.
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol. Rev. 2010, 20, 327–348.
- Vallesi, A. Organisation of executive functions: Hemispheric asymmetries. J. Cogn. Psychol. 2012, 24, 367–386.
- Chiron, C.; Jambaque, I.; Nabbout, R.; Lounes, R.; Syrota, A.; Dulac, O. The right brain hemisphere is dominant in human infants. Brain J. Neurol. 1997, 120, 1057–1065.
- Kinsbourne M. Development of Cerebral Lateralization in Children. In Handbook of Clinical Child Neuropsychology; Reynolds C.R., Fletcher-Janzen E., Eds.; Springer: New York, NY, USA, 2009; pp. 47–66.
- Schore, A.N. Attachment, affect regulation, and the developing right brain: Linking developmental neuroscience to pediatrics. Pediatrics Rev. 2005, 26, 204–217.
- Raine, A.; Ishikawa, S.S.; Arce, E.; Lencz, T.; Knuth, K.H.; Bihrle, S.; LaCasse, L.; Colletti, P. Hippocampal structural asymmetry in unsuccessful psychopaths. Biol. Psychiatry 2004, 55, 185–191.
- Gao, Y.; Glenn, A.L.; Schug, R.A.; Yang, Y.; Raine, A. The Neurobiology of psychopathy: A neurodevelopmental perspective. Can. J. Psychiatry 2009, 54, 813–823.
- Blair, R.J.R.; Peschardt, K.S.; Budhani, S.; Mitchell DG, V.; Pine, D.S. The development of psychopathy. J. Child Psychol. Psychiatry 2006, 47, 262–276.
- Leshem, R.; Weisburd, D. Epigenetics and hot spots of crime: Rethinking the relationship between genetics and criminal behavior. J. Contemp. Crim. Justice 2019, 35, 186–204.
- McAdams, T.A.; Gregory, A.M.; Eley, T.C. Genes of experience: Explaining the heritability of putative environmental variables through their association with behavioural and emotional traits. Behav. Genet. 2013, 43, 314–328.
- Seidel, K.; Poeggel, G.; Holetschka, R.; Helmeke, C.; Braun, K. Paternal deprivation affects the development of corticotrophin‐releasing factor‐expressing neurones in prefrontal cortex, amygdala and hippocampus of the biparental Octodon degus. J. Neuroendocrinol. 2011, 23, 1166–1176.
- Baker, L.A.; Bezdjian, S.; Raine, A. Behavioral genetics: The science of antisocial behavior. Law Contemp. Probl. 2006, 69, 7.
- Blair, R.J.R. The neurobiology of psychopathic traits in youths. Nat. Rev. Neurosci. 2013, 14, 786–799.
- Carlisi, C.O.; Moffitt, T.E.; Knodt, A.R.; Harrington, H.; Ireland, D.; Melzer, T.R.; Poulton, R.; Ramrakha, S.; Caspi, A.; Viding, E. Associations between life-course-persistent antisocial behaviour and brain structure in a population-representative longitudinal birth cohort. Lancet Psychiatry 2020, 7, 245–253.
- Teicher, M.H.; Andersen, S.L.; Polcari, A.; Anderson, C.M.; Navalta, C.P.; Kim, D.M. The neurobiological consequences of early stress and childhood maltreatment. Neurosci. Biobehav. Rev. 2003, 27, 33–44.
- Paul, R.; Henry, L.; Grieve, S.M.; Guilmette, T.J.; Niaura, R.; Bryant, R.; Bruce, S.; Williams, L.M.; Richard, C.C.; Cohen, R.A.; et al. The relationship between early life stress and microstructural integrity of the corpus callosum in a non-clinical population. Neuropsychiatr. Dis. Treat. 2008, 4, 193.
- De Bellis, M.D.; Zisk, A. The biological effects of childhood trauma. Child Adolesc. Psychiatr. Clin. 2014, 23, Jackson, D.B.; Beaver, K.M. The influence of nutritional factors on verbal deficits and psychopathic personality traits: Evidence of the moderating role of the MAOA genotype. Int. J. Environ. Res. Public Health 2015, 12, 15739–15755.
- Teicher, M.H.; Tomoda, A.; Andersen, S.L. Neurobiological consequences of early stress and childhood maltreatment: Are results from human and animal studies comparable? Ann. N. Y. Acad. Sci. 2006, 1071, 313–323.
- Guillemin, C.; Provençal, N.; Suderman, M.; Côté, S.M.; Vitaro, F.; Hallett, M.; Tremblay, R.E.; Szyf, M. DNA methylation signature of childhood chronic physical aggression in T cells of both men and women. PLoS ONE 2014, 9, e86822.
- Szyf, M. DNA methylation, the early-life social environment and behavioral disorders. J. Neurodev. Disord. 2011, 3, 238–249.
- Caspi, A.; McClay, J.; Moffitt, T.E.; Mill, J.; Martin, J.; Craig, I.W.; Poulton, R. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, 851–854.
- Jackson, D.B.; Beaver, K.M. The influence of nutritional factors on verbal deficits and psychopathic personality traits: Evidence of the moderating role of the MAOA genotype. Int. J. Environ. Res. Public Health 2015, 12, 15739–15755.
- Kraaijenvanger, E.J.; He, Y.; Spencer, H.; Smith, A.K.; Bos, P.A.; Boks, M.P. Epigenetic variability in the human oxytocin receptor (OXTR) gene: A possible pathway from early life experiences to psychopathologies. Neurosci. Biobehav. Rev. 2019, 96, 127–142.
- Palumbo, S.; Mariotti, V.; Iofrida, C.; Pellegrini, S. Genes and aggressive behavior: Epigenetic mechanisms underlying individual susceptibility to aversive environments. Front. Behav. Neurosci. 2018, 12, 117.
- Waltes, R.; Chiocchetti, A.G.; Freitag, C.M. The neurobiological basis of human aggression: A review on genetic and epigenetic mechanisms. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2016, 171, 650–675.
- Svrakic, D.M.; Cloninger, R.C. Epigenetic perspective on behaviour development, personality, and personality disorders. Psychiatr. Danub. 2010, 22, 153–166.
- Cecil, C.A.; Walton, E.; Jaffee, S.R.; O’Connor, T.; Maughan, B.; Relton, C.L.; Smith, R.G.; McArdle, W.; Gaunt, T.R.; Ouellet-Morin, I.; et al. Neonatal DNA methylation and early-onset conduct problems: A genome-wide, prospective study. Dev. Psychopathol. 2018, 30, 383–397.
- Reuben, A.; Sugden, K.; Arseneault, L.; Corcoran, D.L.; Danese, A.; Fisher, H.L.; Moffitt, T.E.; Newbury, J.B.; Odgers, C.; Rasmussen, L.J. Association of Neighborhood Disadvantage in Childhood With DNA Methylation in Young Adulthood. JAMA Netw. Open 2020, 3, e206095.
- Hannon, E.; Knox, O.; Sugden, K.; Burrage, J.; Wong, C.C.; Belsky, D.W.; Corcoran, D.L.; Arseneault, L.; Moffitt, T.E.; Caspi, A.; et al. Characterizing genetic and environmental influences on variable DNA methylation using monozygotic and dizygotic twins. PLoS Genet. 2018, 14, e1007544.
- Moffitt, T.E.; Beckley, A. Abandon Twin Research-Embrace Epigenetic Research: Premature Advice for Criminologists. Criminology 2015, 53, 121.
- Baskin-Sommers, A.R. Dissecting antisocial behavior: The impact of neural, genetic, and environmental factors. Clin. Psychol. Sci. 2016, 4, 500–510.
- Burt, S.A.; McGue, M.; Iacono, W.G. Environmental contributions to the stability of antisocial behavior over time: Are they shared or non-shared? J. Abnorm. Child Psychol. 2010, 38, 327–337.
- Shobe, E.R. Independent and collaborative contributions of the cerebral hemispheres to emotional processing. Front. Hum. Neurosci. 2014, 8, 230.
- Davis, S.W.; Cabeza, R. Cross-hemispheric collaboration and segregation associated with task difficulty as revealed by structural and functional connectivity. J. Neurosci. 2015, 35, 8191–8200.
- Stephan, K.E.; Fink, G.R.; Marshall, J.C. Mechanisms of hemispheric specialization: Insights from analyses of connectivity. Neuropsychologia 2007, 45, 209–228.
- Dolcos, F.; Katsumi, Y.; Moore, M.; Berggren, N.; de Gelder, B.; Derakshan, N.; Hamm, A.O.; Koster, E.H.; Ladouceur, C.D.; Okon-Singer, H.; et al. Neural correlates of emotion-attention interactions: From perception, learning, and memory to social cognition, individual differences, and training interventions. Neurosci. Biobehav. Rev. 2020, 108, 559–601.
- Song, S.; Zilverstand, A.; Song, H.; Uquillas, F.D.O.; Wang, Y.; Xie, C.; Cheng, L.; Zou, Z. The influence of emotional interference on cognitive control: A meta-analysis of neuroimaging studies using the emotional Stroop task. Sci. Rep. 2017, 7, 1–9.
- Weissman, D.H.; Banich, M.T. The cerebral hemispheres cooperate to perform complex but not simple tasks. Neuropsychology 2000, 14, 41.
- Nee, D.E.; Wager, T.D.; Jonides, J. Interference resolution: Insights from a meta-analysis of neuroimaging tasks. Cogn. Affect. Behav. Neurosci. 2007, 7, 1–17.
- Steinbeis, N.; Bernhardt, B.C.; Singer, T. Impulse control and underlying functions of the left DLPFC mediate age-related and age-independent individual differences in strategic social behavior. Neuron 2012, 73, 1040–1051.
- Rolls, E.T. The orbitofrontal cortex and emotion in health and disease, including depression. Neuropsychologia 2019, 128, 14–43.
- Wang, M.; Chen, Z.; Zhang, S.; Xu, T.; Zhang, R.; Suo, T.; Feng, T. High Self-Control Reduces Risk Preference: The Role of Connectivity between Right Orbitofrontal Cortex and Right Anterior Cingulate Cortex. Front. Neurosci. 2019, 13, 194.
- Kosson, D.S. Divided visual attention in psychopathic and nonpsychopathic offenders. Personal. Individ. Differ. 1998, 24, 373–391.
- Suchy, Y.; Kosson, D.S. State-dependent executive deficits among psychopathic offenders. J. Int. Neuropsychol. Soc. JINS 2005, 11, 311.
- Kosson, D.S.; Miller, S.K.; Byrnes, K.A.; Leveroni, C.L. Testing neuropsychological hypotheses for cognitive deficits in psychopathic criminals: A study of global-local processing. J. Int. Neuropsychol. Soc. 2007, 13, 267.
- Bernstein, A.; Newman, J.P.; Wallace, J.F.; Luh, K.E. Left-hemisphere activation and deficient response modulation in psychopaths. Psychol. Sci. 2000, 11, 414–417.
- Simmonite, M.; Harenski, C.L.; Koenigs, M.R.; Kiehl, K.A.; Kosson, D.S. Testing the left hemisphere activation hypothesis in psychopathic offenders using the Stroop task. Personal. Individ. Differ. 2018, 135, 182–187.


