 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Magdalena Jarosz | + 8415 word(s) | 8415 | 2020-09-30 10:32:38 |
Video Upload Options
Contrary to the conserved Elongator composition in yeast, animals, and plants, molecular functions and catalytic activities of the complex remain controversial. Elongator was identified as a component of elongating RNA polymerase II holoenzyme in yeast, animals, and plants. Furthermore, it was suggested that Elonagtor facilitates elongation of transcription via histone acetyl transferase activity. Accordingly, phenotypes of Arabidopsis elo mutants, which show development, growth, or immune response defects, correlate with transcriptional downregulation and the decreased histone acetylation in the coding regions of crucial genes. Plant Elongator was also implicated in other processes: transcription and processing of miRNA, regulation of DNA replication by histone acetylation, and acetylation of alpha-tubulin. Moreover, tRNA modification, discovered first in yeast and confirmed in plants, was claimed as the main activity of Elongator, leading to specificity in translation that might also result indirectly in a deficiency in transcription. Heterologous overexpression of individual Arabidopsis Elongator subunits and their respective phenotypes suggest that single Elongator subunits might also have another function next to being a part of the complex.
1. Definition
Elongator is a protein complex originally identified in yeast (Saccharomyces cerevisiae) when all six subunits ELP1-6 of the complex were found in a biochemical purification aimed at the elongating form of RNA polymerase II (RNAPII) [1]. The newly discovered complex was thought to bind and switch RNAPII from the initiation of transcription to the elongation state. The core subcomplex is formed by ELP1, ELP2, and ELP3, whereas the accessory subcomplex consists of ELP4, ELP5, and ELP6 [1,2,3]. A molecular architecture of the fully assembled Elongator complex reveals a symmetric dimer of ELP1, ELP2, and ELP3 with one of these subcomplexes bound to the heterohexameric ELP4, ELP5, ELP6 ring [4,5,6]. ELP3 is a catalytic subunit responsible for the enzymatic activity of the Elongator complex. It contains a C-terminal lysine (K) acetyltransferase (KAT) domain and an N-terminal radical S- adenosylmethionine domain (SAM). KAT activity was shown to be responsible for the acetylation of nucleosomal histone H3 lysine 14 and histone H4 lysine 8 in yeast, and therefore the domain was originally named histone acetyl transferase (HAT) [7]. However, later, the histone acetylation by this domain was challenged [8] and KAT was also described to target substrates other than histones which are tubulin [9,10,11] and tRNAs [8]. The N-terminal domain is a Fe4S4 cluster that is able to bind and cleave SAM in the reaction involved in tRNA wobble uridine modification [12]. ELP1 and ELP2 are primarily responsible for maintaining the structural integrity of the complex, whereas the accessory subcomplex binds tRNA molecules through ELP6 which allows the Elongator complex to perform tRNA wobble uridine modification [13].
2. Introduction
Originally, facilitation of elongation during transcription was proposed to be the main role of yeast Elongator as indicated by the interaction between Elongator and the elongating form of RNAPII, and transcriptional defects in the elp mutants were observed [2,7]. Additionally, the Elongator complex is involved in exocytosis [14], telomeric gene silencing, and DNA repair [15]. Mutations in core Elongator subunits result in the resistance to the toxin zymocin [16,17], which is the RNAse enzyme targeting tRNA anticodons containing modified wobble uridine [18]. The mutant insensitivity to the toxin was convincingly explained by the Elongator activity in the tRNA wobble uridine modifications [12]; consequently, the Elongator-deficient S. cerevisiae mutants do not contain tRNAs targeted by zymocin. Due to the tRNA-related activity, the Elongator complex is involved in the fine-tuning control of protein translation. Deleterious elp3 mutants are also characterized by growth defects such as sensitivity to temperature, salt, caffeine, and DNA-damaging factors [1,2,19].
Soon after Elongator was identified in yeast, successful isolation of the complex was also performed among multicellular eukaryotes like humans [20,21], Mus musculus [22], Caenorhabditis elegans [23], and Arabidopsis thaliana [24,25]. Interestingly, orthologues of the ELP3 protein are found in archaea wherein ELP3 catalyzes the tRNA wobble uridine modification [26,27]. The Elongator complex is highly conserved amongst eukaryotes both in terms of structure and interactions between subunits [28]. Cross species complementation analyses of genes encoding individual subunits and subdomains have experimentally proven Elongator’s conservation for yeast [29], insects [30,31], worms [23], plants [32], and humans [20].
Contrary to the well-preserved structure of Elongator in eukaryotes, activities of the complex and regulated processes are diverse and kingdom-specific. Human genes encoding Elongator subunits are associated with familial dysautonomia, which is a developmental disorder of the sensory and autonomic nervous system [33]. Defects in genes coding for Elongator subunits in mice lead to meiotic deficiencies during spermatogenesis and impairment of zygotic paternal genome demethylation in oocytes [34]. A single point mutation in Elp6 results in neuron degeneration and ataxia-like behavior of wobbly mouse [35]. In Drosophila melanogaster, Elongator has been included in several processes such as larval- and neuro-development [30,31] as well as neurotransmitter release efficiency [36]. In mice and C. elegans, Elongator acetylates neuronal alpha tubulin [9,11]. In plants, Elongator is involved in growth, organ development [37,38], cell proliferation [24], cell cycle [39], immune response [40], abscisic acid (ABA), and stress responses [41,42]. Elongata (elo) mutants of A. thaliana, named for their elongated leaves, led to an identification of the Elongator subunits, and therefore, Elongator mutants in plants are designated elo/elp. The elo/elp mutants are characterized by features such as narrow leaves [24], defective root development [38], hypersensitivity to abscisic acid (ABA) [41], and defective skotomorphogenetic and photomorphogenetic development [43]. Plant growth [25,43], root development [38], and immune response [40,44] pathways are regulated during the transcription by the histone acetylation and/or DNA demethylation enzymatic activities of Elongator. Elongator mediates non-histone acetylation of alpha tubulin [10]; it is also engaged in the microRNA biogenesis [45,46]. Finally, plant Elongator takes part in controlling the translation through its activity in tRNA wobble uridine modification [32,47,48,49,50]. Highly conserved structure of Elongator within eukaryotes and its kingdom-specific roles lead to a conclusion that Elongator regulates processes in growth and development, and response to different stimuli.
Initially, genetic data supported Elongator’s role in the transcription. However, a shift in the way of understanding the Elongator’s role took place when genetic deletion of ELP3 Schizosaccharomyces pombe ortholog resulted in a reduction of the conserved modification of uridine in the wobble position in the tRNA anticodon loop. As for Saccharomyces cerevisiae, Elongator’s role in multiple tRNA modifications was also confirmed [12]. In humans, Elongator was also found to be involved in processes other than those connected with the nucleus as Elongator subunits were located primarily in cytoplasm [20,51,52,53]. Final evidence was provided by the fact that all phenotypes of Elongator-deficient yeast cells are linked to tRNA modification and not transcription, exocytosis, telomeric gene silencing, or DNA repair [19,54]. Thus, defects observed in yeast elp mutants are caused by an indirect effect of deficient tRNA wobble modification which increases the decoding efficiency of the A- and G-ending codons [19,55,56,57]. Therefore, Elongator-mediated tRNA modification fine-tunes translation of individual proteins, and the protein synthesis is affected differently depending on the amino acid content [32].
Elongator’s involvement in tRNA modification appears to be evolutionary conserved. Mutations in genes encoding Elongator subunits cause deprivation of tRNA modification in C. elegans [23] and A. thaliana [32]. Similarly, in mouse germ cells with ELP1-deficiency, wobble uridine modification was found at a lower level [34]. The elp6 mutation in wobbly mouse destabilizes the accessory subcomplex, leading to lower levels of tRNA modifications and defects in protein translation fidelity and folding [35]. Furthermore, brain tissues and fibroblasts isolated from patients with familial dysautonomia also had lower levels of tRNA modification [58,59]. Therefore, in addition to structure, Elongator-mediated tRNA modification is also conserved in eukaryotes. As shown in bacterial Dehalococcoides mccartyi DmcELP3, anticodon stem loops of tRNA molecules are bound by a cleft formed by KAT and SAM domains, and therefore allowing ELP3 to perform the carboxy-methyl (cm5) modification, and one of the steps of this reaction includes hydrolysis of acetyl-CoA catalyzed by the KAT domain [60]. In 2019, Lin and co-workers studied structural and catalytic properties of ELP3, and identified the subunit as a non-canonical acetyltransferase and showed that specific tRNAs, but neither histone or tubulin peptides nor other nucleic acids, act as an exclusive trigger for acetyl-CoA hydrolysis in bacterial, archaeal, and eukaryotic (yeast) ELP3s [8]. With this pool of recent data, it is now believed that Elongator’s primary or sole function in yeast is to modify wobble uridines of tRNA, and that transcription defects observed in Elongator mutants constitute a consequence of a deficiency in tRNA modification which was shown to regulate translation [57]. On the other hand, studies of plant Elongator provide a wide spectrum of physiological processes affected in elo/elp mutants and indicate that Elongator regulates diverse molecular mechanisms through histone and non-histone protein acetylation, DNA (de)methylation, tRNA modification, and potential unknown activities involved in microRNA biogenesis. Furthermore, histone acetylation and DNA (de)methylation are linked to changes in the expression of specific genes related to physiological and molecular processes which are affected by the loss of Elongator which strongly supports its role in the transcription regulation. Therefore, the intriguing and still unanswered question about plant Elongator is whether different biochemical activities of the complex are synchronized in regulating individual physiological processes.
3. Elongator in Nucleus
The plant Elongator complex located in the cell nucleus is considered to regulate transcription of genes encoding proteins at the elongation stage and via DNA (de)methylation as well as pri-miRNA transcription and processing. Additionally, nuclear Elongator is also involved in DNA replication and cell-cycle progression.
3.1. Plant Elongator Regulates Transcription
3.1.1. Processes, Mechanisms, and Pathways Regulated by Elongator via Control of Transcription
Elongator regulates plant development at various stages starting from germination described as severely retarded in elo3 mutants [24]. The detailed systematic study, wherein germination is defined as the time point when endosperm rupture occurs in more than 50% of seeds, showed that in darkness, elo3-6 mutant germinates 6 h later than wild type Col-0 [63]. Further analysis of skoto- and photomorphogenesis, i.e., early seedling development in darkness and light, respectively, indicated that the early post-germination growth was also regulated by the Elongator complex [43,63]. In darkness, hypocotyls of elo/elp seedlings did not elongate as rapidly as wild type Col-0, while during photomorphogenesis, elo/elp seedlings were hyposensitive to red, far-red, and blue light as evidenced by longer hypocotyls and hyponastic cotyledons. The best-known feature of elo/elp mutants depleted in Elongator subunits is narrow and elongated leaf shape. This phenotype was later explained by a decreased palisade cell number [24], indicating that Elongator stimulates cell proliferation. Another phenotypic characteristic observed in elo mutants is the shorter primary root resulting from a reduced cell number and length pointing to the role of Elongator in both cell proliferation and cell elongation [24,38,62]. The meristem zone was smaller with a decreased cell number and also the elongation zone was shorter due to a reduced cell length [38]. Elongator is also required to maintain cell identity in the root stem cell niche [38,62] and to regulate radial patterning in roots [62]. Root phenotype together with the reduced apical dominance, abnormal inflorescence phyllotaxis, and open leaf venation observed in elo/elp mutants are all signs of defective auxin signaling and distribution [25]. The crucial role of the Elongator complex during embryogenesis and seed development was evidenced by synergistic embryo lethal phenotype of double mutants combining elo1 or elo3 and hub1 mutation in gene encoding HISTONE MONOUBIQUITINATION 1–enzyme facilitating elongation of transcription [64]. The co-expression of ELO3 and HUB1 at the torpedo stage, when double mutants’ embryos arrested, was shown by in situ hybridization and confirmed that cooperation of Elongator and HUB1 is essential for gene expression during seed development.
Apart from the important role played by Elongator in plant development, the complex has also been confirmed to regulate several response processes at the transcriptional level. Disruption of Elongator results in hypersensitivity to abscisic acid, a plant hormone regulating seed maturation, plant growth, stress responses; it also plays an important role in triggering stomatal closure. Accordingly, ABA hypersensitivity in elo/elp mutants affected seedling growth and promoted stomatal closing; on the other hand, mutants were more resistant to oxidative stress, they were also more drought-tolerant and had an increased accumulation of anthocyanins [41,42]. What is particularly well studied is the function which Elongator had in various types of immune responses which belong mainly to plant host resistance. Mutations in ELO genes impaired immune mechanisms relaying on rapid transcriptional changes, and identified basal resistance and effector-triggered immunity as regulated by Elongator [40,44,65,66]. More recently, the Elongator complex has also been presented as an important regulator of the non-host resistance against bacterial pathogens [67].
Taken together, Elongator active as a regulator of transcription is involved in a wide range of developmental processes, from germination to vegetative, generative, and seed production phases; it is also essential during stress and immune responses.
3.1.2. Transcript Levels of Individual Genes and Whole Transcriptomes are Altered in elo/elp Mutants and Plants Overexpressing Elongator Subunits
The discovery of a development or response-related defects in elo/elp mutants was followed by assays aimed at the detection of individual differentially expressed genes or transcriptome analyses, which could reveal gene expression alterations underlying the observed mutant phenotypes. The cDNA-AFLP and microarray experiments—performed to compare five elo/elp mutants to wild type—showed that genes differentially expressed in the mutants clustered together and all five mutations affect the transcription similarly [24]. As Elongator positively regulates the transcription, the downregulated genes were analyzed revealing the overrepresented Gene Ontology (GO) categories of chromatin assembly, pattern specification, vascular tissue development, and response to auxin stimulus correlating with the developmental defects of elo/elp mutants related to deficient auxin signaling and distribution [25]. Delayed germination of the elo3-6 mutant was linked to 9 germination-related genes identified as downregulated in the microarray data comparing gene expression in darkness grown mutant and wild type seedlings [63]. Analysis of the same dataset allowed to identify the downregulated GOs which were assembled into the growth-controlling network consisting of four main hubs: circadian clock, regulators of skoto- and photomorphogenesis, hormone pathways, and the primary and the secondary cell wall biogenesis [43]. The network connects upstream regulators and downstream growth effectors via signal transmission pathways. Downregulation of components of this network explains the delayed hypocotyl elongation of mutant seedlings grown in darkness. On the other hand, young elo3-6 and Col-0 seedlings grown in various light qualities and surveyed by the qPCR assay revealed positive photomorphogenesis factors to be downregulated in a mutant, which is in line with disturbed photomorphogenesis [43]. In elp2 mutant, the defective root stem cell niche and quiescent center maintenance were associated with a decreased expression of transcription factors crucial for cell identity: PLT1 (PLETHORA 1), PLT2 (PLETHORA 2), SCR (SCARECROW), SHR (SHORT ROOT), WOX5 (WUSCHEL-RELATED HOMEOBOX 5), and the auxin efflux transporter PIN1 [38]. Downregulated expression of transcription factors was confirmed by means of qPCR, and additionally, by the analysis of transgenic lines transformed with constructs containing reporter fluorescent protein fused to promoter, or promoter and coding sequence of a given transcription factor. Low expression of SHR was shown by similar methods and by in situ hybridization in another elo/elp mutant—elo1, and supported by results of qPCR assay revealing decreased levels of SHR transcriptional targets including SCR [62].The increased resistance of elo/elp mutants to oxidative [42] and drought [41] stress is very likely associated with higher accumulation of anthocyanins in the mutants, resulting from much lower expression of the MYBL2 gene encoding negative regulator of anthocyanins biosynthesis [42].
Positive regulation of immune response by Elongator was concluded based on defense deficiencies in elo/elp mutants which correlated with the aberrant expression of important defense genes [40,44,65,66,67,68]. During the bacterial pathogen induced defense response in five elp1-elp5 mutants, transcript levels of the early defense genes FRK1 (FLG22-INDUCED RECEPTOR-LIKE KINASE1), WRKY29, and GST1 (GLUTATHIONE S-TRANSFERASE1) were reduced, while the expression of the late defense PR (pathogenesis-related) genes was blocked [67]. Following the induction of defense response with exogenous NAD+, expression of PR genes was either severely delayed (PR1) or almost completely inhibited (PR2 and PR5) in the mutants [68]. Inhibition of defense response to bacterial pathogen observed in elo/elp mutants involved the decreased accumulation of reactive oxygen species (ROS) and salicylic acid (SA), and was in line with the suppressed expression of ROS production pathway gene AtrbohD (Arabidopsis respiratory burst oxidase homologue) and SA biosynthesis gene ICS1 (ISOCHORISMATE SYNTHASE1) [67]. The time-course gene expression profiles-performed by means of Northern blotting, qPCR, or microarray-revealed that induction of many defense genes is not only decreased, but also postponed in elp2 or elp3 mutants and may increase with significant delay to reach the level of the wild-type or lower [40,44,65,66]. Therefore, the authors have shown that Elongator regulates the kinetics of the rapid defense gene induction during basal, and more importantly, during effector-triggered immunity which is a highly-accelerated and amplified form of the immune response in plants. The kinetics of large scale transcriptome reprogramming, analyzed by the microarray technology, was slower in elo2 mutant compared to the wild-type after being infected with both bacterial [40] and fungal [44] pathogens. The delayed expression was also confirmed for several individual and particularly important defense genes: PR1 [65], PR2, PR5, ICS1, WRKY18, WRKY33, EDS1 (ENHANCED DISEASE SUSCEPTIBILITY1), EDS5/SA INDUCTION DEFICIENT1, PAD4 (PHYTOALEXIN DEFICIENT4), NPR1 (NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1) [66], NPR1, EDS1, EDS5, PAD4, NDR1 (NON-RACE-SPECIFIC DISEASE RESISTANCE1), ALD1 (AGD2-LIKEDEFENSE RESPONSE PROTEIN1) [40], ORA59 (OCTADECANOID-RESPONSIVE ARABIDOPSIS AP2/ERF59), and PDF1.2 (PLANT DEFENSISN1.2) [44].
Contrary to numerous reports confirming the decreased gene expression in the Elongator-deficient mutants, experiments demonstrating the upregulation of the respective genes in response to overexpression of the ELP subunits, are less common but similarly convincing. Heterologous overexpression of the Arabidopsis thaliana ELP genes in strawberry Fragaria vesca L. and in tomato Solanum lycopersicum caused increased constitutive or pathogen induced defense gene expression [69,70].
3.1.3. Elongator Regulates Elongation of Transcription
The co-purification of the Elongator complex with the phosphorylated (elongating) RNAPII in yeast [1] was an early indication that Elongator regulates transcription during the elongation phase, most likely via its histone acetylation activity. Several lines of evidence prove that the transcription elongation related function of Elongator is conserved in plants, including interaction of the complex with RNAPII and transcript elongation factors [25,61,62,71], the role in RNAPII recruitment and mutant sensitivity to transcription inhibitor [62], and finally, binding to the coding regions of the regulated genes [44] and histone modifications [25,38,40,43,44,67] and/or DNA (de)methylation [38,40]. The immunocolocalization of the Arabidopsis thaliana ELP3 subunit with the elongating form of RNAPII and euchromatin associated histone marks was described already in 2010 [25]. The direct interaction between the Arabidopsis ELP4 and ELP5 subunits and the C-terminal domain (CDT) of RNAPII was detected recently in yeast two-hybrid assays and confirmed in planta by the firefly luciferase complementation imaging (LUI) in the Nicotiana benthamiana leaves [62]. Antosz and co-workers analyzed the RNAPII Transcript Elongation Complex (TEC) in Arabidopsis suspension cultured cells using reciprocal protein tagging combined with affinity purification and mass spectrometry, and identified Elongator among proteins repeatedly associated with TEC factors SPT4 (suppressor of Ty 4), TFIIS, and PAF1-C (RNAPII-associated factor 1 complex) and to a lesser extent with RNAPII [61]. The association between ELP1 and the transcript elongation factor SPT4 was confirmed by coimmunoprecipitation of the two proteins in the protein extract of young Arabidopsis seedlings and by the LUI assay in N. bentamiana cells [62]. Interaction and cooperation of Elongator and SPT4/SPT5 during transcription elongation is suggested by similar phenotypes of elo/elp mutants and the SPT4-RNAi plants showing a reduced leaf and root cell proliferation and defective venation, as well as common downregulated auxin-related genes [72]. Elongator interacts also physically and genetically with another transcription elongation factor IYO which bounds RNAPII in in vitro pull-down and in vivo reconstitution assays [71].
The importance of Elongator in RNAPII recruitment during transcription elongation was confirmed by significantly lower in elp1 versus wild-type accumulation of RNAPII on the transcription start site and, to the greater extent, on the coding region of the SHR gene [62]. In the same study, elp1 mutants were more resistant to 6-azauracil used as an inhibitor of transcription elongation, which additionally corroborates the role of Elongator during the elongation of transcription.
Elongator epigenetically facilitates the transcription elongation via the reaction catalyzed by the KAT domain of ELP3 resulting in acetylation of the histone H3 lysine-14 residues (H3K14Ac) in coding regions of a gene [7]. This activity of Elongator in plants was confirmed by a slightly decreased acetylation level of the H3K14 in elp1 and elp3 mutants [62], but mainly by the reduced histone acetylation in coding regions of individual genes. Elongator modifies histone acetylation with high selectivity (reviewed by [73]) and the number of plant genes identified as regulated by Elongator via histone acetylation is restricted to twenty (Table 1). To find genes targeted by Elongator for histone acetylation, candidates with transcript levels decreased in elo/elp mutant are usually selected based on their role in the pathway(s) affected by the elo mutation. Histone acetylation in the candidate gene is compared between the mutant and the wild-type by means of the Chromatin Immunoprecipitation followed by qPCR (ChIP-qPCR). Chromatin is precipitated with antibodies against H3K14Ac and used to isolate DNA which is quantified in qPCR with primers designed to amplify the promoter and the coding regions of the candidate gene. Reduced histone acetylation in the elo3-6 mutant was detected in the auxin repressor gene SHY2 (SHORT HYPOCOTYL2) and auxin influx carrier gene LAX2 (AUXIN TRANSPORTR-LIKE PROTEIN2)—the two first Elongator targets involved in plant growth regulation via auxin signaling and transport, and therefore corresponding to the auxin-related phenotype of elo/elp mutants [25]. The analysis of the hypocotyl elongation delay of the darkness grown elo3-6 seedlings led to the identification of genes coding for three growth regulators and the highest order transcription factors as direct targets of the Elongator KAT activity: LHY (LATE ELONGATED HYPOCOTYL), HYH (HY-5 HOMOLOG), and HFR1 (LONG HYPOCOTYL IN FAR-RED 1) [43]. Genes of important transcription factors controlling root development: PLT1, PLT2, SHR, SCR, and gene of auxin efflux transporter PIN1 were recognized as targeted by Elongator for histone acetylation in line with their decreased expression and root defects of the elp2 mutant [38]. The SHR gene was also confirmed as the Elongator target by Qi and co-workers [62]. The largest group of the known Elongator KAT activity targets belongs to the defense pathways and comprises ten genes: NPR1, PAD4, EDS1, PR2, PR5 [40], WRKY33, ORA59, PDF1.2 [44], AtrbohD, and ICS1 [67]. Five of these genes (WRKY33, ORA59, PDF1.2, AtrbohD, and ICS1) were identified as targeted by Elongator not only based on the histone acetylation levels reduced in the elp2 mutant, but also through direct association of ELP2-GFP with the chromatin of the genes detected in the ChIP-qPCR assays using anti-GFP antibodies [44,67]. Only in the case of the WRKY33 gene, the association was identified exclusively in the coding region, while for the remaining genes, the chromatin of both promoters and the coding regions associated with ELP2-GFP. Apparently, Elongator may bind to the promoters and the coding regions of the targeted genes, however, its KAT activity is switched on when it moves—possibly as the component of the transcript elongation complex—to the coding part of the gene.
Table 1. Genes controlled by Elongator-mediated epigenetic regulation.
| Epigenetic Regulation | Genes | Biological Process | Reference |
|---|---|---|---|
| Histone acetylation | IAA3 and LAX2 | Auxin signaling pathway | [25] |
| NPR1, PR2, PR5 EDS1, PAD4 | Plant defense | [40] | |
| WRKY33, ORA59, PDF1.2 | Plant defense and ethylene signaling | [44] | |
| PLT1, PLT2, SHR, SCR, PIN1 | Root development | [38,62] | |
| RBOHD and ICS1 | Plant defense | [67] | |
| LHY, HYH, HFR1 | Circadian rhythms and phytochrome signaling pathway | [43] | |
| DNA (de)methylation | NPR1 and PAD4 | Plant defense | [40] |
| CYCB1 | Cell cycle | [38] |
3.1.4. Elongator Promotes Biogenesis of miRNAs
Regulation of the transcription elongation via histone acetylation is the best-known nucleus- and transcription-related role of Elongator played in expression of the protein-coding genes. However, the Elongator complex is also involved in expression of the miRNA-coding genes and is essential for both transcription and processing of the pri-miRNA molecules [45] (Figure 1, Inset 1). The ChIP assay indicated that Elongator is required for the full association between RNAPII and transcribed MIR genes, and therefore positively regulates their transcription as confirmed by reduced accumulation of some miRNAs in elp1–elp6 mutants coinciding with the increased levels of their complementary target transcripts. Resulting thereof deregulated expression of genes controlled by miRNAs, including factors involved in leaf development and auxin responses, might contribute to the most characteristic features of elo/elp mutants (reviewed by [73]). Elongator also interacts with the DCL1 (Dicer-like 1), HYL1 (HYPONASTIC LEAVES 1), and SE (SERRATE) factors of the Dicing complex in Dicing bodies at the nucleus [45]. Moreover, Elongator is crucial for DCL1 association with chromatin, enabling the co-transcriptional processing of pri-miRNAs by DCL1. As the plant Elongator is known to interact with RNAPII, the dual role of the complex during miRNA biogenesis is proposed to rely on simultaneous interactions with RNAPII and DCL1. These interactions physically and functionally tie together the transcription and processing of pri-miRNAs.
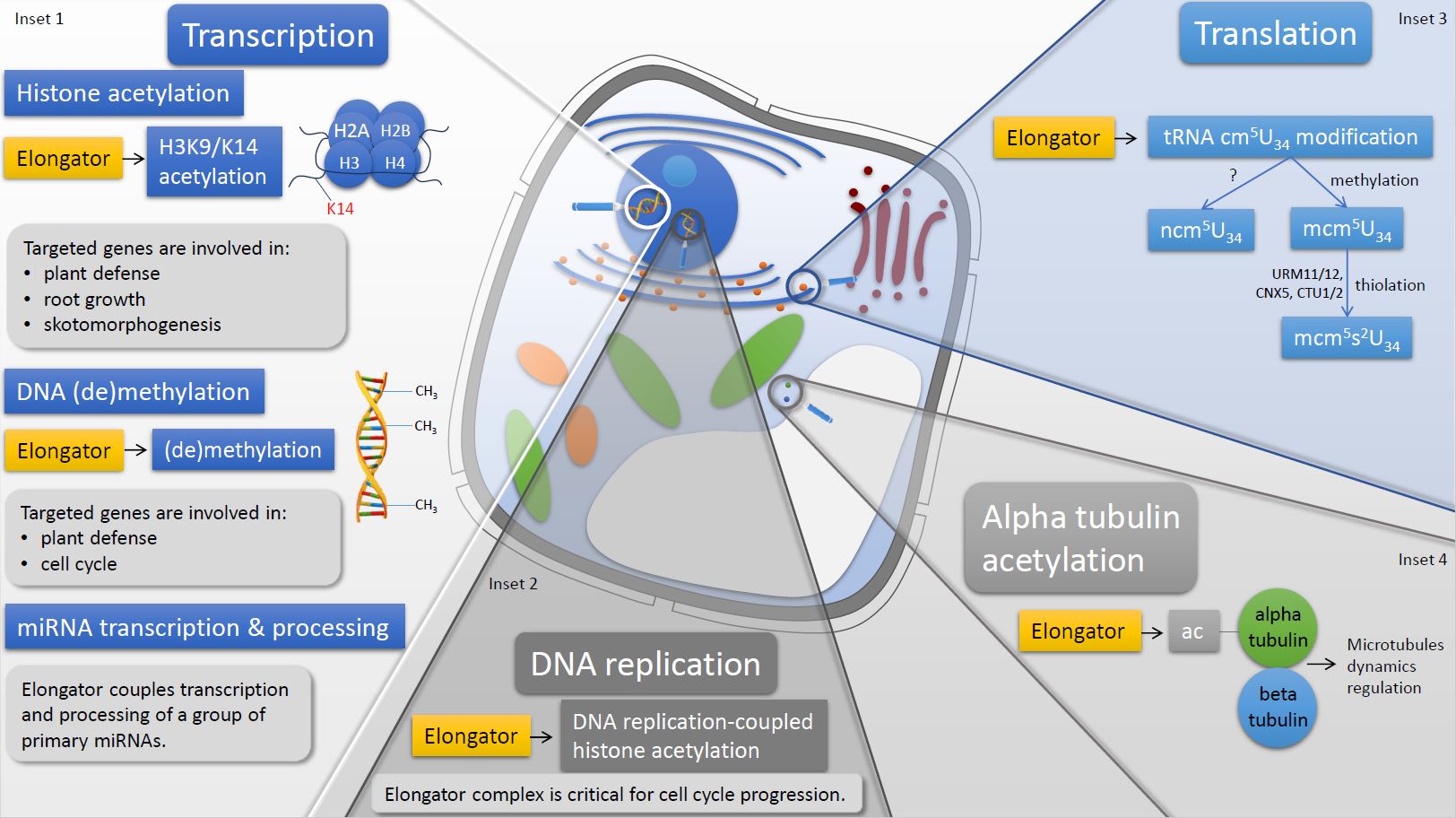
Figure 1. Schematic representation of the Elongator complex biochemical activities in a plant cell. In the nucleus, Elongator regulates the transcription through histone acetylation, DNA (de)methylation, and miRNA transcription and processing. The Elongator complex is involved in DNA replication which also occurs in the nucleus. As for cytoplasm, Elongator fine-tunes the translation by mediating tRNA wobble uridine modification and acetylates alpha tubulin.
3.1.5. Elongator Modifies DNA Methylation
The radical SAM domain of the ELP3 subunit was originally considered active in histone demethylation; however, its role in paternal DNA demethylation in mice zygotes was described later [22]. In plants, changes in DNA methylation were analyzed in elo/elp mutants of A. thaliana in the context of immune response [40] and root development [38]. Methylation levels of cytosines in promoters or the coding regions of several individual genes were compared between the elp2 mutant and wild type by means of bisulfite sequencing. Methylation of cytosines was either increased (PAD4) [40] or reduced (CYCB1, B-type CYCLIN1) [38], or conversely altered in promoter and coding region of NPR1 [40] in the elp2 mutant. The opposite changes in cytosine methylation in the elp2 mutant in different genes cannot be explained by DNA (de)methylation activity of Elongator. Genome-wide bisulfite deep sequencing also resulted in complicated patterns of cytosine methylation, showing that in the elp2 genome, the total number of methylated cytosines was higher than in wild type, however, the average methylation levels of cytosines were lower [40]. Therefore, the significant differences in cytosine methylation patterns between the elp2 mutant and wild type indicate that Elongator is somehow involved in shaping the methylation landscape of the genome (Figure 1, Inset 1), but the exact mode of action of the complex is elusive. In a tomato line with silenced ELP2 gene, the gene expression of the DRM7, DRM8, and MET1 methyltransferases was enhanced which may suggest that Elongator affects DNA methylation indirectly [74]. Moreover, SAM-dependent DNA methyl transferases differ structurally from radical SAM enzymes such as the ELP3 subunit of the Elongator complex. Hence, it seems unlikely that the Elongator complex is able to catalyze both methylation and DNA demethylation in addition to its proven radical SAM activity.
3.2. Elongator Regulates Replication and Cell-Cycle Progression
The majority of research articles concerning the nuclear functions of the plant Elongator is focused on the transcription-related activity of the complex, whereas its role in DNA replication is less recognized. In 2011, Xu and co-workers published a comprehensive study explaining how the KAT activity of Elongator is involved in DNA replication, and how aberrant replication in elo/elp mutants leads to defective cell cycle progression [39]. The replication defect was suggested by a slower increase of the nuclear ploidy in the elo3 mutant compared to wild type and reduced incorporation of EdU (5-ethynyl-2′-deoxyuridine), reflecting the rate of active DNA synthesis. In addition, the pull-down, co-immunoprecipitation, and ChIP experiments in planta indicated physical interaction of Elongator and PCNA (proliferating cells nuclear antigen), an important replication factor, and their possible common association with replicons during DNA replication. Reduced levels of the H3K56Ac and H4K5Ac histone acetylation signals in the elo3 nuclei, specifically in replicons chromatin, implied that Elongator is required for DNA replication-coupled H3 and H4 acetylation (Figure 1, Inset 2). The aberrant replication most possibly caused the inefficient DNA repair because accumulation of damaged DNA molecules was very high in the elo3 mutant and led to activation of DNA replication checkpoint to arrest the cell cycle.
The crucial role played by Elongator in DNA replication was confirmed [75], showing that mutation in the ELO3 gene leads to the failure in DNA replication and activation of cell division in the meristem following germination, which results in blocked cell cycle and arrested seedling growth. Interestingly, in situ hybridization detected dramatically reduced histone H4 mRNA levels in the shoot apex and meristem root zone of the elo3-14 mutant corresponding to aberrant DNA replication. Similarly, the transcript levels of several cell cycle-related genes, estimated using the semi-quantitative RT-PCR assay, were downregulated in line with arrested cell cycle. Therefore, it is possible that Elongator regulated progression of the cell cycle via replication-coupled histone acetylation and one of its’ transcription-related functions.
4. Elongator in Cytoplasm
Elongator complex in cytoplasm is required for post-transcriptional regulation of PIN auxin transport proteins, endoreduplication cycling, and possibly for drought stress response via tRNA wobble uridine modification. Secondly, Elongator acetylates alpha tubulin and through this process, Elongator is involved in the regulation of microtubules dynamics.
4.1. Processes and Mechanisms Regulated by Elongator via Activity in the Cytoplasm
Apart from cell nuclei, the Elongator complex is also active in the cytoplasm wherein this protein is involved in tRNA wobble uridine modification and acetylation of alpha tubulin. tRNA modification activity fine-tunes the translation process, and therefore Elongator is an important regulator of protein synthesis in plants. In the cytoplasm, Elongator also acetylates alpha tubulin and thus it is involved in regulating microtubules dynamics. Elongator-deficient plants both at the seedling phase and later developmental stages show defects in patterning, and shoot and root morphology [48,76]. It was found that these abnormalities are a consequence of the lack of the tRNA wobble uridine modification, leading to a reduced abundance of PIN auxin transport proteins. Therefore, the impaired regulation of translation leads to a defective auxin distribution. The second Elongator-dependent developmental process linked to its function in translation is leaf morphogenesis. tRNAs with wobble uridines modified by the Elongator complex are important for endoreduplication cycling, which represents a crucial process for proper formation of epidermal and mesophyll tissues [50]. Another interesting finding about Elongator’s activity in translational regulation is a strong resemblance between elo3-6 and grxs17 mutants at the morphological, molecular, and physiological level. These similarities indicate corresponding roles of Elongator and GRXS17 proteins in tRNA modification [49]. Moreover, recent studies in poplar (Populus trichocarpa) indicate that tRNA modification activity of the Elongator complex requires interaction with the protein products of the PtKTI12 genes—the yeast Kti12 ortholog required for stress response and drought stress tolerance [77].
Taken together, Elongator-mediated tRNA modification is required for fine-tuning the translation of individual proteins by increasing the decoding efficiency. Thus far, only a few processes were linked to the plant Elongator’s activity in the cytoplasm which are auxin responses and endoreduplication cycling—crucial for proper leaf morphogenesis. Most probably there are more processes controlled by the Elongator complex tRNA modification activity which are not defined yet.
4.2. Elongator Modifies Wobble U34 in tRNAs and Regulates Protein Translation
4.2.1. Conservation of Wobble U34 Modification between Yeast and Plants
In plants, Elongator’s activity during the translation was firstly described when Arabidopsis mutants lacking the ELP3 subunit showed defects in tRNA wobble uridine modification, which is at the 34th position around the anticodon stem loop [32] (Figure 1, Inset 3). tRNA modifications of bases located in this region enhance the decoding potential and accuracy of codon-anticodon pairing [78,79], which is provided by creating chemical bonds between the anticodon stem loop and its cognate and near-cognate codons [80,81]. This type of tRNA modifications primarily occurs at the 34th and 37th positions around the anticodon stem loop [79,81,82], and they are important for fine-tuning the elongation phase during protein synthesis [83,84]. Mehlgarten and co-workers were the first to show that the plant Elongator is involved in the mcm5s2U34 (5-methoxycarbonylmethyl uridine) and ncm5U34 (5-carbamoylmethyl uridine) tRNA modifications [32]. It is proposed that the Elongator complex catalyzes the attachment of carbonylmethyl to the 5th position of the tRNAs wobble uridine which results in forming 5-carbonylmethyl uridine-cm5U34 [27,32]. The wobble uridine after modification by the Elongator complex may be converted into ncm5U34 through an unknown mechanism or further methylated and thiolated to create mcm5s2U34, wherein the wobble uridine has additional sulfur atom incorporated into the 2nd position. In plants, the tRNAs transporting lysine-LysUUU, glutamic acid-GluUUC, and glutamine GlnUUG are targeted for thiolation [49,85,86].
4.2.2. The Role of the DRL1 Interactor of Elongator in tRNA Modification
In yeast, direct contact with motifs of the killer toxin-insensitive12 protein (Kti12) seems to be required for the Elongator-mediated tRNA modification [87,88]. In Arabidopsis, deformed root and leaves1 protein (DRL1) represents an orthologue of Kti12 [16,89,90]. Jun and co-workers performed functional comparative analysis of the yeast Kti12 and Arabidopsis DRL1, indicating low overall homology between these proteins; however, the presence of structurally conserved domains in different species was noted in DRL1 [91]. Complementation experiments in the yeast kti12 with Arabidopsis and rice DRL1 rescued growth retardation suggesting conserved function [91]. However, the function of DRL1 in caffeine sensitivity and zymocin-mediated growth inhibition did not overlap with Kti12 [91]. Later, the lack of functional exchange was additionally confirmed, suggesting the evolutionary diversification of Elongator and its regulatory proteins [87]. DRL1 is physically associated with the Elongator complex [92] and the yeast Kti12 motifs required for cofactors binding were shown to be conserved in the plant DRL1 [87], suggesting that similarly to Kti12, DRL1 is involved in the Elongator activity in translational regulation. In Arabidopsis, similar morphological phenotypes of drl1 and elo/elp mutants together with overlapping transcriptome changes seem to confirm corresponding functions of DRL1 and the Elongator complex [24,39,90]. Furthermore, DRL1 is involved in processes that are also regulated by the Elongator complex, such as cell division and differentiation, establishment of adaxial-abaxial polarity [91], and contribution to defense responses; however, Elongator has a broader role in plant immunity [92]. Moreover, DRL1 is required for the ncm5U34 modification which represents one of the tRNA wobble uridine modifications [47]. The newest findings in poplar—a woody plant phylogenetically close to Arabidopsis—showed that expression of PtKTI12A and PtKTI12B was induced by drought and heat stresses, whereas downregulation of these genes increased the drought tolerance [77]. Additionally, reduced levels of mcm5s2U34, mcm5s2U34, ncm5U34 modifications were identified by means of liquid chromatography–mass spectrometry LC-MS [92]. Moreover, similarly to the Elongator complex, PtKTI12A and PtKTI12B proteins are located both in the nucleus and the cytoplasm. Therefore, Wang and co-workers speculate that in woody plants, Kti12 homologs interact with the Elongator complex to participate in tRNA wobble uridine modifications [77]. Recently, Krutyholowa and co-workers presented the crystal structure of Chaetomium thermophilum N-terminal domain of Kti12 in transition state of ATP hydrolysis, and its structure strikingly resembles PSTK-an archaea O-phosphoseryl-tRNA kinase required for the synthesis of tRNA molecules transferring selenocysteine tRNASec [93]. The architecture of CtKti12 nucleotide binding pocket resembles conserved canonical P-loop ATPases. The P-loop present in Kti12 proteins is probably involved in ATP binding and hydrolysis, important for the Elongator complex to bind tRNAs [93]. Predicted structure of Populus Kti12 showed overall resemblance to CtKti12, including loop regions, suggesting that in plants, Kti12 is involved in tRNA binding and wobble uridine modification [77].
4.2.3. The role of Elongator-Mediated tRNA Modification in Auxin Responses
Further research on the Elongator’s role in the control of translation showed that its tRNA modification activity takes part in auxin distribution and responses. Mutations in the Elongator subunits ELP3 and ELP6 cause defects in the auxin-controlled development which is associated with the reduced abundance of PIN-formed (PIN) auxin transport proteins [48]. No changes in PIN transcript level were observed, suggesting that Elongator’s tRNA modification activity is involved in the post-transcriptional PINs regulation. Indeed, a reduction in mcm5s2U34 and ncm5s2U34 modifications was observed in a high-pressure liquid chromatography analysis. Moreover, expression of the endonuclease γ-toxin under RP40 promoter (RP40p::gam) caused the auxin-related defects in Arabidopsis resembling the phenotype of the Elongator-deficient plants [48]. γ-toxin preferentially degrades tRNAs modified by Elongator [18], and accordingly, the reduced abundance of tRNAGlnUUG and tRNAGluUUC was detected in RP40p::gam. Similar plant auxin-related abnormalities were observed after the downregulation of the RNA ligase RNL involved in tRNA splicing [48] which indicates the importance of the tRNA maturation in auxin signaling. Therefore, Elongator-mediated tRNA modification is involved in regulation of the PIN auxin transport protein levels which are required for proper auxin responses [48]. However, incomplete overlap of the Elongator and γ-toxin activities was observed as the elp6 RP40p::gam double mutant showed synergistic phenotype of delayed development in comparison to parental lines, suggesting for the Elongator complex and/or γ-toxin roles additional to the tRNA modification [48].
4.2.4. Similar Role of Elongator and GRXS17
The activity of Elongator in the tRNA modification was further supported by the similarity between the elo3-6 mutant and the grxs17-1 line with defective expression of the Arabidopsis cytosolic monothiol glutaredoxin GRXS17, which associated in the tandem affinity purification assays with components of the cytosolic F-S assembly pathway and proteins implicated in the tRNA metabolism. GRXS17 might be required for transferring putative Fe-S clusters to these proteins, however, the exact role of this interaction is not investigated thoroughly yet [49]. GRXS17 binds cytosolic thiouridylase subunit1 and 2 (CTU1 and CTU2), which are both essential for the sulfur atom incorporation to the tRNA wobble uridine resulting in formation of mcm5s2U34 [49]. However, GRXS17 is not required for tRNA anticodon thiolation by the CTUs as evidenced by results of PAGE retardation assay visualizing thiolated tRNAs and showing no difference between the grxs17-1 mutant and wild type Col-0. In line with the Elongator activity during the earlier stage of the same tRNA modification pathway, the elo3-6 and grxs17-1 mutants show very similar leaf and root phenotypes. Similarities were also noted at the molecular level as a large group of mostly induced genes involved in DNA-damage network overlapped between grxs17-1 and elo/elp mutants [24,39,49]. Finally, at the physiological level, it was found that likewise the Elongator complex, GRXS17 takes part in the defense response against B. cinereal [44,49]. Interestingly, despite many similarities shared by the grxs17-1 and elo3-6 mutants, grxs17 did not show a short hypocotyl phenotype of darkness-grown seedlings detected in elo/elp mutants representing the ELP1, ELP3, ELP4 subunits, and the DRL1 interactor of Elongator [43]. As shown in Figure 2, the hypocotyl lengths of the four-day-old darkness-grown seedlings of wild type Col-0 and the grxs17-1 mutant in Col-0 ecotype are not significantly different in several independent experiments.
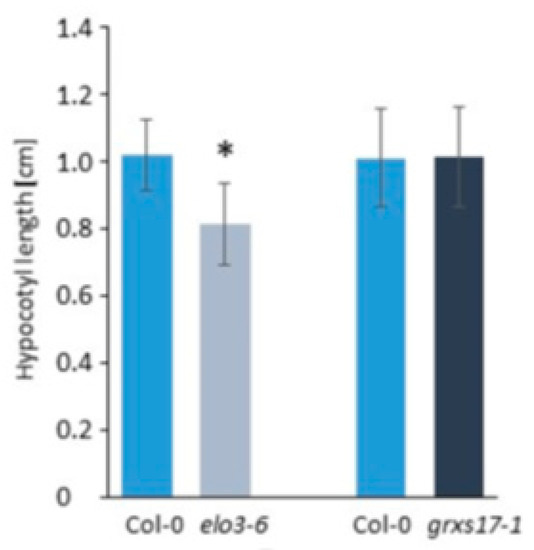
Figure 2. Hypocotyl lengths of 4-day-old elo3-6, grxs17-1, and their respective Col-0 wild-type control seedlings grown in darkness on half-strength MS medium using Image J analysis. Bars represent mean hypocotyl length of 30 seedlings (mean ± standard deviation). Differences between the mutant and wild type were statistically analyzed with an unpaired two-tailed Student’s t-test, * p < 0.01.
The short hypocotyl detected only in the elo3-6 and not in grxs17-1 mutants suggests that this phenotype is not a consequence of the compromised Elongator’s activity in tRNA modification and regulation of translation, but that it rather corresponds to another role of the complex, putatively played in the transcription regulation as indicated by transcriptome changes and reduced histone acetylation of selected genes in elo3-6 [43].
4.2.5. tRNA Wobble Uridine Modification Regulates Leaf Development
Recently, it was established that tRNAs with mcm5s2U34 modification are important for maintaining proper leaf morphogenesis [50]. Nakai and co-workers examined the Elongator-deficient plants and urm11urm12 double mutants wherein the activity of URM1-like components of the wobble uridine sulfur modification pathway is compromised. Similarly to elo/elp mutants, urm11urm12 completely lacked the sulfur modification shown in a gel retardation assays of sulfur modification of tRNAs. The phenotypes of elo/elp and urm11urm12 were also comparable as these lines had expanded the leaf area, lower chlorophyll content, and under a stereomicroscope showed disordered mesophyll formation due to the increased intercellular spaces-with elo/elp phenotype being more severe. Furthermore, observation under scanning electron microscope indicated that the elo3 and urm11urm12 plants had increased the number of epidermal cells with parallel reduction of their size, which suggests endoreduplication cycling defects. Indeed, a delay in the second endoreduplication event was found in urm11urm12, whereas in the elo3, leaf cells exhibited delayed progression already at the first endoreduplication stage measured by flow cytometry analysis [50]. The elo3 mutation enhances the leaf abaxial–adaxial polarity defect of the as2 plants with mutation in the AS2 (ASYMMETRIC LEAVES2) gene [94,95]. To explain whether the interaction between ELP3 and AS2 was caused by the Elongator function in the tRNA modification, leaf phenotypes of double elo3as2 and triple urm11urm12as2 mutants were compared. Only elo3as2 exhibited the enhanced phenotype of trumpet/needle-like leaf shape and increased expression of genes involved in maintaining abaxial leaf polarity which was not found in urm11urm12as2 [50]. These results suggest that the t-RNA modification does not affect the AS2 function, but rather another activity of Elongator—possibly the transcription-related activity—is linked to its function in abaxial–adaxial patterning of leaves.
The mcm modification mediated by Elongator is the prerequisite of the sulfur modification of wobble uridine [32,48]. Therefore, more severe phenotype of the Elongator-deficient plants than those lacking URM1-like proteins [50] might be a consequence of the absence of both the mcm and the sulfur modifications in elo mutants. The urm11urm12 double mutant lacked only the sulfur modification [50]; and whether mutations in genes encoding components of the mcm5s2U34 modification pathway have influence on the level of mcm5U34 modification in plants is not defined yet. Moreover, corresponding abnormalities of the leaf development in elo and urm11urm12 mutants indicate that the impairment of the tRNA wobble uridine modification leads to the delay of the endoreduplication cycling resulting in the unbalanced epidermal and mesophyll development [50]. Therefore, the Elongator-mediated tRNA modification together with tRNA thiolation are important for the fine-tuning the translation during leaf development.
In summary, the role of the Elongator complex in the tRNA wobble uridine modification in plants is confirmed, which allows to connect the Elongator activity with the regulation of translation. Thus far it was proven that Elongator-mediated tRNA modification translationally controls PIN proteins, and as a consequence, it regulates auxin responses which is crucial for maintaining proper plant development. Another factor which might be related to Elongator activity in tRNA wobble uridine modification is GRXS17 as its participation in the aforementioned process was proven; however, this participation is not investigated thoroughly yet. Finally, the most recently found processes wherein tRNA modification takes place are endoreduplication and leaf morphogenesis. Considering the above-given, it is interesting whether Elongator’s participation in the translation has any influence on plant processes which are not found yet. What is even more thought-provoking concerns the issue as to how translation activity is synchronized with transcriptional regulation of gene expression in plants because though in yeast the Elongator’s only activity is the regulation of translation, there is a substantial amount of evidence that supports a role for Elongator in the regulation of transcription in plants.
4.3. Elongator Acetylates alpha-Tubulin
Alpha and beta tubulins polymerase as heterodimers to form microtubules, of which functionality and dynamics are regulated by post-translational modifications including acetylation [96]. First experimental evidence showed that in mice, ELP3-as a component of the Elongator complex acetylates alpha tubulin [9]. In plants, ELP3 was purified together with PP2A phosphatase and histone deacetylase HDA14 upon microcystin-affinity chromatography, and all proteins were highly enriched in the microtubule fraction [10]. HDA14 interacts with the PP2A-A subunit and functions as a tubulin deacetylase, whereas ELP3 potentially plays an opposite role, namely it acts as an alpha tubulin acetylating enzyme [10] (Figure 1, Inset 4). Therefore, these results proved that ELP3 is able to acetylate non-histone proteins. However, the remaining subunits of the Elongator complex were not identified in this study, and therefore, it is not known whether ELP3 independently acetylates alpha tubulin or the fully assembled Elongator complex is required for maintaining this biochemical activity.
5. Do Subunits of the Plant Elongator Complex Always Act Together?
It has been well evidenced that all six ELP subunits are indispensable and have to cooperate as the integral complex to allow proper functioning of the plant Elongator. This conclusion has been repeatedly reached based on assays comparing morphological, physiological, and molecular features of plants harboring deleterious mutations in the various ELP genes and presenting very similar phenotypes. Additionally, the DRL1 interactor of Elongator has also been implicated in these studies, and in most cases, proved as necessary to assist the complex. The analyzed elo/elp mutants displayed highly related leaf phenotypes as well as gene expression profiles [24]; similar reductions in root growth [25,48,62]; common inflorescence, venation and root defects related to auxin biology [25], and comparably aberrant hypocotyl elongation in darkness or light [43]. The role of Elongator acting as the whole complex in enhancement of the leaf polarity defects associated with the mutation in AS2 (ASYMMETRIC LEAVES2) was also suggested by a similar though not equally severe contribution of different elo/elp mutants to defective adaxial–abaxial polarity observed in the double as2elo mutants [95]. Mutations in each of ELP1-ELP5 Elongator genes inhibit defense responses to bacterial pathogens [67], while all elo/elp mutants have reduced miRNAs accumulation, indicating that all Elongator subunits are required for miRNA biogenesis [45]. Finally, the wobble sulfur modification of tRNAs was equally affected in all analyzed elo/elp mutants, showing that the whole Elongator complex is involved in mcm modification, which is required for the subsequent sulfur modification [50]. Taken together, a number of strong lines of evidence linked both nuclear and cytoplasmic roles of Elongator to the complex configuration of its subunits. However, already in 2009, Zhou and co-authors discovered a substantial difference between the mutants representing various subunits of the Elongator complex [42]. Although all analyzed subunits were found similarly involved in the majority of the ABA response aspects, the stomatal closure was supersensitive to ABA only in elp1 and elp2 mutants, but not in elp4 and elp6. Considering that ELP1 and ELP2 belong to the core subcomplex, while ELP4 and ELP6 are members of the accessory subcomplex, the differences in the stomatal movement observed between the mutants may result from different functions of the subcomplexes in the epidermal guard cells. However, the most spectacular results, challenging the concept of complex composition as the ultimate condition for all Elongator activities, were provided by heterologous overexpression of individual Arabidopsis thaliana subunits [69,70]. The AtELP3 and AtELP4 genes were introduced via Agrobacterium-mediated transformation into woodland strawberry Fragaria vesca [69]. Expression of FvPR1 and FvPR5-orthologs of the A. thaliana defense genes regulated by Elongator was analyzed in lines overexpressing the AtELP3 or AtELP4 subunits. The transcript levels of both genes were significantly and markedly higher in the transgenic lines as compared to the control plants, indicating that the overexpression of subunits of the Arabidopsis Elongator leads to constitutive expression of the FvPR1 and FvPR5 genes. In line with high expression of the defense genes, the strawberry lines overexpressing AtELP3 or AtELP4 displayed enhanced resistance to fungal and bacterial pathogens. The same Elongator subunits were introduced into tomato Solanum lycopersicum, resulting in the increased—after being infected with Pseudomonas syringae—induction of the defense genes PR1b1, PR-5x, DES (DIVINYL ETHER SYNTHASE), and ER1 (ETHYLENE-RESPONSIVE PROTEASE INHIBITOR1) as detected by RNASeq and confirmed via the qPCR assay [70]. In contrast to the strawberry transgenic lines, the overexpression of the AtELP3 or AtELP4 subunits in tomato plants did not lead to constitutive expression of the defense genes but only to the increased induction of these genes following the pathogen infection. However, similarly to strawberry transgenic lines, also in tomato overexpression of AtELP3 and, particularly, AtELP4 resulted in significantly enhanced resistance to tomato bacterial speck caused by Pseudomonas syringae. Similarly stunning consequences of the overexpression of individual Elongator subunits were earlier observed in human cells, wherein the ELP3 suppressed growth of 293T embryonic kidney cells and enhanced transcription, while ELP3 and ELP4 synergistically activated transcription [97]. In addition, in yeast, ELP3 suppressed the defects in the anaphase-promoting complex 5 mutant [98]. As discussed in [69,70], effects of the individual subunits overexpression can be caused by activities of ELP3 and ELP4 independent from the Elongator complex or the A. thaliana ELP3/ELP4 may increase the activity of the Elongator complex in strawberry and tomato by an unknown mechanism. The second possibility is supported by the results of the complementation assays showing that the strawberry FvELP4-1 and the tomato SlELP3 and SlELP4 subunits are able to complement the respective Arabidopsis thaliana elp3 and elp4 mutants, indicating that the subunits of A. thaliana/strawberry or A. thaliana/tomato can interact and form functional complexes.
6. Conclusions
Studies of plant Elongator provide a large amount of data proving that, unlike in yeast, Elongator in plants combines several roles. In the nucleus, Elongator is involved in transcriptional regulation and DNA replication, and in the cytoplasm, the complex regulates translation and acetylates alpha tubulin (Figure 1 and Table 2). All activities of the plant Elongator are proved by convincing and high-quality assays including highly reliable mutant analyses which support both the translation- and transcription-related functions of the Elongator complex. The phenotypes of elo/elp mutants and grxs17 or urm11urm12 lines with defective expression of proteins involved in tRNA wobble U34 modification share many morphological, physiological, and molecular features, although show also some differences, indicating that Elongator plays an important role in tRNA modification, however, it has also other activities. On the other hand, strong resemblance between the elo/elp and SPT4-RNAi plants with affected expression of the transcription elongation factor SPT4 implicates that Elongator regulates transcription during the elongation phase. Therefore, in plants, the collected data strongly indicates the coexistence of transcriptional and translational activities of the Elongator complex and suggests their potential synchronization; however, the physiological role and mechanism of this interplay remain elusive. Contrary to the long-lasting and well-documented discussion about the dominant function of Elongator, data suggesting that individual subunits or sub-complexes may have activities independent of the entire Elongator complex are still sparse and mostly recent. Revealing the mechanism by which the heterologously overexpressed individual subunits of Elongator may effectively influence gene expression and pathogen resistance will be important to understand the exact mode of action of Elongator.
Table 2. Elongator regulates molecular mechanisms in the nucleus and the cytoplasm through different biochemical activities in Arabidopsis thaliana.
| Localization | Molecular Process | Biochemical Activity | The Role of Elongator | Reference |
|---|---|---|---|---|
| Nucleus | Transcription elongation | Histone acetylation | Elongator regulates auxin signaling. | [25] |
| Elongator regulates mitotic cell cycle and leaf patterning. |
[39] | |||
| Elongator is involved in plant defense. | [40] | |||
| The role of Elongator in B. cinerea infection. | [44] | |||
| Elongator regulates root development. | [38,62] | |||
| Elongator is involved in plant defense. | [67] | |||
| Elongator regulates skoto- and photomorphogenesis. | [43] | |||
| - | IYO interacts with RNAPII and Elongator to promote elongation and initiate cell differentiation. | [71] | ||
| Elongator and TEFs associate with elongating RNAPII. | [61] | |||
| Elongator is involved in root stem cell maintenance through unknown mechanism. | [62] | |||
| Transcription | DNA (de)methylation | Elongator is involved in plant defense. | [40] | |
| Elongator regulates root development. | [38] | |||
| miRNA biogenesis | Elongator regulates miRNA transcription and processing | [45] | ||
| [46] | ||||
| Unknown | Elongator regulates MYBL2 expression which is involved in anthocyanin biosynthesis. | [42] | ||
| Elongator mediates the establishment of leaf polarity through unknown gene expression system. | [95] | |||
| Elongator acts as meristem cell cycle activator. | [75] | |||
| Elongator controls expression of germination-related genes. | [63] | |||
| Cytoplasm | Translation | tRNA wobble uridine modification | Elongator-mediated tRNA modification is conserved in plants. Elongator is involved in mcm5s2U34 and ncm5U34 modification. | [32] |
| Elongator and DRL1 are involved in ncm5U34 modification. | [47] | |||
| Elongator regulates auxin responses by controlling PIN protein level. | [48] | |||
| Strong resemblance between elo3-6 and grxs17. GRXS17 is involved in tRNA modification. | [49] | |||
| Elongator-mediated tRNA modification regulates leaf morphogenesis. | [50] | |||
| Non-histone protein acetylation | Alpha tubulin acetylation | Elongator is involved in microtubules dynamics regulation. | [10] |
[1][2][3][4][5][6][7][8][9][10][11][12][13][14][15][16][17][18][19][20][21][22][23][24][25][26][27][28][29][30][31][32][33][34]
[35][36][37][38][39][40][41][42][43][44][45][46][47][48][49][50][51][52][53][54][55][56][57][58][59][60][61][62][63][64][65][66][67][68][69][70][71][72][73][74][75][76][77][78][79][80][81][82][83][84][85][86][87][88][89][90][91][92][93][94][95][96][97][98]
References
- Gabriel Otero; Jane Fellows; Yang Li; Therese De Bizemont; Annette M.G Dirac; Claes M. Gustafsson; Hediye Erdjument-Bromage; Paul Tempst; Jesper Q. Svejstrup; Elongator, a Multisubunit Component of a Novel RNA Polymerase II Holoenzyme for Transcriptional Elongation. Molecular Cell 1999, 3, 109-118, 10.1016/s1097-2765(00)80179-3.
- Nevan J. Krogan; Jack F. Greenblatt; Characterization of a Six-Subunit Holo-Elongator Complex Required for the Regulated Expression of a Group of Genes in Saccharomyces cerevisiae. Molecular and Cellular Biology 2001, 21, 8203-8212, 10.1128/mcb.21.23.8203-8212.2001.
- Gerlof Sebastiaan Winkler; Thodoris G. Petrakis; Steen Ethelberg; Masao Tokunaga; Hediye Erdjument-Bromage; Paul Tempst; Jesper Q. Svejstrup; RNA Polymerase II Elongator Holoenzyme Is Composed of Two Discrete Subcomplexes. Journal of Biological Chemistry 2001, 276, 32743-32749, 10.1074/jbc.m105303200.
- Sebastian Glatt; Juliette Létoquart; Céline Faux; Nicholas M.I. Taylor; Bertrand Séraphin; Christoph W Müller; Christoph W Müller; The Elongator subcomplex Elp456 is a hexameric RecA-like ATPase. Nature Structural & Molecular Biology 2012, 19, 314-320, 10.1038/nsmb.2234.
- María I. Daudén; Jan Kosinski; Olga Kolaj‐Robin; Ambroise Desfosses; Alessandro Ori; Céline Faux; Niklas Alexander Hoffmann; Osita F Onuma; Karin D. Breunig; Martin Beck; et al.Carsten SachseBertrand SéraphinSebastian GlattChristoph W Müller Architecture of the yeast Elongator complex. EMBO reports 2016, 18, 264-279, 10.15252/embr.201643353.
- Dheva T Setiaputra; Derrick Th Cheng; Shan Lu; Jesse M Hansen; Udit Dalwadi; Cindy Hy Lam; Jeffrey L To; Meng‐Qiu Dong; Calvin K. Yip; Molecular architecture of the yeast Elongator complex reveals an unexpected asymmetric subunit arrangement. EMBO reports 2016, 18, 280-291, 10.15252/embr.201642548.
- Gerlof Sebastiaan Winkler; Arnold Kristjuhan; Hediye Erdjument-Bromage; Paul Tempst; Jesper Q. Svejstrup; Elongator is a histone H3 and H4 acetyltransferase important for normal histone acetylation levelsin vivo. Proceedings of the National Academy of Sciences 2002, 99, 3517-3522, 10.1073/pnas.022042899.
- Ting-Yu Lin; Nour El Hana Abbassi; Karol Zakrzewski; Andrzej Chramiec-Głąbik; Małgorzata Jemioła-Rzemińska; Jan Różycki; Sebastian Glatt; The Elongator subunit Elp3 is a non-canonical tRNA acetyltransferase. Nature Communications 2019, 10, 1-12, 10.1038/s41467-019-08579-2.
- Catherine Creppe; Lina Malinouskaya; Marie-Laure Volvert; Magali Gillard; Pierre Close; Olivier Malaise; Sophie Laguesse; Isabelle Cornez; Souad Rahmouni; Sandra Ormenese; et al.Shibeshih BelachewBrigitte EmalgrangeJean-Paul ChapelleUlrich SiebenlistGustave MoonenAlain ChariotNguyen Laurent Elongator Controls the Migration and Differentiation of Cortical Neurons through Acetylation of α-Tubulin. Cell 2009, 136, 551-564, 10.1016/j.cell.2008.11.043.
- Hue T. Tran; Mhairi Nimick; R. Glen Uhrig; George Templeton; Nick Morrice; Robert Gourlay; Alison Delong; Greg B.G. Moorhead; Arabidopsis thaliana histone deacetylase 14 (HDA14) is an α-tubulin deacetylase that associates with PP2A and enriches in the microtubule fraction with the putative histone acetyltransferase ELP3. The Plant Journal 2012, 71, 263-272, 10.1111/j.1365-313x.2012.04984.x.
- Jachen A. Solinger; Roberta Paolinelli; Holger Klöß; Francesco Berlanda Scorza; Stefano Marchesi; Ursula Sauder; Dai Mitsushima; Fabrizio Capuani; Stephen R. Stürzenbaum; Giuseppe Cassata; et al. The Caenorhabditis elegans Elongator Complex Regulates Neuronal α-tubulin Acetylation. PLOS Genetics 2010, 6, e1000820, 10.1371/journal.pgen.1000820.
- B. Huang; Marcus J.O. Johansson; Anders S. Byström; An early step in wobble uridine tRNA modification requires the Elongator complex. RNA 2005, 11, 424-436, 10.1261/rna.7247705.
- Zhijie Lin; Weijing Zhao; Wentao Diao; Xingqiao Xie; Zheng Wang; Jinxiu Zhang; Yuequan Shen; Jiafu Long; Crystal Structure of Elongator Subcomplex Elp4–6. Journal of Biological Chemistry 2012, 287, 21501-21508, 10.1074/jbc.m112.341560.
- Peter B. Rahl; Catherine Z. Chen; Ruth N. Collins; Elp1p, the Yeast Homolog of the FD Disease Syndrome Protein, Negatively Regulates Exocytosis Independently of Transcriptional Elongation. Molecular Cell 2005, 17, 841-853, 10.1016/j.molcel.2005.02.018.
- Qing Li; A. M. Fazly; Hui Zhou; Shengbing Huang; Zhiguo Zhang; Bruce Stillman; The Elongator Complex Interacts with PCNA and Modulates Transcriptional Silencing and Sensitivity to DNA Damage Agents. PLOS Genetics 2009, 5, e1000684, 10.1371/journal.pgen.1000684.
- Frank Frohloff; Lars Fichtner; Daniel Jablonowski; Karin D. Breunig; Raffael Schaffrath; Saccharomyces cerevisiae Elongator mutations confer resistance to the Kluyveromyces lactis zymocin. The EMBO Journal 2001, 20, 1993-2003, 10.1093/emboj/20.8.1993.
- D. Jablonowski; R. Schaffrath; Zymocin, a composite chitinase and tRNase killer toxin from yeast. Biochemical Society Transactions 2007, 35, 1533-1537, 10.1042/bst0351533.
- Jian Lu; Bo Huang; Anders Esberg; Marcus J.O. Johansson; Anders S. Byström; The Kluyveromyces lactis -toxin targets tRNA anticodons. RNA 2005, 11, 1648-1654, 10.1261/rna.2172105.
- Anders Esberg; Bo Huang; Marcus J.O. Johansson; Anders S. Byström; Elevated Levels of Two tRNA Species Bypass the Requirement for Elongator Complex in Transcription and Exocytosis. Molecular Cell 2006, 24, 139-148, 10.1016/j.molcel.2006.07.031.
- Nicola A. Hawkes; Gabriel Otero; G. Sebastiaan Winkler; Nick Marshall; Michael E. Dahmus; Daniel Krappmann; Claus Scheidereit; Claire L. Thomas; Giampietro Schiavo; Hediye Erdjument-Bromage; et al.Paul TempstJesper Q. Svejstrup Purification and Characterization of the Human Elongator Complex. Journal of Biological Chemistry 2001, 277, 3047-3052, 10.1074/jbc.m110445200.
- Pierre Close; Magali Gillard; Aurélie Ladang; Zheshen Jiang; Jessica Papuga; Nicola Hawkes; Nguyen Laurent; Jean-Paul Chapelle; Fabrice Bouillenne; Jesper Q. Svejstrup; et al.Marianne FilletAlain Chariot DERP6 (ELP5) and C3ORF75 (ELP6) Regulate Tumorigenicity and Migration of Melanoma Cells as Subunits of Elongator*. Journal of Biological Chemistry 2012, 287, 32535-32545, 10.1074/jbc.M112.402727.
- Yuki Okada; Kazuo Yamagata; Kwonho Hong; Teruhiko Wakayama; Yanping Zhang; A role for the elongator complex in zygotic paternal genome demethylation. Nature Cell Biology 2010, 463, 554-558, 10.1038/nature08732.
- Changchun Chen; Simon Tuck; A. S. Byström; Defects in tRNA Modification Associated with Neurological and Developmental Dysfunctions in Caenorhabditis elegans Elongator Mutants. PLOS Genetics 2009, 5, e1000561, 10.1371/journal.pgen.1000561.
- Hilde Nelissen; Delphine Fleury; Leonardo Bruno; Pedro Robles; Lieven De Veylder; Jan Traas; José Luis Micol; Marc Van Montagu; Dirk Inzé; Mieke Van Lijsebettens; et al. The elongata mutants identify a functional Elongator complex in plants with a role in cell proliferation during organ growth. Proceedings of the National Academy of Sciences 2005, 102, 7754-7759, 10.1073/pnas.0502600102.
- Hilde Nelissen; Steven De Groeve; Delphine Fleury; Pia Neyt; Leonardo Bruno; Maria Beatrice Bitonti; Filip Vandenbussche; Dominique Van Der Straeten; Takahiro Yamaguchi; Hirokazu Tsukaya; et al.Erwin WittersGeert De JaegerAndreas HoubenMieke Van Lijsebettens Plant Elongator regulates auxin-related genes during RNA polymerase II transcription elongation. Proceedings of the National Academy of Sciences 2010, 107, 1678-1683, 10.1073/pnas.0913559107.
- Christina Paraskevopoulou; Shirley A. Fairhurst; David J. Lowe; Peter Brick; Silvia Onesti; The Elongator subunit Elp3 contains a Fe4S4 cluster and binds S-adenosylmethionine. Molecular Microbiology 2006, 59, 795-806, 10.1111/j.1365-2958.2005.04989.x.
- Kiruthika Selvadurai; Pei Wang; Joseph Seimetz; Raven H. Huang; Archaeal Elp3 catalyzes tRNA wobble uridine modification at C5 via a radical mechanism. Nature Chemical Biology 2014, 10, 810-812, 10.1038/nchembio.1610.
- Sebastian Glatt; Christoph W Müller; Christoph W Müller; Structural insights into Elongator function. Current Opinion in Structural Biology 2013, 23, 235-242, 10.1016/j.sbi.2013.02.009.
- F. Frohloff; D. Jablonowski; L. Fichtner; R. Schaffrath; Subunit Communications Crucial for the Functional Integrity of the Yeast RNA Polymerase II Elongator ( -Toxin Target (TOT)) Complex. Journal of Biological Chemistry 2002, 278, 956-961, 10.1074/jbc.m210060200.
- Neetu Singh; Meridith T. Lorbeck; Ashley Zervos; John Zimmerman; Felice Elefant; The histone acetyltransferase Elp3 plays in active role in the control of synaptic bouton expansion and sleep in Drosophila. Journal of Neurochemistry 2010, 115, 493-504, 10.1111/j.1471-4159.2010.06892.x.
- Jane Walker; So Yeon Kwon; Paul Badenhorst; Phil East; Helen McNeill; Jesper Q. Svejstrup; Role of Elongator Subunit Elp3 in Drosophila melanogaster Larval Development and Immunity. Genetics 2011, 187, 1067-1075, 10.1534/genetics.110.123893.
- Constance Mehlgarten; Daniel Jablonowski; Uta Wrackmeyer; Susan Tschitschmann; David Sondermann; Gunilla Jäger; Zhizhong Gong; Anders S. Byström; Raffael Schaffrath; Karin D. Breunig; et al. Elongator function in tRNA wobble uridine modification is conserved between yeast and plants. Molecular Microbiology 2010, 77, 531-531, 10.1111/j.1365-2958.2010.07253.x.
- Susan A. Slaugenhaupt; Anat Blumenfeld; Sandra P. Gill; Maire Leyne; James Mull; Math P. Cuajungco; Christopher B. Liebert; Brian Chadwick; Maria Idelson; Luba Reznik; et al.Christiane M. RobbinsIzabela MakalowskaMichael J. BrownsteinDaniel KrappmannClaus ScheidereitChanna MaayanFelicia B. AxelrodJames F. Gusella Tissue-Specific Expression of a Splicing Mutation in the Gene Causes Familial Dysautonomia. The American Journal of Human Genetics 2001, 68, 598-605, 10.1086/318810.
- Fu-Jung Lin; Li Shen; Chuan-Wei Jang; Pål Ø. Falnes; Yanping Zhang; Ikbkap/Elp1 Deficiency Causes Male Infertility by Disrupting Meiotic Progression. PLOS Genetics 2013, 9, e1003516, 10.1371/journal.pgen.1003516.
- Marija Kojic; Monika Gaik; Bence Kiska; Anna Salerno-Kochan; Sarah Hunt; Angelo Tedoldi; Sergey Mureev; Alun Jones; Belinda Whittle; Laura A. Genovesi; et al.Christelle AdolpheDarren L. BrownJennifer L. StowKirill AlexandrovPankaj SahSebastian GlattBrandon J. Wainwright Elongator mutation in mice induces neurodegeneration and ataxia-like behavior. Nature Communications 2018, 9, 3195, 10.1038/s41467-018-05765-6.
- Katarzyna Miskiewicz; Liya E. Jose; André Bento-Abreu; Marcus Fislage; Ines Taes; Jarosław Kasprowicz; Jef Swerts; Stephan Sigrist; Wim Versées; Wim Robberecht; et al.Patrik Verstreken ELP3 Controls Active Zone Morphology by Acetylating the ELKS Family Member Bruchpilot. Neuron 2011, 72, 776-788, 10.1016/j.neuron.2011.10.010.
- Andrea Falcone; Hilde Nelissen; Delphine Fleury; Mieke Van Lijsebettens; Maria Beatrice Bitonti; Cytological Investigations of the Arabidopsis thaliana elo1 Mutant Give New Insights into Leaf Lateral Growth and Elongator Function. Annals of Botany 2007, 100, 261-270, 10.1093/aob/mcm102.
- Yuebin Jia; Huiyu Tian; Hongjiang Li; Qianqian Yu; Lei Wang; Jiri Friml; Zhaojun Ding; TheArabidopsis thalianaelongator complex subunit 2 epigenetically affects root development. Journal of Experimental Botany 2015, 66, 4631-4642, 10.1093/jxb/erv230.
- Deyang Xu; Weihua Huang; Yang Li; Hua Wang; Hai Huang; Xiaofeng Cui; Elongator complex is critical for cell cycle progression and leaf patterning in Arabidopsis. The Plant Journal 2011, 69, 792-808, 10.1111/j.1365-313x.2011.04831.x.
- Yongsheng Wang; Chuanfu An; Xudong Zhang; Jiqiang Yao; Yanping Zhang; Yijun Sun; Fahong Yu; David Moraga Amador; Zhonglin Mou; The Arabidopsis Elongator Complex Subunit2 Epigenetically Regulates Plant Immune Responses. The Plant Cell 2013, 25, 762-776, 10.1105/tpc.113.109116.
- Zhizhong Chen; Hairong Zhang; Daniel Jablonowski; Xiaofeng Zhou; Xiaozhi Ren; Xuhui Hong; Raffael Schaffrath; Jian-Kang Zhu; Zhizhong Gong; Mutations in ABO1/ELO2, a Subunit of Holo-Elongator, Increase Abscisic Acid Sensitivity and Drought Tolerance in Arabidopsis thaliana. Molecular and Cellular Biology 2006, 26, 6902-6912, 10.1128/mcb.00433-06.
- Xiaofeng Zhou; Deping Hua; Zhizhong Chen; Zhengjun Zhou; Zhizhong Gong; Elongator mediates ABA responses, oxidative stress resistance and anthocyanin biosynthesis in Arabidopsis. The Plant Journal 2009, 60, 79-90, 10.1111/j.1365-313x.2009.03931.x.
- Magdalena Woloszynska; Olimpia Gagliardi; Filip Vandenbussche; Steven De Groeve; Luis Alonso Baez; Pia Neyt; Sabine Le Gall; Jorge Fung; Paloma Mas; Dominique Van Der Straeten; et al.Mieke Van Lijsebettens The Elongator complex regulates hypocotyl growth in darkness and during photomorphogenesis. Journal of Cell Science 2017, 131, jcs203927, 10.1242/jcs.203927.
- Chenggang Wang; Yezhang Ding; Jin Yao; Yanping Zhang; Yijun Sun; James Colee; Zhonglin Mou; Arabidopsis Elongator subunit 2 positively contributes to resistance to the necrotrophic fungal pathogensBotrytis cinereaandAlternaria brassicicola. The Plant Journal 2015, 83, 1019-1033, 10.1111/tpj.12946.
- Xiaofeng Fang; Yuwei Cui; Yaoxi Li; Yijun Qi; Transcription and processing of primary microRNAs are coupled by Elongator complex in Arabidopsis. Nature Plants 2015, 1, 15075, 10.1038/nplants.2015.75.
- Sascha Laubinger; MicroRNA transcription and processing: Elongator caught in the act. Nature Plants 2015, 1, 15076, 10.1038/nplants.2015.76.
- Peng Chen; Gunilla Jäger; Bo Zheng; Transfer RNA modifications and genes for modifying enzymes in Arabidopsis thaliana. BMC Plant Biology 2010, 10, 1-19, 10.1186/1471-2229-10-201.
- Johannes Leitner; Katarzyna Retzer; Nenad Malenica; Rasa Bartkeviciute; Doris Lucyshyn; Gunilla Jäger; Barbara Korbei; A. S. Byström; Christian Luschnig; Meta-regulation of Arabidopsis Auxin Responses Depends on tRNA Maturation. Cell Reports 2015, 11, 516-526, 10.1016/j.celrep.2015.03.054.
- Sabrina Inigo; Astrid Nagels Durand; Andrés Ritter; Sabine Le Gall; Martin Termathe; Roland Klassen; Takayuki Tohge; Barbara De Coninck; Jelle Van Leene; Rebecca De Clercq; et al.Bruno P. A. CammueAlisdair R. FernieKris GevaertGeert De JaegerSebastian A. LeidelRaffael SchaffrathMieke Van LijsebettensLaurens PauwelsAlain Goossens Glutaredoxin GRXS17 Associates with the Cytosolic Iron-Sulfur Cluster Assembly Pathway. Plant Physiology 2016, 172, 858-873, 10.1104/pp.16.00261.
- Yumi Nakai; Gorou Horiguchi; Kosei Iwabuchi; Akiko Harada; Masato Nakai; Ikuko Hara-Nishimura; Takato Yano; tRNA Wobble Modification Affects Leaf Cell Development in Arabidopsis thaliana.. Plant And Cell Physiology 2019, 60, 2026-2039, 10.1093/pcp/pcz064.
- Lars Fichtner; Frank Frohloff; Daniel Jablonowski; Michael J.R. Stark; R Schaffrath; Protein interactions within Saccharomyces cerevisiae Elongator, a complex essential for Kluyveromyces lactis zymocicity.. Molecular Microbiology 2002, 45, 817-826, 10.1046/j.1365-2958.2002.03055.x.
- Jae-Hyun Kim; William S. Lane; Danny Reinberg; Human Elongator facilitates RNA polymerase II transcription through chromatin. Proceedings of the National Academy of Sciences 2002, 99, 1241-1246, 10.1073/pnas.251672198.
- Dmitry K Pokholok; Nancy M Hannett; Richard A Young; Exchange of RNA Polymerase II Initiation and Elongation Factors during Gene Expression In Vivo. Molecular Cell 2002, 9, 799-809, 10.1016/s1097-2765(02)00502-6.
- Changchun Chen; Bo Huang; Mattias Eliasson; Patrik Ryden; A. S. Byström; Elongator Complex Influences Telomeric Gene Silencing and DNA Damage Response by Its Role in Wobble Uridine tRNA Modification. PLOS Genetics 2011, 7, e1002258, 10.1371/journal.pgen.1002258.
- Philippe C. Durant; Ashok C. Bajji; Mallikarjun Sundaram; Raju K. Kumar; Darrell R. Davis; Structural Effects of Hypermodified Nucleosides in theEscherichia coliand Human tRNALysAnticodon Loop: The Effect of Nucleosides s2U, mcm5U, mcm5s2U, mnm5s2U, t6A, and ms2t6A†. Biochemistry 2005, 44, 8078-8089, 10.1021/bi050343f.
- Ulrike Begley; Madhu Dyavaiah; Ashish Patil; John P. Rooney; Dan DiRenzo; Christine M. Young; Douglas S. Conklin; Richard S. Zitomer; Thomas J. Begley; Trm9-Catalyzed tRNA Modifications Link Translation to the DNA Damage Response. Molecular Cell 2007, 28, 860-870, 10.1016/j.molcel.2007.09.021.
- Marcus J. O. Johansson; Anders Esberg; Bo Huang; Glenn R. Björk; Anders S. Byström; Eukaryotic Wobble Uridine Modifications Promote a Functionally Redundant Decoding System. Molecular and Cellular Biology 2008, 28, 3301-3312, 10.1128/mcb.01542-07.
- Tony Karlsborn; Hasan Tükenmez; Changchun Chen; A. S. Byström; Familial dysautonomia (FD) patients have reduced levels of the modified wobble nucleoside mcm5s2U in tRNA. Biochemical and Biophysical Research Communications 2014, 454, 441-445, 10.1016/j.bbrc.2014.10.116.
- Mayumi Yoshida; Naoyuki Kataoka; Kenjyo Miyauchi; Kenji Ohe; Kei Iida; Suguru Yoshida; Takayuki Nojima; Yukiko Okuno; Hiroshi Onogi; Tomomi Usui; et al.Akihide TakeuchiTakamitsu HosoyaTsutomu SuzukiMasatoshi Hagiwara Rectifier of aberrant mRNA splicing recovers tRNA modification in familial dysautonomia. Proceedings of the National Academy of Sciences 2015, 112, 2764-2769, 10.1073/pnas.1415525112.
- Sebastian Glatt; René Zabel; Olga Kolaj-Robin; Osita F. Onuma; Florence Baudin; Andrea Graziadei; Valerio Taverniti; Ting-Yu Lin; Frauke Baymann; Bertrand Séraphin; et al.Karin D. BreunigChristoph W Müller Structural basis for tRNA modification by Elp3 from Dehalococcoides mccartyi. Nature Structural & Molecular Biology 2016, 23, 794-802, 10.1038/nsmb.3265.
- Wojciech Antosz; Alexander Pfab; Hans F. Ehrnsberger; Philipp Holzinger; Karin Köllen; Simon A. Mortensen; Astrid Bruckmann; Thomas Schubert; Gernot Längst; Joachim Griesenbeck; et al.Veit SchubertMarion GrasserKlaus D. Grasser The Composition of the Arabidopsis RNA Polymerase II Transcript Elongation Complex Reveals the Interplay between Elongation and mRNA Processing Factors.. The Plant Cell 2017, 29, 854-870, 10.1105/tpc.16.00735.
- Linlin Qi; Xiaoyue Zhang; Huawei Zhai; Jian Liu; Fangming Wu; Chuanyou Li; Qian Chen; Elongator Is Required for Root Stem Cell Maintenance by Regulating SHORTROOT Transcription. Plant Physiology 2018, 179, 220-232, 10.1104/pp.18.00534.
- Magdalena Woloszynska; Olimpia Gagliardi; Filip Vandenbussche; Mieke Van Lijsebettens; Elongator promotes germination and early post-germination growth. Plant Signaling & Behavior 2018, 13, e1422465, 10.1080/15592324.2017.1422465.
- Kristiina Himanen; Magdalena Woloszynska; Tommaso M Boccardi; Steven De Groeve; Hilde Nelissen; Leonardo Bruno; Marnik Vuylsteke; Mieke Van Lijsebettens; Histone H2B monoubiquitination is required to reach maximal transcript levels of circadian clock genes in Arabidopsis.. The Plant Journal 2012, 72, 249–260, 10.1111/j.1365-313X.2012.05071.x.
- Christopher T. DeFraia; Xudong Zhang; Zhonglin Mou; Elongator subunit 2 is an accelerator of immune responses in Arabidopsis thaliana. The Plant Journal 2010, 64, 511-523, 10.1111/j.1365-313x.2010.04345.x.
- Christopher T DeFraia; Yongsheng Wang; Jiqiang Yao; Zhonglin Mou; Elongator subunit 3 positively regulates plant immunity through its histone acetyltransferase and radical S-adenosylmethionine domains. BMC Plant Biology 2013, 13, 102-102, 10.1186/1471-2229-13-102.
- Chuanfu An; Chenggang Wang; Zhonglin Mou; The Arabidopsis Elongator complex is required for nonhost resistance against the bacterial pathogensXanthomonas citrisubsp.citriandPseudomonas syringaepv.phaseolicolaNPS3121. New Phytologist 2017, 214, 1245-1259, 10.1111/nph.14442.
- Chuanfu An; Yezhang Ding; Xudong Zhang; Chenggang Wang; Zhonglin Mou; Elongator Plays a Positive Role in Exogenous NAD-Induced Defense Responses in Arabidopsis. Molecular Plant-Microbe Interactions® 2016, 29, 396-404, 10.1094/mpmi-01-16-0005-r.
- Katchen Julliany P. Silva; Asha M. Brunings; Juliana A. Pereira; Natalia A. Peres; Kevin M. Folta; Zhonglin Mou; The Arabidopsis ELP3/ELO3 and ELP4/ELO1 genes enhance disease resistance in Fragaria vesca L.. BMC Plant Biology 2017, 17, 230, 10.1186/s12870-017-1173-5.
- Juliana A. Pereira; Fahong Yu; Yanping Zhang; Jeffrey B. Jones; Zhonglin Mou; The Arabidopsis Elongator Subunit ELP3 and ELP4 Confer Resistance to Bacterial Speck in Tomato. Frontiers in Plant Science 2018, 9, 1-14, 10.3389/fpls.2018.01066.
- Maite Sanmartín; Michael Sauer; Alfonso Munoz; Jan Zouhar; Angel Ordoñez; Wilhelmina T.G. Van De Ven; Elena Caro; María De La Paz Sánchez; Natasha V. Raikhel; Crisanto Gutierrez; et al.Jose J. Sánchez-SerranoEnrique Rojo A Molecular Switch for Initiating Cell Differentiation in Arabidopsis. Current Biology 2011, 21, 999-1008, 10.1016/j.cub.2011.04.041.
- Mieke Van Lijsebettens; Julius Durr; Magdalena Woloszynska; Klaus D. Grasser; Elongator and SPT4/SPT5 complexes as proxy to study RNA polymerase II transcript elongation control of plant development. PROTEOMICS 2014, 14, 2109-2114, 10.1002/pmic.201400024.
- Magdalena Woloszynska; Sabine Le Gall; Mieke Van Lijsebettens; Plant Elongator-mediated transcriptional control in a chromatin and epigenetic context. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2016, 1859, 1025-1033, 10.1016/j.bbagrm.2016.06.008.
- Mingku Zhu; Yali Li; Guoping Chen; Lijun Ren; Qiaoli Xie; Zhiping Zhao; Hu Zongli; Silencing SlELP2L, a tomato Elongator complex protein 2-like gene, inhibits leaf growth, accelerates leaf, sepal senescence and produces dark-green fruit. Scientific Reports 2015, 5, 7693, 10.1038/srep07693.
- Anna Skylar; Sean Matsuwaka; Xuelin Wu; ELONGATA3 is required for shoot meristem cell cycle progression in Arabidopsis thaliana seedlings. Developmental Biology 2013, 382, 436-445, 10.1016/j.ydbio.2013.08.008.
- Nenad Malenica; Lindy Abas; Rene Benjamins; Saeko Kitakura; Harald F. Sigmund; Kim S. Jun; Marie-Theres Hauser; Jiří Friml; Christian Luschnig; MODULATOR OF PIN genes control steady-state levels of Arabidopsis PIN proteins. The Plant Journal 2007, 51, 537-550, 10.1111/j.1365-313x.2007.03158.x.
- Wang, H.; Xu, C.; Zhang, Y.; Yan, X.; Jin, X.; Yao, X.; Chen, P.; Zheng, B. PtKTI12 genes influence wobbleuridine modifications and drought stress tolerance in hybrid poplar. Tree Physiol. 2020.
- Paul F. Agris; The importance of being modified: an unrealized code to RNA structure and function.. RNA 2015, 21, 552-4, 10.1261/rna.050575.115.
- Ville Y. P. Väre; Emily R. Eruysal; Amithi Narendran; Kathryn L. Sarachan; Paul F. Agris; Chemical and Conformational Diversity of Modified Nucleosides Affects tRNA Structure and Function. Biomolecules 2017, 7, 29, 10.3390/biom7010029.
- Franck A. P. Vendeix; Frank V. Murphy; William A. Cantara; Grażyna Leszczyńska; Estella M. Gustilo; Brian Sproat; Andrzej Malkiewicz; Paul F. Agris; Human tRNALys3UUU Is Pre-Structured by Natural Modifications for Cognate and Wobble Codon Binding through Keto–Enol Tautomerism. Journal of Molecular Biology 2012, 416, 467-485, 10.1016/j.jmb.2011.12.048.
- Sandra Blanchet; David Cornu; Isabelle Hatin; Henri Grosjean; Pierre Bertin; Olivier Namy; Deciphering the reading of the genetic code by near-cognate tRNA. Proceedings of the National Academy of Sciences 2018, 115, 3018-3023, 10.1073/pnas.1715578115.
- Paul F. Agris; Franck A.P. Vendeix; William D. Graham; tRNA’s Wobble Decoding of the Genome: 40 Years of Modification. Journal of Molecular Biology 2007, 366, 1-13, 10.1016/j.jmb.2006.11.046.
- Alexey Rozov; Natalia Demeshkina; Iskander Khusainov; Eric Westhof; Marat Yusupov; Gulnara Yusupova; Novel base-pairing interactions at the tRNA wobble position crucial for accurate reading of the genetic code. Nature Communications 2016, 7, 10457, 10.1038/ncomms10457.
- Namit Ranjan; Marina V. Rodnina; Thio-Modification of tRNA at the Wobble Position as Regulator of the Kinetics of Decoding and Translocation on the Ribosome. Journal of the American Chemical Society 2017, 139, 5857-5864, 10.1021/jacs.7b00727.
- Ruth-Maria Leiber; Florian John; Yves Verhertbruggen; Anouck Diet; Paul Knox; Christoph Ringli; The TOR Pathway Modulates the Structure of Cell Walls in Arabidopsis. The Plant Cell 2010, 22, 1898-1908, 10.1105/tpc.109.073007.
- Yumi Nakai; Akiko Harada; Yasuyuki Hashiguchi; Masato Nakai; Hideyuki Hayashi; Arabidopsis Molybdopterin Biosynthesis Protein Cnx5 Collaborates with the Ubiquitin-like Protein Urm11 in the Thio-modification of tRNA*. Journal of Biological Chemistry 2012, 287, 30874-30884, 10.1074/jbc.M112.350090.
- Constance Mehlgarten; Heike Prochaska; Alexander Hammermeister; Wael R. Abdel-Fattah; Melanie Wagner; Rościsław Krutyhołowa; Sang Eun Jun; Gyung-Tae Kim; Sebastian Glatt; Karin D. Breunig; et al.Michael J.R. StarkRaffael Schaffrath Use of a Yeast tRNase Killer Toxin to Diagnose Kti12 Motifs Required for tRNA Modification by Elongator. Toxins 2017, 9, 272, 10.3390/toxins9090272.
- Rościsław Krutyhołowa; Annekathrin Reinhardt; Andrzej Chramiec-Głąbik; Karin D. Breunig; Sebastian Glatt; Fungal Kti12 proteins display unusual linker regions and unique ATPase p-loops. Current Genetics 2020, 66, 823–833, 10.1007/s00294-020-01070-2.
- A R Butler; J H White; Y Folawiyo; A Edlin; D Gardiner; Michael J.R. Stark; Two Saccharomyces cerevisiae genes which control sensitivity to G1 arrest induced by Kluyveromyces lactis toxin.. Molecular and Cellular Biology 1994, 14, 6306-6316, 10.1128/mcb.14.9.6306.
- Hilde Nelissen; Jonathan H Clarke; Marc De Block; Sabine De Block; Rudy Vanderhaeghen; Raymond E Zielinski; Tristan Dyer; Sofie Lust; Dirk Inzé; Mieke Van Lijsebettens; et al. DRL1, a homolog of the yeast TOT4/KTI12 protein, has a function in meristem activity and organ growth in plants.. The Plant Cell 2003, 15, 639–654.
- Sang Eun Jun; Kiu-Hyung Cho; Ji-Young Hwang; Wael R. Abdel-Fattah; Alexander Hammermeister; Raffael Schaffrath; John L. Bowman; Gyung-Tae Kim; Comparative analysis of the conserved functions of Arabidopsis DRL1 and yeast KTI12.. Molecules and Cells 2014, 38, 243-50, 10.14348/molcells.2015.2297.
- Chenggang Wang; Xudong Zhang; Jian-Liang Li; Yanping Zhang; Zhonglin Mou; The Elongator complex-associated protein DRL1 plays a positive role in immune responses against necrotrophic fungal pathogens in Arabidopsis. Molecular Plant Pathology 2017, 19, 286-299, 10.1111/mpp.12516.
- Rościsław Krutyhołowa; Alexander Hammermeister; Rene Zabel; Wael R. Abdel-Fattah; Annekathrin Reinhardt; Mark Helm; Michael J.R. Stark; Karin D. Breunig; Raffael Schaffrath; Sebastian Glatt; et al. Kti12, a PSTK-like tRNA dependent ATPase essential for tRNA modification by Elongator.. Nucleic Acids Research 2019, 47, 4814-4830, 10.1093/nar/gkz190.
- Chiyoko Machida; Ayami Nakagawa; Shoko Kojima; Hiro Takahashi; Yasunori Machida; The complex of ASYMMETRIC LEAVES (AS) proteins plays a central role in antagonistic interactions of genes for leaf polarity specification in Arabidopsis. Wiley Interdisciplinary Reviews: Developmental Biology 2015, 4, 655-671, 10.1002/wdev.196.
- Shoko Kojima; Mayumi Iwasaki; Hiro Takahashi; Tomoya Imai; Yoko Matsumura; Delphine Fleury; Mieke Van Lijsebettens; Yasunori Machida; Chiyoko Machida; ASYMMETRIC LEAVES2 and Elongator, a Histone Acetyltransferase Complex, Mediate the Establishment of Polarity in Leaves of Arabidopsis thaliana. Plant And Cell Physiology 2011, 52, 1259-1273, 10.1093/pcp/pcr083.
- Wang, W.; Vignani, R.; Scali, M.; Sensi, E.; Cresti, M. Post-translational modifications of α-tubulin in Zea maysL. are highly tissue specific. Planta 2004, 218, 460–465.
- Junxia Gu; Ngmei Sun; Qiping Zheng; Xiaochun Wang; Huicui Yang; Jingcheng Miao; Changping Wu; Wenxiang Wei; Human Elongator complex is involved in cell cycle and suppresses cell growth in 293T human embryonic kidney cells.. Acta Biochimica et Biophysica Sinica 2009, 41, 831-838, 10.1093/abbs/gmp072.
- Emma L. Turner; Mackenzie E. Malo; Marnie G. Pisclevich; Megan D. Dash; Gerald F. Davies; Terra G. Arnason; Troy Anthony Alan Harkness; The Saccharomyces cerevisiae Anaphase-Promoting Complex Interacts with Multiple Histone-Modifying Enzymes To Regulate Cell Cycle Progression. Eukaryotic Cell 2010, 9, 1418-1431, 10.1128/ec.00097-10.


