 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Bonny Burns-Whitmore | + 3368 word(s) | 3368 | 2021-09-01 05:14:09 | | | |
| 2 | Peter Tang | Meta information modification | 3368 | 2021-09-02 03:50:08 | | |
Video Upload Options
Good sources of the long-chain n-3 fatty acids, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) include cold-water fish and seafood; however, vegan diets (VGNs) do not include animal-origin foods.
1. Introduction
2. Dietary Sources of the Essential Fatty Acids
|
Nutrient 1 |
Energy |
Protein |
Total Lipid |
18:2n-6 |
18:3n-3 |
CHO |
Total Fiber |
|
|---|---|---|---|---|---|---|---|---|
|
Unit |
Kcal |
g |
g |
g |
g |
g |
g |
|
|
Oils |
||||||||
|
Oil, Canola |
100 g |
884 |
0 |
100 |
18.64 |
9.137 |
0 |
0 |
|
Oil, Flaxseed/Linseed (Panos) |
100 g |
884 |
0.110 |
99.98 |
14.25 |
53.37 |
0 |
0 |
|
Oil, Soybean |
100 g |
763 |
0 |
100 |
50.42 |
6.789 |
0 |
0 |
|
Oil, Walnut |
100 g |
884 |
0 |
100 |
52.90 |
10.40 |
0 |
0 |
|
Oil, High Oleic Sunflower |
100 g |
884 |
0 |
100 |
3.61 * |
0.192 ** |
0 |
0 |
|
Oil, High Oleic Safflower |
100 g |
884 |
0 |
100 |
12.72 |
0.096 |
0 |
0 |
|
Oil, Culinary Algae |
100 g |
800 |
0 |
93.33 |
n/a |
n/a |
0 |
0 |
|
Ovega-3 supplement 2 |
1 gel |
n/a |
0 |
n/a |
n/a |
0.5 |
0 |
0 |
|
Deva Vegan supplement 3 |
1 cap |
5 |
0 |
0.5 |
n/a |
0.2 |
0 |
0 |
|
Nordic Naturals supplement 4 |
2 gels |
10.0 |
0 |
1.00 |
n/a |
0.715 |
0 |
0 |
|
Food Sources |
||||||||
|
Almonds Raw |
100 g |
579 |
21.15 |
49.93 |
12.30 |
0.003 |
21.55 |
12.50 |
|
Amaranth |
100 g |
371 |
13.65 |
7.020 |
2.736 |
0.042 |
65.25 |
6.70 |
|
Avocados, Raw, California |
100 g |
167 |
1.960 |
15.41 |
1.674 |
0.111 |
8.640 |
6.80 |
|
Black Walnuts Dried |
100 g |
619 |
24.06 |
59.33 |
33.80 |
2.680 |
9.580 |
6.80 |
|
Brazil Nuts, Dried |
100 g |
659 |
14.32 |
67.10 |
23.859 |
0.018 |
11.74 |
7.50 |
|
Brown Rice Cooked |
100 g |
123 |
2.740 |
0.970 |
0.355 |
0.011 |
25.58 |
1.60 |
|
Bulgur Cooked |
100 g |
83 |
3.08 |
0.240 |
0.094 |
0.004 |
18.58 |
4.50 |
|
Cashews Raw |
100 g |
553 |
18.22 |
43.85 |
7.782 |
0.062 |
30.19 |
3.30 |
|
Chia Seeds Dried |
100 g |
486 |
16.54 |
30.74 |
5.840 |
17.80 |
42.12 |
34.4 |
|
English Walnuts Dried |
100 g |
654 |
15.23 |
65.21 |
38.09 |
9.08 |
13.71 |
6.70 |
|
Flaxseed Raw |
100 g |
534 |
18.29 |
42.16 |
5.903 |
22.81 |
28.88 |
27.3 |
|
Hempseed Hulled |
100 g |
553 |
31.56 |
48.75 |
1.340 |
8.864 |
8.670 |
4.00 |
|
Millet Cooked |
100 g |
119 |
3.510 |
1.000 |
0.480 |
0.028 |
23.67 |
1.300 |
|
Oat Bran Cooked |
100 g |
40 |
3.210 |
0.860 |
0.324 |
0.015 |
11.44 |
2.60 |
|
Pistachio Raw |
100 g |
560 |
20.16 |
45.32 |
13.10 |
0.210 |
27.17 |
10.60 |
|
Poppy Seeds |
100 g |
525 |
17.99 |
41.56 |
28.30 |
0.273 |
28.13 |
19.50 |
|
Quinoa |
100 g |
368 |
14.12 |
6.070 |
2.977 |
0.260 |
64.16 |
7.00 |
|
Rye |
100 g |
338 |
10.34 |
1.630 |
0.659 |
0.108 |
75.86 |
15.1 |
|
Sesame Seeds dried |
100 g |
573 |
17.73 |
49.67 |
21.375 |
0.376 |
23.45 |
11.8 |
|
Soybeans Raw |
100 g |
446 |
36.49 |
19.94 |
9.925 |
1.330 |
30.16 |
9.30 |
|
Soybeans, Boiled |
100 g |
141 |
12.35 |
6.400 |
2.657 |
0.354 |
11.05 |
4.20 |
|
Sunflower Seeds |
100 g |
584 |
20.78 |
51.50 |
23.05 * |
0.06 ** |
20.00 |
8.60 |
3. Nutrient Bioavailability
4. Physiological Reasons for Special Consideration-Conversion of ALA to EPA and DHA
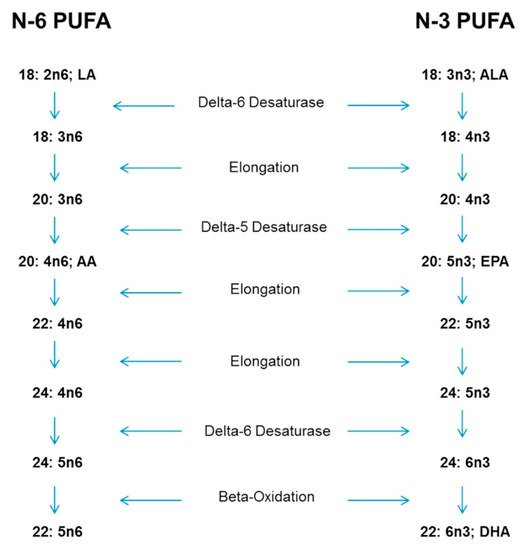
5. Recommended Intakes of n-3 and the n-6:n-3 Ratio
|
Author, Year |
Recommendation: Males; Females Combined |
18:2n-6 LA |
18:3 n-3 ALA |
20:5 n-3 EPA |
22:6 n-3 DHA |
n-6:n-3 or Omega-3 Index |
|---|---|---|---|---|---|---|
|
Agnoli et al., 2017 [36] |
Females |
Limit intake of sources of n-6 FA, & TFA. Limit consumption of processed, deep-fried foods, and alcohol. |
- |
Pregnant/breastfeeding or women with increased requirement for long chain n-3 fatty acids, should be advised to consume an algae-based supplement of known nutrient content. |
Pregnant/breastfeeding or women with increased requirement for long chain n-3 fatty acids, should be advised to consume an algae-based supplement of known nutrient content. |
- |
|
Combined-Males and Females |
- |
Improve n-3 nutritional status by regular consumption of good sources of alpha-linolenic acid. |
- |
- |
- |
|
|
Burdge et al., 2017 [37] |
Females |
- |
- |
- |
Need more cognitive studies for children of vegetarian females due to low DHA levels. |
- |
|
Pinto et al., 2017 [9] |
Combined |
- |
Further research whether populations with low n-3 status are more at risk of having a pro-inflammatory profile. |
- |
- |
- |
|
Melina et al., 2016 [5] |
Females |
- |
- |
- |
Low dose microalgae DHA supplements for pregnancy and lactation. |
- |
|
Combined |
High intakes of LA may suppress ALA conversion. |
May be prudent to ensure higher intakes of ALA. N-3 needs of healthy individuals can be met with ALA alone; endogenous synthesis of EPA and DHA from ALA is sufficient. |
Clinical relevance of reduced EPA and DHA status in VGNs are unknown. Low-dose micro-algae based DHA supplements are available for those with increased needs. |
Clinical relevance of reduced EPA and DHA status in VGNs are unknown. Low-dose micro-algae based DHA supplements are available for those with increased needs. |
A ratio of LA/ALA not exceeding 4:1 has been suggested for optimal conversion. |
|
|
Harris, 2014 [38] |
Combined |
Consume more ALA and less LA to increase the Omega-3 Index. |
Consume more ALA and less LA to increase the Omega-3 Index. |
- |
- |
Omega-3 Index >8%. |
|
Sarter et al., 2014 [16] |
Combined |
- |
- |
VGNs respond robustly to a relatively low dose of a vegetarian omega-3 supplement. |
VGNs respond robustly to a relatively low dose of a vegetarian omega-3 supplement. |
No direct evidence that omega-3 index confers additional health benefits over and above their already protective VGN diet. |
|
Saunders et al., 2013 [15] |
Combined |
- |
Double current AI of ALA if no direct sources of EPA and DHA are consumed. Increased needs or reduced conversion may benefit from DHA and EPA supplements derived from microalgae of 200–300 mg/day of DHA and EPA (pregnant and lactating women) and reduced conversion ability. |
- |
- |
- |
|
Craig, 2010; Review [39] |
Combined |
- |
Regular consumption of plant foods naturally rich in n-3 fatty acid ALA, such as ground flaxseed, walnuts, soy products, and hemp seed beverages. |
- |
Pregnant and lactating women may benefit from DHA-fortified foods and microalgae-derived DHA supplements. |
- |
|
Welch et al., 2010 [40] |
Combined |
- |
Further research for conversion of ALA to long chain n-3 PUFAs for maintenance of adequate status in non-fish and fish-oil consumers is required. |
- |
- |
- |
|
Simopoulos, 2008 [41] |
Combined |
- |
- |
- |
- |
Ratio of 4:1 is associated with 70% decrease in total mortality in secondary prevention of cardiovascular disease. |
|
Kornsteiner et al., 2008 [32] |
Combined |
- |
Erythrocyte phospholipid n-3 status of VGNs is critical. It is important to maintain n-3 fatty acid intake during adult life. |
- |
- |
Ensure physical, mental and neurological health, reduce n-6/n-3 ratio with an additional intake of direct sources of EPA and DHA, regardless of age and gender. |
|
Mann et al., 2006 and Li et al., 1999 (Mann used data collected from Li, 1999) [42] |
Combined |
- |
Advised to increase intake of n-3 fatty acids to increase platelet PL n-3 PUFA and reduce platelet aggregability. |
- |
- |
- |
|
Rosell et al., 2005 [10] |
Combined |
- |
Suggests (with caution) VGNs increase intake of ALA and limit intake of LA to optimize FA status. The importance of long chain fatty acids in the diet needs further investigation. |
- |
- |
- |
|
Davis et al., 2003 [34] |
Combined |
5.5–8% Kcals from n-6 FA. Reduce high n-6 oils in the diet, decrease processed foods. Primary fat should be MUFA. |
<1.5–2% of calories should be obtained from n-3. Decrease EtOH and trans fatty acids to increase conversion of ALA to EPA and DHA. Double intake of ALA; provide >1% of energy from n-3 or about 1.1 g/1000 calories. Increased needs require 2.2 g/1000 calories. Aim for 2–4 g of ALA/day. |
Not essential, but it is important to ensure sufficient levels by relying on conversion from parent fatty acids. Two important steps to improve EPA status: 1) Maximize conversion of ALA to EPA and DHA and 2) Provide a direct source of EPA and DHA. |
Not technically essential, but ensure sufficient levels by relying on conversion from parent fatty acids. Supplements of DHA should be 100–300 mg/day. DHA and ALA should also be adequate. |
To achieve the 4:1 ratio, <1.5–2% of calories should be obtained from n-3 and 5.5–8% of calories from n-6 FA. |
|
Simopoulos, 2000 [35] |
Combined |
- |
- |
- |
- |
Ratio was 1:1 to 2:1 in ancient diets. Present ratio is 10 to 1:20 to 25 to 1- indicates a deficiency in Western Diets of n-3. |
|
Fokkema et al., 2000 [33] |
Combined |
- |
- |
- |
- |
Found a 3:1 ratio increased EPA from 0.3 to about 1.0% in VGN plasma. A 3.7 g/day ALA diet for 4 weeks (ratio of 3.8:1) augmented ALA about 1% more. Total n-3 levels increased from 2.3 to 3.4% in plasma. DHA status was not increased. |
|
Agren et al., 1995 [43] |
Combined |
Depressed levels of n-3 FA are depressed due to very low diet levels and high LA and oleic acid intake. |
- |
- |
- |
- |
|
Sanders et al., 1978 [44] |
Females |
- |
More research is needed on low delta 4 desaturase in lactating women. |
- |
- |
- |
|
Combined |
- |
Further research needed to establish whether differences in the proportion of n-3 derivatives in tissues are of physiological importance. |
- |
- |
- |
References
- Spector, A.; Kim, H.Y. Discovery of Essential Fatty Acids. J. Lipid Res. 2014, 56, 11–21.
- The National Academies of Sciences Engineering Medicine. Dietary Reference Intakes tables and application. Available online: http://nationalacademies.org/hmd/Activities/Nutrition/SummaryDRIs/DRI-Tables.aspx (accessed on 18 August 2019).
- Institute of Medicine (IOM). Introduction to dietary reference intakes. In Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academics Press: Washington, DC, USA, 2005; pp. 29–37.
- Institute of Medicine (IOM). Dietary fats: Total fat and fatty acids. In Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academics Press: Washington, DC, USA, 2005; pp. 470–472, 030908525X (pbk.) 0309085373 (hardcover).
- Melina, V.; Craig, W.; Levin, S. Position of the Academy of Nutrition and Dietetics: Vegetarian Diets. J. Acad. Nutr. Diet. 2016, 116, 1970–1980.
- Alles, B.; Baudry, J.; Mejean, C.; Touvier, M.; Peneau, S.; Hercberg, S.; Kesse-Guyot, E. Comparison of Sociodemographic and Nutritional Characteristics between Self-Reported Vegetarians, Vegans, and Meat-Eaters from the NutriNet-Sante Study. Nutrients 2017, 9, 1023.
- Clarys, P.; Deliens, T.; Huybrechts, I.; Deriemaeker, P.; Vanaelst, B.; De Keyzer, W.; Hebbelinck, M.; Mullie, P. Comparison of nutritional quality of the vegan, vegetarian, semi-vegetarian, pesco-vegetarian and omnivorous diet. Nutrients 2014, 6, 1318–1332.
- Rizzo, N.S.; Jaceldo-Siegl, K.; Sabate, J.; Fraser, G.E. Nutrient profiles of vegetarian and nonvegetarian dietary patterns. J. Acad. Nutr. Diet. 2013, 113, 1610–1619.
- Pinto, A.M.; Sanders, T.A.; Kendall, A.C.; Nicolaou, A.; Gray, R.; Al-Khatib, H.; Hall, W.L. A comparison of heart rate variability, n-3 PUFA status and lipid mediator profile in age- and BMI-matched middle-aged vegans and omnivores. Br. J. Nutr. 2017, 117, 669–685.
- Rosell, M.S.; Lloyd-Wright, Z.; Appleby, P.N.; Sanders, T.A.; Allen, N.E.; Key, T.J. Long-chain n-3 polyunsaturated fatty acids in plasma in British meat-eating, vegetarian, and vegan men. Am. J. Clin. Nutr. 2005, 82, 327–334.
- Elorinne, A.L.; Alfthan, G.; Erlund, I.; Kivimaki, H.; Paju, A.; Salminen, I.; Turpeinen, U.; Voutilainen, S.; Laakso, J. Food and Nutrient Intake and Nutritional Status of Finnish Vegans and Non-Vegetarians. PLoS ONE 2016, 11, e0148235.
- Perrin, M.T.; Pawlak, R.; Dean, L.L.; Christis, A.; Friend, L. A cross-sectional study of fatty acids and brain-derived neurotropic factor (BDNF) in human milk from lactating women following vegan vegetarian and omnivore diets. Eur. J. Clin. Nutr. 2019, 58, 2401–2410.
- Conquer, J.A.; Holub, B.J. Supplementation with an algae source of docosahexaenoic acid increases (n-3) fatty acid status and alters selected risk factors for heart disease in vegetarian subjects. J. Nutr. 1996, 126, 3032–3039.
- Geppert, J.; Kraft, V.; Demmelmair, H.; Koletzko, B. Docosahexaenoic acid supplementation in vegetarians effectively increases omega-3 index: A randomized trial. Lipids 2005, 40, 807–814.
- Saunders, A.V.; Davis, B.C.; Garg, M.L. Omega-3 polyunsaturated fatty acids and vegetarian diets. Med. J. Aust. 2013, 199, S22–S26.
- Sarter, B.; Kelsey, K.S.; Schwartz, T.A.; Harris, W.S. Blood docosahexaenoic acid and eicosapentaenoic acid in vegans: Associations with age and gender and effects of an algal-derived omega-3 fatty acid supplement. Clin. Nutr. 2015, 34, 212–218.
- U.S. Department of Agriculture (USDA). Nutrient Database. Available online: https://ndb.nal.usda.gov/ndb/nutrients/index (accessed on 11 June 2019).
- Ovega-3 i-Health Inc. Available online: https://www.ovega.com/ovega-3 (accessed on 25 October 2017).
- Deva Nutrition LLC. Available online: http://www.devanutrition.com/vegan-omega-3-dha-delayed-release.html (accessed on 25 October 2017).
- Nordic Naturals Inc. Available online: https://www.nordicnaturals.com/en/Products/Product_Details/514/?ProdID=1654 (accessed on 25 October 2017).
- Sanders, T.A.; Roshanai, F. Platelet phospholipid fatty acid composition and function in vegans compared with age- and sex-matched omnivore controls. Eur. J. Clin. Nutr. 1992, 46, 823–831.
- Burdge, G.C.; Wootton, S.A. Conversion of alpha-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br. J. Nutr. 2002, 88, 411–420.
- Burdge, G.C.; Jones, A.E.; Wootton, S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men*. Br. J. Nutr. 2002, 88, 355–363.
- Brenna, J.T. Efficiency of conversion of alpha-linolenic acid to long chain n-3 fatty acids in man. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 127–132.
- Sauerwald, T.U.; Hachey, D.L.; Jensen, C.L.; Chen, H.; Anderson, R.E.; Heird, W.C. Effect of dietary alpha-linolenic acid intake on incorporation of docosahexaenoic and arachidonic acids into plasma phospholipids of term infants. Lipids 1996, 31, S131–S135.
- Salem, N., Jr.; Wegher, B.; Mena, P.; Uauy, R. Arachidonic and docosahexaenoic acids are biosynthesized from their 18-carbon precursors in human infants. Proc. Natl. Acad. Sci. USA 1996, 93, 49–54.
- Emken, E.A.; Adlof, R.O.; Gulley, R.M. Dietary linoleic acid influences desaturation and acylation of deuterium-labeled linoleic and linolenic acids in young adult males. Biochim. Biophys. Acta 1994, 1213, 277–288.
- Akerele, O.A.; Cheema, S.K. A balance of omega-3 and omega-6 polyunsaturated fatty acids is important in pregnancy. J. Nutr. Intermed. Metab. 2016, 5, 23–33.
- Domenichiello, A.; Kitson, A.; Bazinet, R. Is docosahexaenoic acid synthesis from α-linolenic acid sufficient to supply the adult brain? Prog. Lipid Res. 2015, 59, 54–66.
- Barceló-Coblijn, G.; Murphy, E. Alpha-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res. 2009, 48, 355–374.
- Valenzuela, R.; Videla, L. The importance of the long-chain polyunsaturated fatty acid n-6 / n-3 ratio in development of non-alcoholic fatty liver associated with obesity. Food Funct. 2011, 2, 644–648.
- Kornsteiner, M.; Singer, I.; Elmadfa, I. Very low n-3 long-chain polyunsaturated fatty acid status in Austrian vegetarians and vegans. Ann. Nutr. Metab. 2008, 52, 37–47.
- Fokkema, M.R.; Brouwer, D.A.; Hasperhoven, M.B.; Martini, I.A.; Muskiet, F.A. Short-term supplementation of low-dose gamma-linolenic acid (GLA), alpha-linolenic acid (ALA), or GLA plus ALA does not augment LCP omega 3 status of Dutch vegans to an appreciable extent. Prostaglandins Leukot. Essent. Fat. Acids 2000, 63, 287–292.
- Davis, B.C.; Kris-Etherton, P.M. Achieving optimal essential fatty acid status in vegetarians: Current knowledge and practical implications. Am. J. Clin. Nutr. 2003, 78, 640S–646S.
- Simopoulos, A.P. Human requirement for N-3 polyunsaturated fatty acids. Poult. Sci. 2000, 79, 961–970.
- Agnoli, C.; Baroni, L.; Bertini, I.; Ciappellano, S.; Fabbri, A.; Papa, M.; Pellegrini, N.; Sbarbati, R.; Scarino, M.L.; Siani, V.; et al. Position paper on vegetarian diets from the working group of the Italian Society of Human Nutrition. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 1037–1052.
- Burdge, G.C.; Tan, S.Y.; Henry, C.J. Long-chain n-3 PUFA in vegetarian women: A metabolic perspective. J. Nutr. Sci. 2017, 6, e58.
- Harris, W.S. Achieving optimal n-3 fatty acid status: The vegetarian’s challenge... or not. Am. J. Clin. Nutr. 2014, 100, 449S–452S.
- Craig, W.J. Nutrition concerns and health effects of vegetarian diets. Nutr. Clin. Pract. 2010, 25, 613–620.
- Welch, A.A.; Shakya-Shrestha, S.; Lentjes, M.A.; Wareham, N.J.; Khaw, K.T. Dietary intake and status of n-3 polyunsaturated fatty acids in a population of fish-eating and non-fish-eating meat-eaters, vegetarians, and vegans and the product-precursor ratio [corrected] of alpha-linolenic acid to long-chain n-3 polyunsaturated fatty acids: Results from the EPIC-Norfolk cohort. Am. J. Clin. Nutr. 2010, 92, 1040–1051.
- Simopoulos, A. The importance of the omega6/omega 3 fatty acid ratio in cardiovascular disease and other diseases. Exp. Biol. Med. (Maywood) 2008, 223, 674–688.
- Mann, N.; Pirotta, Y.; O’Connell, S.; Li, D.; Kelly, F.; Sinclair, A. Fatty acid composition of habitual omnivore and vegetarian diets. Lipids 2006, 41, 637–646.
- Agren, J.J.; Tormala, M.L.; Nenonen, M.T.; Hanninen, O.O. Fatty acid composition of erythrocyte, platelet, and serum lipids in strict vegans. Lipids 1995, 30, 365–369.
- Sanders, T.A.; Ellis, F.R.; Dickerson, J.W. Studies of vegans: The fatty acid composition of plasma choline phosphoglycerides, erythrocytes, adipose tissue, and breast milk, and some indicators of susceptibility to ischemic heart disease in vegans and omnivore controls. Am. J. Clin. Nutr. 1978, 31, 805–813.
- Gerster, H. Can adults adequately convert alpha-linolenic acid (18:3n-3) to eicosapentaenoic acid (20:5n-3) and docosahexaenoic acid (22:6n-3)? Int. J. Vitam. Nutr. Res. 1998, 68, 159–173.
- Emken, E.A.; Adlof, R.O.; Duval, S.M.; Nelson, G.J. Effect of dietary arachidonic acid on metabolism of deuterated linoleic acid by adult male subjects. Lipids 1998, 33, 471–480.
- Mantzioris, E.; James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary substitution with an alpha-linolenic acid-rich vegetable oil increases eicosapentaenoic acid concentrations in tissues. Am. J. Clin. Nutr. 1994, 59, 1304–1309.
- Indu, M. n-3 fatty acids in Indian diets—Comparison of the effects of precursor (alpha-linolenic acid) Vs product (long chain n-3 poly unsaturated fatty acids). Nutr. Res. 1992, 12, 569–582.
- Mest, H.J.; Beitz, J.; Heinroth, I.; Block, H.U.; Forster, W. The influence of linseed oil diet on fatty acid pattern in phospholipids and thromboxane formation in platelets in man. Klin. Wochenschr. 1983, 61, 187–191.
- Li, D.; Sinclair, A.; Wilson, A.; Nakkote, S.; Kelly, F.; Abedin, L.; Mann, N.; Turner, A. Effect of dietary alpha-linolenic acid on thrombotic risk factors in vegetarian men. Am. J. Clin. Nutr. 1999, 69, 872–882.
- Sanders, T.A.; Younger, K.M. The effect of dietary supplements of omega 3 polyunsaturated fatty acids on the fatty acid composition of platelets and plasma choline phosphoglycerides. Br. J. Nutr. 1981, 45, 613–616.
- Bjerve, K.S.; Mostad, I.L.; Thoresen, L. Alpha-linolenic acid deficiency in patients on long-term gastric-tube feeding: Estimation of linolenic acid and long-chain unsaturated n-3 fatty acid requirement in man. Am. J. Clin. Nutr. 1987, 45, 66–77.


