Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Daria Shunkina (Skuratovskaia) | + 699 word(s) | 699 | 2021-07-15 05:58:16 | | | |
| 2 | Nora Tang | + 227 word(s) | 926 | 2021-08-20 02:49:38 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Shunkina (Skuratovskaia), D. Macrophages and Endothelial Dysfunction. Encyclopedia. Available online: https://encyclopedia.pub/entry/13357 (accessed on 07 February 2026).
Shunkina (Skuratovskaia) D. Macrophages and Endothelial Dysfunction. Encyclopedia. Available at: https://encyclopedia.pub/entry/13357. Accessed February 07, 2026.
Shunkina (Skuratovskaia), Daria. "Macrophages and Endothelial Dysfunction" Encyclopedia, https://encyclopedia.pub/entry/13357 (accessed February 07, 2026).
Shunkina (Skuratovskaia), D. (2021, August 19). Macrophages and Endothelial Dysfunction. In Encyclopedia. https://encyclopedia.pub/entry/13357
Shunkina (Skuratovskaia), Daria. "Macrophages and Endothelial Dysfunction." Encyclopedia. Web. 19 August, 2021.
Copy Citation
In clinical epidemiological studies, chronic stress was reported to be an independent risk factor for the development of atherosclerosis and the function of the intima. One of the hypotheses is based on the effect of chronic stress on the accumulation of macrophages in the intima and the acceleration of damage to the mucous membrane of vascular endothelial cells. Neutrophils, endothelial cells, platelets and fibroblasts are involved in the formation of a chronic inflammatory focus, accompanied by microcirculation disorders and angiogenic disorders. However, the main regulators of all cellular processes are immunocompetent cells and, predominantly, activated macrophages.
obesity
inflammation
macrophage
1. Macrophage Subpopulation M(Hb)
In addition to the M1 and M2 populations, a subpopulation of M (Hb) macrophages has been identified in areas of plaque hemorrhage in humans. The transition to the M (Hb) phenotype is stimulated by hemoglobin (Figure 1). M (Hb) macrophages are characterized by high levels of the mannose receptor (MR) (CD206) and CD163, a scavenger receptor for the hemoglobin/haptoglobin (Hb/Hp) complex (Figure 1). The Hb/Hp complex is involved in the clearance of hemoglobin from plaques after hemorrhage [1] and induces the secretion of anti-inflammatory cytokines, such as IL-10 via CD163/phosphoinositide 3-kinase and phosphatidylinositol 3-kinase (PI3K)/phospho-AKT pathways, in human macrophages in vitro and in tissue macrophages ex vivo [2][3]. M (Hb) macrophages are able to eliminate cholesterol by increasing the activity of the nuclear receptor X of the liver receptor (LXR) α (NR1H3), thereby preventing the formation of foam cells [4]. The increased LXRα activity also induces the expression of ferroportin, an iron exporter, thereby decreasing cellular iron concentrations and reducing ROS production by M (Hb) macrophages [4]. This decrease in ROS production in M (Hb) macrophages has been confirmed in atherosclerotic plaques in vivo [4][5][6].
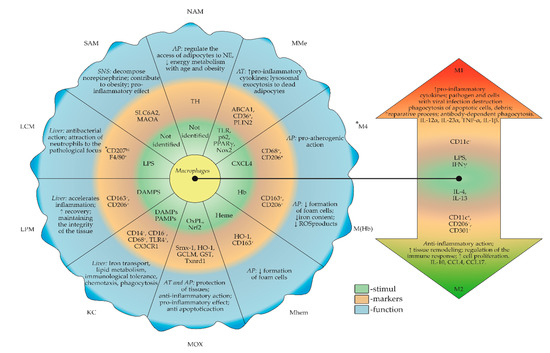
Figure 1. Inductors, markers and functions of macrophages subtypes in tissues. All macrophage subtypes may take a pro- or anti-inflammatory phenotype in obesity. MoMFs are a common precursor for subtypes of migrating macrophages in tissue. When damaged, MoMFs can assume a tissue-specific phenotype. Determining the exact origin of macrophages (resident or migratory) is a task in modern science. The plasticity of macrophages between M1 and M2 is also present in the resident forms and is shown by an arrow. The arrow indicates the stimuli, markers and functions that characterize both types of polarization. ABCA1—ATP binding cassette subfamily a member 1; AP—atherosclerosis plaque; AT—adipose tissue; DAMPS—damage-associated molecular patterns; GCLM—glutamate–cysteine ligase modifier subunit; GST—glutathione S-transferase; Hb/Hp—hemoglobin/haptoglobin; HO-1—heme oxygenase 1; IL—Interleukin; KC—Kupffer cells; LCM—liver capsular macrophages; LPM— large peritoneal macrophages; LPS—lipopolysaccharide; MAOA—monoamine oxidase A; MoMF—monocyte-derived macrophage; Mox—oxidized macrophages; MR—mannose receptor; NAM—neuro-associated macrophages; NE—norepinephrine; Nox2—NADPH oxidase-2; OxPL—oxidized phospholipids; PLIN2—Perilipin 2; PPARγ—peroxisome proliferator-activated receptor gamma; ROS—reactive oxygen species; SAM—sympathetic neuron-associated macrophages; SLC6A2—carrier family 6 member 2; SNS—sympathetic nervous system; Srnx-1—sulfiredoxin-1; TH—tyrosine hydroxylase; TLR—Toll-like receptor; Txnrd1—thioredoxin reductase 1; MMe—metabolically activated macrophages; *—mouse-specific markers; #—cannot be M1 and M2. This figure has been created by modifying the templates from Servier Medical Art (https://smart.servier.com).
2. Macrophage Subpopulation Mhem
During the endocytosis of the Hb/Hp complex, heme is released from erythrocytes, which stimulates the transition of macrophages to the Mhem phenotype (Figure 1). The main markers for Mhem macrophages are heme oxygenase 1 (HMOX1) and CD163 [5][6]. Mhem macrophages stimulate activating transcription factor (ATF)-1 in various ways, promoting LXRβ (NR1H2) and HMOX1 expression in humans. This process increases the expression of LXRα and ABCA1, which subsequently increase cholesterol efflux, associated with increased production of IL-10 and apolipoprotein (Apo) E [5][6][7]. Moreover, Mhem have an increased adaptation to plaque hemorrhage. Thus, the M (Hb) and Mhem phenotypes prevent foam cell formation and oxidative stress.
3. Macrophage Subpopulation M4
Another type of macrophage, iron-loaded M4 macrophages, is predominantly detected in areas of neovascularization in atherosclerotic plaques (Figure 1). After migration through the endothelium, monocytes under the influence of CXCL4 differentiate into M4. The main markers of M4 macrophages are CD68+MR+ [5][6][8] (Figure 1).
M4 macrophages are called CXCL4-differentiated macrophages that express the phenotypic markers metalloproteinase 7 (MMP7) and calcium-binding protein S100A8 [9]. At the same time, M4 macrophages do not express CD163 and exhibit low expression of the scavenger receptors CD36 or SR-1, which leads to a failure to induce the expression of the atheroprotective protein HMOX1 when cells are exposed to the Hb/Hp complex [5][6][10]. Thus, M4 macrophages have a pro-atherogenic profile and can be involved in complications of late atherosclerosis, such as acute coronary syndrome and arterial thrombosis. They produce the enzyme MMP12, which can be involved in the degradation of the fibrous coating of the plaque and the plaque destabilization. Furthermore, M4 macrophages express IL-6 and TNF-α, which increase inflammation. However, the fundamental role of M4 cells in atherogenesis is unknown and requires research [5][6].
Using scRNA-seq technology, a new type of macrophage, Trem2hi, has been identified, which is characterized by high expression of Trem2 (triggering receptors expressed on myeloid cells 2), Spp1 (secreted phosphoprotein1), Ctsl (cathepsin L) and CD9. The number of Trem2hi macrophages in plaques decreases with a high-fat diet [11]. Trem2 controls the expression of genes associated with energy metabolism and lipid catabolism [12].
Thus, in individuals with obesity, the structure and function of the cardiovascular system adapt to excess body weight. Metabolic disorders, such as obesity, are accompanied by endothelial cell dysfunction and decreased vascular density [13]. The modern paradigm argues that metabolic changes are associated with obesity secondary to endothelial dysfunction. The hypothesis regarding the ability of the endothelium to cause metabolic dysregulation itself must be revised and supplemented.
References
- Nielsen, M.J.; Møller, H.J.; Moestrup, S.K. Hemoglobin and heme scavenger receptors. Antioxid. Redox Signal. 2010, 12, 261–273.
- Philippidis, P.; Mason, J.C.; Evans, B.J.; Nadra, I.; Taylor, K.M.; Haskard, D.O.; Landis, R.C. Hemoglobin scavenger receptor CD163 mediates interleukin-10 release and heme oxygenase-1 synthesis: Antiinflammatory monocyte-macrophage responses in vitro, in resolving skin blisters in vivo, and after cardiopulmonary bypass surgery. Circ. Res. 2004, 94, 119–126.
- Landis, R.C.; Philippidis, P.; Domin, J.; Boyle, J.J.; Haskard, D.O. Haptoglobin Genotype-Dependent Anti-Inflammatory Signaling in CD163(+) Macrophages. Int. J. Inflamm. 2013, 2013, 980327.
- Finn, A.V.; Nakano, M.; Polavarapu, R.; Karmali, V.; Saeed, O.; Zhao, X.; Yazdani, S.; Otsuka, F.; Davis, T.; Habib, A.; et al. Hemoglobin directs macrophage differentiation and prevents foam cell formation in human atherosclerotic plaques. J. Am. Coll. Cardiol. 2012, 59, 166–177.
- Colin, S.; Chinetti-Gbaguidi, G.; Staels, B. Macrophage phenotypes in atherosclerosis. Immunol Rev. 2014, 262, 153–166.
- Chistiakov, D.A.; Bobryshev, Y.V.; Nikiforov, N.G.; Elizova, N.V.; Sobenin, I.A.; Orekhov, A.N. Macrophage phenotypic plasticity in atherosclerosis: The associated features and the peculiarities of the expression of inflammatory genes. Int. J. Cardiol. 2015, 184, 436–445.
- Boyle, J.J.; Johns, M.; Kampfer, T.; Nguyen, A.T.; Game, L.; Schaer, D.J.; Mason, J.C.; Haskard, D.O. Activating transcription factor 1 directs Mhem atheroprotective macrophages through coordinated iron handling and foam cell protection. Circ. Res. 2012, 110, 20–33.
- Bories, G.; Colin, S.; Vanhoutte, J.; Derudas, B.; Copin, C.; Fanchon, M.; Daoudi, M.; Belloy, L.; Haulon, S.; Zawadzki, C.; et al. Liver X receptor activation stimulates iron export in human alternative macrophages. Circ. Res. 2013, 113, 1196–1205.
- Erbel, C.; Tyka, M.; Helmes, C.M.; Akhavanpoor, M.; Rupp, G.; Domschke, G.; Linden, F.; Wolf, A.; Doesch, A.; Lasitschka, F.; et al. CXCL4-induced plaque macrophages can be specifically identified by co-expression of MMP7+S100A8+ in vitro and in vivo. Innate Immun. 2015, 21, 255–265.
- Gleissner, C.A.; Shaked, I.; Erbel, C.; Böckler, D.; Katus, H.A.; Ley, K. CXCL4 downregulates the atheroprotective hemoglobin receptor CD163 in human macrophages. Circ. Res. 2010, 106, 203–211.
- Deng, H.; Sun, Y.; Zeng, W.; Li, H.; Guo, M.; Yang, L.; Lu, B.; Yu, B.; Fan, G.; Gao, Q.; et al. New Classification of Macrophages in Plaques: A Revolution. Curr Atheroscler Rep. 2020, 22, 31.
- Liu, C.; Li, P.; Li, H.; Wang, S.; Ding, L.; Wang, H.; Ye, H.; Jin, Y.; Hou, J.; Fang, X.; et al. TREM2 regulates obesity-induced insulin resistance via adipose tissue remodeling in mice of high-fat feeding. J. Transl. Med. 2019, 17, 300.
- Graupera, I.; Coll, M.; Pose, E.; Elia, C.; Piano, S.; Solà, E.; Blaya, D.; Huelin, P.; Solé, C.; Moreira, R.; et al. Adipocyte Fatty-Acid Binding Protein is Overexpressed in Cirrhosis and Correlates with Clinical Outcomes. Sci. Rep. 2017, 7.
More
Information
Subjects:
Others
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
988
Revisions:
2 times
(View History)
Update Date:
20 Aug 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No