 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | S. Rodriguez-Rojo | + 5773 word(s) | 5773 | 2021-08-11 05:52:18 | | | |
| 2 | S. Rodriguez-Rojo | Meta information modification | 5773 | 2021-08-14 19:53:00 | | | | |
| 3 | Vivi Li | Meta information modification | 5773 | 2021-08-16 03:11:25 | | | | |
| 4 | S. Rodriguez-Rojo | Meta information modification | 5773 | 2021-08-20 08:52:27 | | |
Video Upload Options
Oxidative- and inflammatory-related ocular surface diseases have high prevalence and are an emerging issue in ophthalmology. Olive pomace (OP) is the olive oil’s industry main by-product, and is potentially environmentally hazardous. Nevertheless, it contains phenolic compounds with important bioactivities, like oleuropein (OL) and hydroxytyrosol (HT). The antioxidant and anti-inflammatory effects of four OP extracts (CONV, OPT(1–3)), pure OL and HT, and mixtures thereof were screened on human corneal (HCE) and conjunctival epithelial (IM-ConjEpi) cells. CONV was conventionally extracted, while OPT(1–3) were produced by pressurized liquid extraction. Thanks to their improved activity, CONV and OPT3 (HT-enriched) were selected for dose-dependent studies. Cells were stimulated with tumor necrosis factor-α or ultraviolet-B radiation, measuring interleukin (IL)-1β, IL-6, IL-8, and IL-17A as well as interferon γ-induced protein [IP]-10 secretion or intracellular ROS production, respectively. On HCE, both extracts and HT inhibited the secretion of most measured ILs, demonstrating a strong anti-inflammatory effect; while in IM-ConjEpi, all samples decreased IP-10 secretion. Moreover, HT, OL, and both extracts showed strong dose-dependent antioxidant activity in both cell lines. Compared with CONV, OPT3 was active at lower concentrations, demonstrating that intensified extraction techniques are selective towards targeted biomarkers. Hence, a high-value application as potential ocular surface therapy was proposed for the OP valorization.
1. Introduction
2. Preparation of Phenolic Solutions
HT and OL were dissolved in culture medium (DMEM/F-12 + GlutaMax, without any supplement), and freeze dried OP extracts CONV, OPT1, OPT2, and OPT3 were dissolved in 0.4% EtOH in culture medium, on the same day of the experiment.
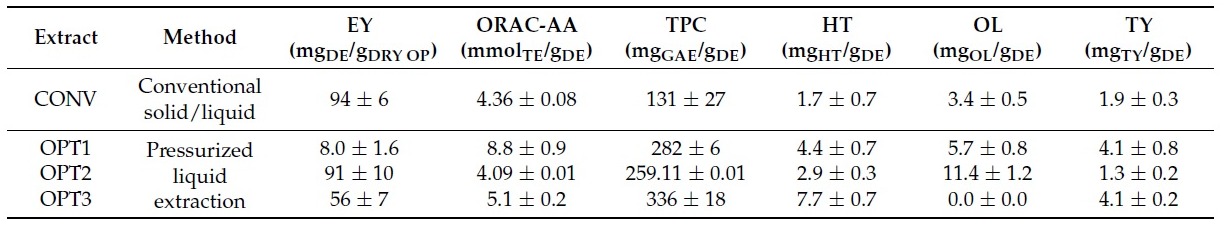
The OP extracts were prepared as previously described [38]. The CONV extract was prepared by conventional solid-liquid extraction from freeze dried OP while OPT1, OPT2 and OPT 3 were produced by pressurized liquid extraction from freeze-dried OP previously deffated by supercritical carbon dioxide extraction. Their composition is described in Table 1. The extraction conditions for OPT1, OPT2 and OPT3 were optimized by a desing of experiments [38] so that they correspond to the extract with the highest chemical antioxidant activity by ORAC assay (Oxigen Radical Absorbance Capacity assay), the highest concentration of OL, and the highest concentration of HT, TY and TPC (total phenolic concentration), respectively.
Table 1. Characterization of the conventional (CONV) and the three optimized (OPT1, OPT2, and OPT3) olive pomace (OP) extracts in terms of extraction yield (EY) (as mg of dry extract (DE)/g of DRY OP), oxygen radical absorbance capacity antioxidant activity (ORAC-AA) (as mmol of Trolox equivalents (TE)/g of DE), and total phenolic content (TPC) (as mg of gallic acid equivalents (GAE)/g of DE), as well as extract richness in Hydroxytyrosol (HT), Oleuropein (OL) and Tyrosol(TY) (as mg of compound/g of DE). The results are presented as average ± standard deviation.
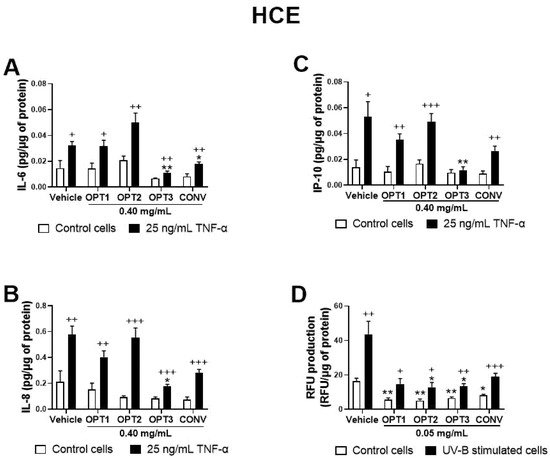
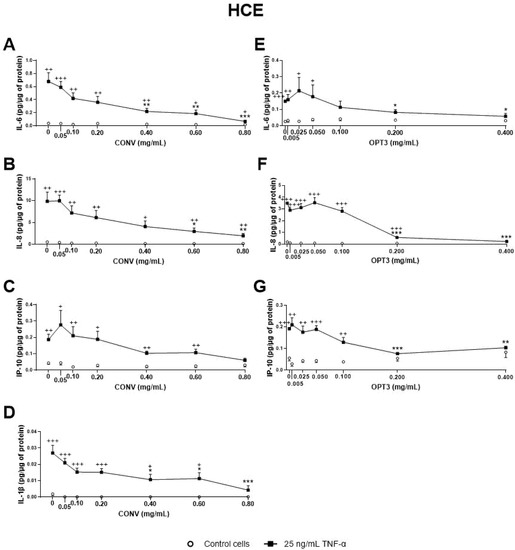
3. Anti-Inflammatory Activity of Phenolic Solutions
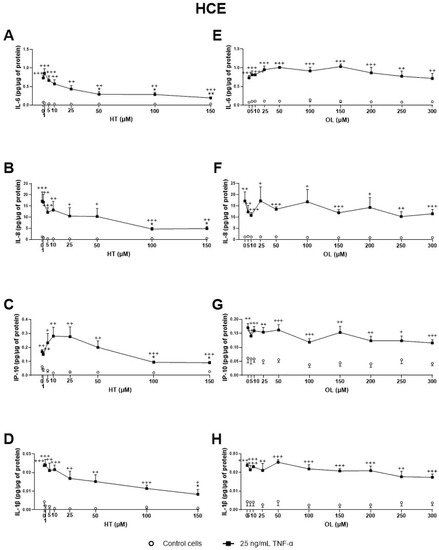
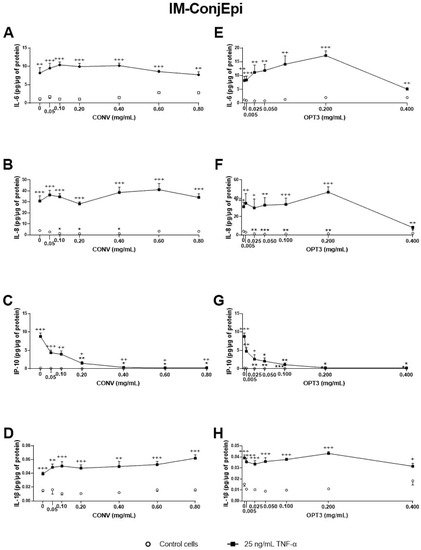
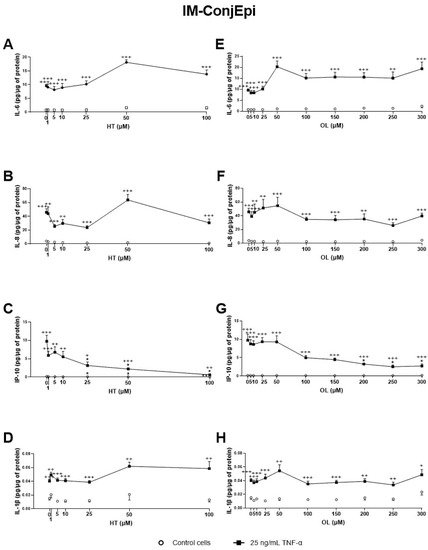
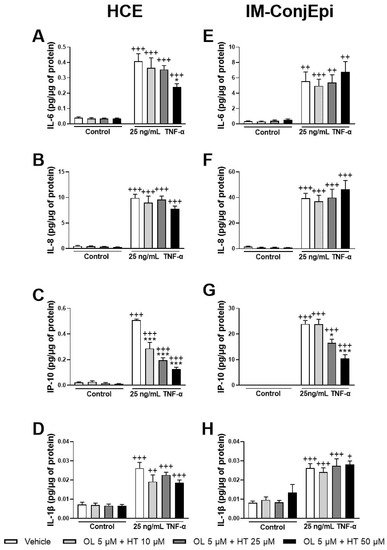
4. Antioxidant Activity of Phenolic Solutions
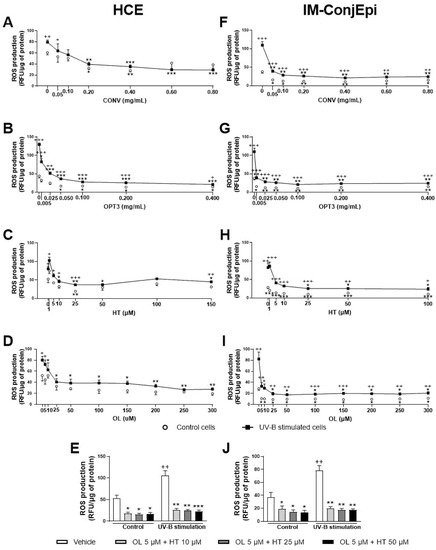
5. Discussion
References
- Huerva, V.; Ascaso, F.J.; Grzybowski, A. Ocular inflammation. Mediat. Inflamm. 2015, 2015, 1–2.
- Giasson, B.I.; Duda, J.E.; Murray, I.V.J.; Chen, Q.; Souza, J.M.; Hurtig, H.I.; Ischiropoulos, H.; Trojanowski, J.Q.; Lee, V.M.Y. Oxidative damage linked to neurodegeneration by selective α-synuclein nitration in synucleinopathy lesions. Science 2000, 290, 985–989.
- Winkler, B.S.; Boulton, M.E.; Gottsch, J.D.; Sternberg, P. Oxidative damage and age-related macular degeneration. Mol. Vis. 1999, 5, 32.
- Imamura, Y.; Noda, S.; Hashizume, K.; Shinoda, K.; Yamaguchi, M.; Uchiyama, S.; Shimizu, T.; Mizushima, Y.; Shirasawa, T.; Tsubota, K. Drusen, choroidal neovascularization, and retinal pigment epithelium dysfunction in SOD1-deficient mice: A model of age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 11282–11287.
- Bhuyan, K.C.; Bhuyan, D.K.; Podos, S.M. Lipid peroxidation in cataract of the human. Life Sci. 1986, 38, 1463–1471.
- Ward, S.K.; Wakamatsu, T.H.; Dogru, M.; Ibrahim, O.M.A.; Kaido, M.; Ogawa, Y.; Matsumoto, Y.; Igarashi, A.; Ishida, R.; Shimazaki, J.; et al. The role of oxidative stress and inflammation in conjunctivochalasis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1994–2002.
- Wakamatsu, T.H.; Dogru, M.; Ayako, I.; Takano, Y.; Matsumoto, Y.; Ibrahim, O.M.A.; Okada, N.; Satake, Y.; Fukugawa, K.; Shimazaki, J.; et al. Evaluation of lipid oxidative stress status and inflammation in atopic ocular surface disease. Mol. Vis. 2010, 16, 2465–2475.
- Craig, J.P.; Nelson, J.D.; Azar, D.T.; Belmonte, C.; Bron, A.J.; Chauhan, S.K.; de Paiva, C.S.; Gomes, J.A.P.; Hammitt, K.M.; Jones, L.; et al. TFOS DEWS II Report Executive Summary. Ocul. Surf. 2017, 15, 802–812.
- Nakamura, S.; Shibuya, M.; Nakashima, H.; Hisamura, R.; Masuda, N.; Imagawa, T.; Uehara, M.; Tsubota, K. Involvement of oxidative stress on corneal epithelial alterations in a blink-suppressed dry eye. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1552–1558.
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.-S.; Schaumberg, D.; Od, S.D.; Uchino, M.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365.
- McDonald, M.; Patel, D.A.; Keith, M.S.; Snedecor, S.J. Economic and Humanistic Burden of Dry Eye Disease in Europe, North America, and Asia: A Systematic Literature Review. Ocul. Surf. 2016, 14, 144–167.
- Kojima, T.; Dogru, M.; Kawashima, M.; Nakamura, S.; Tsubota, K. Advances in the diagnosis and treatment of dry eye. Prog. Retin. Eye Res. 2020, 78, 100842.
- Abengózar-Vela, A.; Calonge, M.; Stern, M.E.; González-García, M.J.; Enríquez-De-Salamanca, A. Quercetin and resveratrol decrease the inflammatory and oxidative responses in human ocular surface epithelial cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2709–2719.
- Abengózar-Vela, A.; Schaumburg, C.S.; Stern, M.E.; Calonge, M.; Enríquez-de-Salamanca, A.; González-García, M.J. Topical Quercetin and Resveratrol Protect the Ocular Surface in Experimental Dry Eye Disease. Ocul. Immunol. Inflamm. 2019, 27, 1023–1032.
- Abdallah, M.; Marzocco, S.; Adesso, S.; Zarrouk, M.; Guerfel, M. Olive oil polyphenols extracts inhibit inflammatory markers in J774A.1 murine macrophages and scavenge free radicals. Acta Histochem. 2018, 120, 1–10.
- Romero-García, J.M.; Niño, L.; Martínez-Patiño, C.; Álvarez, C.; Castro, E.; Negro, M.J. Biorefinery based on olive biomass. State of the art and future trends. Bioresour. Technol. 2014, 159, 421–432.
- Gómez-Muñoz, B.; Hatch, D.J.; Bol, R.; Garca-Ruiz, R. The Compost of Olive Mill Pomace: From a Waste to a Resource-Environmental Benefits of Its Application in Olive Oil Groves. In Sustainable Development-Authoritative and Leading Edge Content for Environmental Management; InTechphen Ltd.: Rijeka, Croatia, 2012.
- Rodrigues, F.; Pimentel, F.B.; Oliveira, M.B.P.P. Olive by-products: Challenge application in cosmetic industry. Ind. Crop. Prod. 2015, 70, 116–124.
- Lozano-Sánchez, J.; Giambanelli, E.; Quirantes-Piné, R.; Cerretani, L.; Bendini, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Wastes generated during the storage of extra virgin olive oil as a natural source of phenolic compounds. J. Agric. Food Chem. 2011, 59, 11491–11500.
- Obied, H.K.; Bedgood, D.R.; Prenzler, P.D.; Robards, K. Chemical screening of olive biophenol extracts by hyphenated liquid chromatography. Anal. Chim. Acta 2007, 603, 176–189.
- Vazquez-Roncero, A.; Maestro-Duran, R.; Graciani-Constante, E. Phenolic compounds in olive fruits. II: Polyphenols in vegetation water. Grasas Aceites 1974, 25, 341–345.
- Servili, M.; Baldioli, M.; Selvaggini, R.; Miniati, E.; Macchioni, A.; Montedoro, G. High-performance liquid chromatography evaluation of phenols in olive fruit, virgin olive oil, vegetation waters, and pomace and 1D- and 2D-nuclear magnetic resonance characterization. J. Am. Oil Chem. Soc. 1999, 76, 873–882.
- Mulinacci, N.; Romani, A.; Galardi, C.; Pinelli, P.; Giaccherini, C.; Vincieri, F.F. Polyphenolic content in olive oil waste waters and related olive samples. J. Agric. Food Chem. 2001, 49, 3509–3514.
- Capasso, R.; Evidente, A.; Visca, C. Production of hydroxytyrosol from olive oil vegetation waters. Agrochimica 1994, 38, 165–172.
- Visioli, F.; Romani, A.; Mulinacci, N.; Zarini, S.; Conte, D.; Vincieri, F.F.; Galli, C. Antioxidant and other biological activities of olive mill waste waters. J. Agric. Food Chem. 1999, 47, 3397–3401.
- Ceccon, L.; Saccù, D.; Procida, G.; Cardinali, S. Liquid Chromatographic Determination of Simple Phenolic Compounds in Waste Waters from Olive Oil Production Plants. J. AOAC Int. 2001, 84, 1739–1744.
- Fernández-Bolaños, J.; Rodríguez, G.; Rodríguez, R.; Heredia, A.; Guillén, R.; Jimínez, A. Production in large quantities of highly purified hydroxytyrosol from liquid-solid waste of two-phase olive oil processing or “alperujo”. J. Agric. Food Chem. 2002, 50, 6804–6811.
- Plastina, P.; Benincasa, C.; Perri, E.; Fazio, A.; Augimeri, G.; Poland, M.; Witkamp, R.; Meijerink, J. Identification of hydroxytyrosyl oleate, a derivative of hydroxytyrosol with anti-inflammatory properties, in olive oil by-products. Food Chem. 2019, 279, 105–113.
- Benincasa, C.; La Torre, C.; Plastina, P.; Fazio, A.; Perri, E.; Caroleo, M.C.; Gallelli, L.; Cannataro, R.; Cione, E. Hydroxytyrosyl Oleate: Improved Extraction Procedure from Olive Oil and By-Products, and In Vitro Antioxidant and Skin Regenerative Properties. Antioxidants 2019, 8, 233.
- Granner, T.; Maloney, S.; Antecka, E.; Correa, J.A.; Burnier, M.N. 3,4 Dihydroxyphenyl Ethanol Reduces Secretion of Angiogenin in Human Retinal Pigment Epithelial Cells. Br. J. Ophthalmol. 2013, 97, 371–374.
- Zou, X.; Feng, Z.; Li, Y.; Wang, Y.; Wertz, K.; Weber, P.; Fu, Y.; Liu, J. Stimulation of GSH synthesis to prevent oxidative stress-induced apoptosis by hydroxytyrosol in human retinal pigment epithelial cells: Activation of Nrf2 and JNK-p62/SQSTM1 pathways. J. Nutr. Biochem. 2012, 23, 994–1006.
- Zhu, L.; Liu, Z.; Feng, Z.; Hao, J.; Shen, W.; Li, X.; Sun, L.; Sharman, E.; Wang, Y.; Wertz, K.; et al. Hydroxytyrosol protects against oxidative damage by simultaneous activation of mitochondrial biogenesis and phase II detoxifying enzyme systems in retinal pigment epithelial cells. J. Nutr. Biochem. 2010, 21, 1089–1098.
- Liu, Z.; Sun, L.; Zhu, L.; Jia, X.; Li, X.; Jia, H.; Wang, Y.; Weber, P.; Long, J.; Liu, J. Hydroxytyrosol protects retinal pigment epithelial cells from acrolein-induced oxidative stress and mitochondrial dysfunction. J. Neurochem. 2007, 103, 2690–2700.
- Deiana, M.; Aruoma, O.I.; Bianchi, M.D.L.P.; Spencer, J.P.E.; Kaur, H.; Halliwell, B.; Aeschbach, R.; Banni, S.; Dessi, M.A.; Corongiu, F.P. Inhibition of peroxynitrite dependent DNA base modification and tyrosine nitration by the extra virgin olive oil-derived antioxidant hydroxytyrosol. Free Radic. Biol. Med. 1999, 26, 762–769.
- González-Correa, J.A.; Rodríguez-Pérez, M.D.; Márquez-Estrada, L.; López-Villodres, J.A.; Reyes, J.J.; Rodriguez-Gutierrez, G.; Fernández-Bolaños, J.; De La Cruz, J.P. Neuroprotective Effect of Hydroxytyrosol in Experimental Diabetic Retinopathy: Relationship with Cardiovascular Biomarkers. J. Agric. Food Chem. 2018, 66, 637–644.
- Şimşek, T.; Altınışık, U.; Erşan, İ.; Şahin, H.; Altınışık, B.; Erbaş, M.; Pala, Ç. Prevention of intraocular pressure elevation with oleuropein rich diet in rabbits, during the general anaesthesia. Springerplus 2016, 5.
- Di Mauro, M.D.; Fava, G.; Spampinato, M.; Aleo, D.; Melilli, B.; Saita, M.G.; Centonze, G.; Maggiore, R.; D’Antona, N. Polyphenolic Fraction from Olive Mill Wastewater: Scale-Up and in vitro Studies for Ophthalmic Nutraceutical Applications. Antioxidants 2019, 8, 462.
- Katsinas, N.; da Silva, A.B.; Enríquez-de-Salamanca, A.; Fernández, N.; Bronze, M.R.; Rodríguez-Rojo, S. Pressurized Liquid Extraction Optimization from Supercritical Defatted Olive Pomace: A Green and Selective Phenolic Extraction Process. ACS Sustain. Chem. Eng. 2021, 9, 5590–5602.
- Paladino, R.A.; Miller, S.N.; Kleiber, K.F.; Byers, D.M. Resveratrol reverses the effect of TNF-α on inflammatory markers in a model of autoimmune uveitis. Eur. J. Integr. Med. 2020, 36, 101137.
- Stoddard, A.R.; Koetje, L.R.; Mitchell, A.K.; Schotanus, M.P.; Ubels, J.L. Bioavailability of antioxidants applied to stratified human corneal epithelial cells. J. Ocul. Pharmacol. Ther. 2013, 29, 681–687.
- Woźniak, A.; Paduch, R. Aloe vera extract activity on human corneal cells. Pharm. Biol. 2012, 50, 147–154.
- Kim, Y.-H.; Oh, T.; Park, E.; Yim, N.-H.; Park, K.; Cho, W.; Ma, J. Anti-Inflammatory and Anti-Apoptotic Effects of Acer Palmatum Thumb. Extract, KIOM-2015EW, in a Hyperosmolar-Stress-Induced In vitro Dry Eye Model. Nutrients 2018, 10, 282.
- Park, B.; Lee, I.S.; Hyun, S.W.; Jo, K.; Lee, T.G.; Kim, J.S.; Kim, C.S. The protective effect of Polygonum cuspidatum (PCE) aqueous extract in a dry eye model. Nutrients 2018, 10, 1550.
- Tishler, M.; Yaron, I.; Geyer, O.; Shirazi, I.; Naftaliev, E.; Yaron, M. Elevated tear interleukin-6 levels in patients with Sjogren syndrome. Ophthalmology 1998, 105, 2327–2329.
- Pflugfelder, S.C.; Jones, D.; Ji, Z.; Afonso, A.; Monroy, D. Altered cytokine balance in the tear fluid and conjunctiva of patients with Sjogren’s syndrome keratoconjunctivitis sicca. Curr. Eye Res. 1999, 19, 201–211.
- Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Zheng, X.; Butler, M.; Farley, W.; Stern, M.; Li, D.; Pflugfelder, S. Desiccating Stress Promotes Th-17 Pattern Response in the Conjunctiva. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5420.
- Yoon, K.C.; Jeong, I.Y.; Park, Y.G.; Yang, S.Y. Interleukin-6 and tumor necrosis factor-α levels in tears of patients with dry eye syndrome. Cornea 2007, 26, 431–437.
- Lam, H.; Bleiden, L.; de Paiva, C.S.; Farley, W.; Stern, M.E.; Pflugfelder, S.C. Tear Cytokine Profiles in Dysfunctional Tear Syndrome. Am. J. Ophthalmol. 2009, 147.
- Jones, D.; Monroy, D.; Ji, Z.; Atherton, S.; Pflugfelder, S. Sjögren’s syndrome: Cytokine and Epstein-Barr viral gene expression within the conjunctival epithelium. Invest. Ophthalmol. Vis. Sci. 1994, 35, 3493–3504.
- Yoon, K.C.; Park, C.S.; You, I.C.; Choi, H.J.; Lee, K.H.; Im, S.K.; Park, H.Y.; Pflugfelder, S.C. Expression of CXCL9, -10, -11, and CXCR3 in the tear film and ocular surface of patients with dry eye syndrome. Investig. Ophthalmol. Vis. Sci. 2010, 51, 643–650.
- Enríquez-de-Salamanca, A.; Castellanos, E.; Stern, M.E.; Fernández, I.; Carreño, E.; García-Vázquez, C.; Herreras, J.M.; Calonge, M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. Mol. Vis. 2010, 16, 862–873.
- Corrales, R.M.; Villarreal, A.; Farley, W.; Stern, M.E.; Li, D.Q.; Pflugfelder, S.C. Strain-related cytokine profiles on the murine ocular surface in response to desiccating stress. Cornea 2007, 26, 579–584.
- Yoon, K.C.; De Paiva, C.S.; Qi, H.; Chen, Z.; Farley, W.J.; Li, D.Q.; Pflugfelder, S.C. Expression of Th-1 chemokines and chemokine receptors on the ocular surface of C57BL/6 mice: Effects of desiccating stress. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2561–2569.
- Solomon, A.; Dursun, D.; Liu, Z.; Xie, Y.; Macri, A.; Pflugfelder, S.C. Pro- and anti-inflammatory forms of interleukin-1 in the tear fluid and conjunctiva of patients with dry-eye disease. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2283–2292.
- Enríquez-de-Salamanca, A.; Calonge, M. Cytokines and chemokines in immune-based ocular surface inflammation. Expert Rev. Clin. Immunol. 2008, 4, 457–467.
- Calonge, M. Classification of ocular atopic/allergic disorders and conditions: An unsolved problem. Acta Ophthalmol. Scand. Suppl. 1999, 77, 10–13.
- Leonardi, A.; Curnow, S.J.; Zhan, H.; Calder, V.L. Multiple cytokines in human tear specimens in seasonal and chronic allergic eye disease and in conjunctival fibroblast cultures. Clin. Exp. Allergy 2006, 36, 777–784.
- Hingorani, M.; Calder, V.L.; Buckley, R.J.; Lightman, S.L. The role of conjunctival epithelial cells in chronic ocular allergic disease. Exp. Eye Res. 1998, 67, 491–500.
- De Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D.; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.F.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253.
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990.
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504–511.
- Wakamatsu, T.H.; Dogru, M.; Tsubota, K. Tearful relations: Oxidative stress, inflammation and eye diseases. Arq. Bras. Oftalmol. 2009, 71, 72–79.
- Wenk, J.; Brenneisen, P.; Meewes, C.; Wlaschek, M.; Peters, T.; Blaudschun, R.; Ma, W.; Kuhr, L.; Schneider, L.; Scharffetter-Kochanek, K. UV-induced oxidative stress and photoaging. Curr. Probl. Dermatol. 2001, 29, 83–94.
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubotav, K. Potential role of oxidative stress in ocular surface inflammation and dry eye disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168.
- Čejkova, J.; Ardan, T.; Jirsová, K.; Jechová, G.; Malec, J.; Šimonová, Z.; Čejka, Č.; Filipec, M.; Dotřelová, D.; Brunová, B. The role of conjunctival epithelial cell xanthine oxidoreductase/xanthine oxidase in oxidative reactions on the ocular surface of dry eye patients with Sjögren’s syndrome. Histol. Histopathol. 2007, 22, 997–1003.
- Čejková, J.; Ardan, T.; Šimonová, Z.; Čejka, Č.; Malec, J.; Dotřelova, D.; Brůnová, B. Decreased expression of antioxidant enzymes in the conjunctival epithelium of dry eye (Sjögren’s syndrome) and its possible contribution to the development of ocular surface oxidative injuries. Histol. Histopathol. 2008, 23, 1477–1483.
- Birkedal-Hansen, H.; Moore, W.G.I.; Bodden, M.K.; Windsor, L.J.; Birkedal-Hansen, B.; DeCarlo, A.; Engler, J.A. Matrix metalloproteinases: A review. Crit. Rev. Oral Biol. Med. 1993, 4, 197–250.
- Chen, M.; Hu, D.N.; Pan, Z.; Lu, C.W.; Xue, C.Y.; Aass, I. Curcumin protects against hyperosmoticity-induced IL-1β elevation in human corneal epithelial cell via MAPK pathways. Exp. Eye Res. 2010, 90, 437–443.
- Seo, M.J.; Kim, J.M.; Lee, M.J.; Sohn, Y.S.; Kang, K.K.; Yoo, M. The therapeutic effect of DA-6034 on ocular inflammation via suppression of MMP-9 and inflammatory cytokines and activation of the MAPK signaling pathway in an experimental dry eye model. Curr. Eye Res. 2010, 35, 165–175.
- Xiao, F.; Cui, H.; Zhong, X. Beneficial effect of daidzin in dry eye rat model through the suppression of inflammation and oxidative stress in the cornea. Saudi J. Biol. Sci. 2018, 25, 832–837.
- Cavet, M.E.; Harrington, K.L.; Vollmer, T.R.; Ward, K.W.; Zhang, J.Z. Anti-inflammatory and anti-oxidative effects of the green tea polyphenol epigallocatechin gallate in human corneal epithelial cells. Mol. Vis. 2011, 17, 533–542.
- Luo, L.J.; Lai, J.Y. Epigallocatechin Gallate-Loaded Gelatin-g-Poly(N-Isopropylacrylamide) as a New Ophthalmic Pharmaceutical Formulation for Topical Use in the Treatment of Dry Eye Syndrome. Sci. Rep. 2017, 7, 1–14.
- Tseng, C.-L.; Hung, Y.-J.; Chen, Z.-Y.; Fang, H.-W.; Chen, K.-H. Synergistic Effect of Artificial Tears Containing Epigallocatechin Gallate and Hyaluronic Acid for the Treatment of Rabbits with Dry Eye Syndrome. PLoS ONE 2016, 11, e0157982.
- Huang, H.Y.; Wang, M.C.; Chen, Z.Y.; Chiu, W.Y.; Chen, K.H.; Lin, I.C.; Yang, W.C.V.; Wu, C.C.; Tseng, C.L. Gelatin–epigallocatechin gallate nanoparticles with hyaluronic acid decoration as eye drops can treat rabbit dry-eye syndrome effectively via inflammatory relief. Int. J. Nanomed. 2018, 13, 7251–7273.
- Li, J.; Deng, R.; Hua, X.; Zhang, L.; Lu, F.; Coursey, T.G.; Pflugfelder, S.C.; Li, D.Q. Blueberry Component Pterostilbene Protects Corneal Epithelial Cells from Inflammation via Anti-oxidative Pathway. Sci. Rep. 2016, 6, 1–10.
- Chen, H.-C.; Chen, Z.-Y.; Wang, T.-J.; Drew, V.; Tseng, C.-L.; Fang, H.-W.; Lin, F.-H. Herbal Supplement in a Buffer for Dry Eye Syndrome Treatment. Int. J. Mol. Sci. 2017, 18, 1697.
- Li, Y.J.; Luo, L.J.; Harroun, S.G.; Wei, S.C.; Unnikrishnan, B.; Chang, H.T.; Huang, Y.F.; Lai, J.Y.; Huang, C.C. Synergistically dual-functional nano eye-drops for simultaneous anti-inflammatory and anti-oxidative treatment of dry eye disease. Nanoscale 2019, 11, 5580–5594.
- Foti, M.C. Antioxidant properties of phenols. J. Pharm. Pharmacol. 2007, 59, 1673–1685.
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849.
- Songa, Y.; Buettner, G.R. Thermodynamic and kinetic considerations for the reaction of semiquinone radicals to form superoxide and hydrogen peroxide. Free Radic. Biol. Med. 2010, 49, 919–962.
- Rodríguez-Bonilla, P.; Gandía-Herrero, F.; Matencio, A.; García-Carmona, F.; López-Nicolás, J.M. Comparative Study of the Antioxidant Capacity of Four Stilbenes Using ORAC, ABTS+, and FRAP Techniques. Food Anal. Methods 2017, 10, 2994–3000.
- Wang, W.; Jing, T.; Yang, X.; He, Y.; Wang, B.; Xiao, Y.; Shang, C.; Zhang, J.; Lin, R. Hydroxytyrosol regulates the autophagy of vascular adventitial fibroblasts through the SIRT1-mediated signaling pathway. Can. J. Physiol. Pharmacol. 2018, 96, 88–96.
- Kumar, N.; Gorai, B.; Gupta, S.; Shiva; Goel, N. Extrapolation of hydroxytyrosol and its analogues as potential anti-inflammatory agents. J. Biomol. Struct. Dyn. 2020, 1–12.
- Lee, H.S.; Choi, J.H.; Cui, L.; Li, Y.; Yang, J.M.; Yun, J.J.; Jung, J.E.; Choi, W.; Yoon, K.C. Anti-inflammatory and antioxidative effects of Camellia japonica on human corneal epithelial cells and experimental dry eye: In vivo and in vitro study. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1196–1207.
- Cui, L.; Lee, H.S.; Li, Y.; Choi, J.H.; Yun, J.J.; Jung, J.E.; Choi, W.; Yoon, K.C. Experimental and clinical applications of chamaecyparis obtusa extracts in dry eye disease. Oxid. Med. Cell. Longev. 2017, 1–12.
- Paduch, R.; Woźniak, A.; Niedziela, P.; Rejdak, R. Assessment of eyebright (Euphrasia officinalis L.) extract activity in relation to human corneal cells using in vitro tests. Balk. Med. J. 2014, 31, 29–36.
- Paduch, R.; Woźniak, A. The effect of Lamium album extract on cultivated human corneal epithelial cells (10.014 PRSV-T). J. Ophthalmic Vis. Res. 2015, 10, 229–237.
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621.
- Lee, H.; Shim, W.; Kim, C.E.; Choi, S.Y.; Lee, H.; Yang, J. Therapeutic efficacy of nanocomplex of poly(ethylene glycol) and catechin for dry eye disease in a mouse model. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1682–1691.
- Shim, W.; Kim, C.E.; Lee, M.; Lee, S.H.; Park, J.; Do, M.; Yang, J.; Lee, H. Catechin solubilization by spontaneous hydrogen bonding with poly(ethylene glycol) for dry eye therapeutics. J. Control. Release 2019, 307, 413–422.
- Chung, S.; Choi, S.H.; Choi, J.A.; Chuck, R.; Joo, C. Curcumin suppresses ovalbumin-induced allergic conjunctivitis. Mol. Vis. 2012, 18, 1966–1972.
- Choi, W.; Lee, J.B.; Cui, L.; Li, Y.; Li, Z.; Choi, J.S.; Lee, H.S.; Yoon, K.C. Therapeutic Efficacy of Topically Applied Antioxidant Medicinal Plant Extracts in a Mouse Model of Experimental Dry Eye. Oxid. Med. Cell. Longev. 2016, 1–10.
- Kim, K.A.; Hyun, L.C.; Jung, S.H.; Yang, S.J. The leaves of Diospyros kaki exert beneficial effects on a benzalkonium chloride–induced murine dry eye model. Mol. Vis. 2016, 22, 284–293.
- Zhang, C.; Li, K.; Yang, Z.; Wang, Y.; Si, H. The Effect of the Aqueous Extract of Bidens Pilosa L. on Androgen Deficiency Dry Eye in Rats. Cell. Physiol. Biochem. 2016, 39, 266–277.
- Peng, Q.-H.; Yao, X.-L.; Wu, Q.-L.; Tan, H.-Y.; Zhang, J.-R. Effects of extract of Buddleja officinalis eye drops on androgen receptors of lacrimal gland cells of castrated rats with dry eye. Int. J. Ophthalmol. 2010, 3, 43–48.
- Chen, M.-H.; Tsai, C.-F.; Hsu, Y.-W.; Lu, F. Epigallocatechin gallate eye drops protect against ultraviolet B–induced corneal oxidative damage in mice. Mol. Vis. 2014, 20, 153–162.


